











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Los sistemas de producción bovina de doble propósito representan alrededor del 70% de ranchos y ganaderos en los trópicos y subtrópicos (Rangel et al., 2020), debido a su flexibilidad, así como una mejor inversión y necesidad de soporte técnico en comparación a sistemas más intensivos (Magaña- Monforte et al., 2006; Rojo-Rubio et al., 2009; Rangel et al., 2020). Estos sistemas se caracterizan por el uso de animales cruzados de razas lecheras Bos taurus x Bos indicus, donde las vacas son ordeñadas con el becerro al pie para estimular la bajada de la leche. La base de su alimentación es el pastoreo directo de gramíneas nativas o introducidas de bajo valor nutritivo y disponibilidad, sobre todo en la época de menor precipitación pluvial (Magaña-Monforte et al., 2006; Aguilar-Pérez et al., 2009; Peniche-González et al., 2014). Esta situación no contribuye a satisfacer los requerimientos nutricionales de las vacas para la producción de leche durante el posparto temprano, provocando que las vacas pierdan peso y condición corporal para subsanar la demanda nutricional, en especial energía. Las vacas bajo esas condiciones experimentan un balance energético negativo, muchas veces severo, que va en detrimento del reinicio de la actividad posparto temprano (Montiel y Ahuja, 2005; Aguilar- Pérez et al., 2009; Rojo-Rubio et al., 2009; González-Padilla, 2018).
En vacas productoras de leche el grado del balance energético negativo durante el posparto temprano se correlaciona con los días a la primera ovulación (Roche et al., 2000; Tanaka et al., 2008), asimismo, la reducción del consumo de alimento modifica los patrones de desarrollo folicular en la vaca y contribuye a la variación en la respuesta ovulatoria (Hess et al., 2005; Crowe, 2008; Aguilar-Pérez et al., 2009; Tinoco-Magaña et al., 2012).
Uno de los factores que condicionan el intervalo a la primera ovulación posparto es el desarrollo folicular que ocurre luego del parto, el cual está muy relacionado con el consumo de alimento (Kawashima et al., 2012). A pesar de la importancia de estos eventos reproductivos, son escasos los estudios que relacionan el efecto de la nutrición sobre el desarrollo de la actividad ovárica en el posparto temprano en vacas de los trópicos. El objetivo del trabajo fue caracterizar la actividad ovárica y el comportamiento reproductivo durante 98 días posparto de vacas multíparas de doble propósito en pastoreo con y sin suplementación en el trópico subhúmedo.
MATERIALES Y MÉTODOS
Localización del Estudio y Animales
El trabajo se realizó en la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Autónoma de Yucatán. El clima de la región es cálido subhúmedo (AWO), con lluvias en verano, temperatura anual entre 25- 28 °C, precipitación pluvial media de 984 mm y humedad relativa entre 75-80% (INEGI, 2004).
Se utilizó un experimento completamente al azar con 30 vacas multíparas de más de dos partos con diferente grado de encaste de Bos taurus x Bos indicus, con un peso pro- medio de 474.9 ± 58.3 kg y una condición corporal (CC) de 4.7 ± 0.8 en la escala del 1 al 9, donde 1= emaciadas y 9= obesas (Ayala et al., 1995). Las vacas se ordeñaban con un equipo mecánico tipo Tandem de cuatro plazas y se realizaba dos veces al día (05:00 y 15:00 h) con el becerro al pie para estimular la bajada de la leche. El becerro se criaba bajo un esquema de amamantamiento restringido por 30 minutos después de cada ordeña y se le dejaba uno de los cuartos de la ubre durante los primeros 45 días de vida. Posteriormente, se ordeñaba la ubre completa y el becerro solo mamaba leche residual.
La obtención de datos de cada vaca se realizó desde el día del parto hasta 98 días posparto. Las vacas fueron introducidas al estudio conforme fueron pariendo y se formaron dos grupos al azar de 15 animales por grupo. Al grupo 1 (grupo con suplemento), además del pastoreo se le suministró un su- plemento comercial y el grupo 2 (grupo sin suplemento) solo se mantuvo en pastoreo sin suplementación. Los animales de ambos grupos se mantuvieron en un solo lote en pasto Estrella de África (Cynodon nlemfuensis).
Pastoreo y Suplementación
Para el pastoreo, se utilizaron 14 ha de pasto Estrella de África bajo riego sin fertilizar. El pastoreo fue rotacional y el horario de pastoreo fue entre 08:00-14:00 y 17:00-05:00 h (posterior a cada ordeño). Todas las vacas tuvieron libre acceso a sales minerales y agua fresca durante su permanencia en el corral (antes y después de cada ordeño).
Las vacas del grupo 1 fueron suplementadas durante la ordeña, con un alimento comercial a base de cereales; sorgo, soya y salvado, con 16.3% de proteína cruda y 2.6 Mcal/KgMs de energía y fueron adaptadas al suplemento 15 días antes de la fecha estimada de parto. Para la asignación del suplemento se consideró un consumo total de materia seca (MS) del 3% del peso vivo al parto (Kearl, 1982). El concentrado fue asignado de manera que representara un 30% del con- sumo total estimado de MS (Combellas, 1998), manteniéndose fijo durante toda la prueba, y se dividió entre las dos ordeñas. En general se les proporcionaba un promedio de 4.3±0.5 kg de concentrado comercial en base fresca por día.
Las vacas del grupo control solamente recibieron una cantidad mínima de salvado de trigo con el fin de mantenerlas quietas durante el ordeño. El salvado fue asignado a un 5% del consumo total estimado de MS (en promedio 300 g/vaca/ordeña).
Manejo Reproductivo, Peso y Condición Corporal
Las vacas estuvieron expuestas durante todo el estudio con dos toros fértiles Holstein x Cebú bajo un esquema de monta libre. Las vacas fueron observadas dos ve- ces al día en periodos de media hora cada vez (07:00 y 17:00 h) para la detección del estro (Richards et al., 1989). El criterio de inicio del estro fue la aceptación de la monta homosexual cuando menos en dos ocasiones o la monta directa del toro. La preñez se diagnosticó por ultrasonido a los 30, 60 y 90 días posparto, para calcular la tasa de preñez a los 90 días posparto.
El peso vivo se registró en una báscula ganadera con capacidad para 1500 kg y una precisión de 5 kg y la CC se evaluó visualmente. El peso vivo y la CC se registró por una misma persona cada siete días desde el parto hasta los 98 días posparto.
Ovarios y Folículos
Los ovarios fueron observados mediante un ecógrafo Pie Medical (Falco Vet-100, Holanda) equipado con un transductor lineal de 6-8 Mhz. Las estructuras observadas se registraron individualmente tomando en cuenta el número y el diámetro de los folículos; así como su ubicación en los ovarios para el seguimiento de su desarrollo. También se registró la presencia o ausencia del cuerpo lúteo. Las mediciones de las estructuras ováricas se realizaron cada segundo día a partir del día 7 posparto y finalizaron cuando se completaron los 98 días posparto o cuando se diagnosticó preñez positiva.
Solamente se consideró la presencia de folículos grandes (folículos dominantes >10 mm; Perry et al., 1991). Se determinó como actividad ovárica el intervalo parto-primer folículo dominante (IPPF), el tiempo de permanencia del primer folículo dominante (TPPF) (intervalo entre el día de su primera detección [>10 mm] y el momento en que disminuyó por debajo de 10 mm), el número de folículos dominantes antes del primer estro (NFDAE) y del primer cuerpo lúteo (NFDACL). Se consideró como comportamiento reproductivo el intervalo parto-primer estro (IPPE), el intervalo parto-primer cuerpo lúteo (IPPCL) y la tasa de preñez (TP).
Análisis Estadístico
Las variables de peso vivo, CC, IPPF, TPPF, NFDAE, NFDACL, IPPE e intervalo IPPCL fueron analizados mediante análisis de varianza considerando como unidades experimentales a los 15 animales por tratamiento. Para el análisis de las variables número de vacas que presentaron estro, número de vacas que presentaron primer CL posparto y tasa de gestación se utilizó la prueba exacta de Fisher. Se utilizó el programa estadístico Statgraphics Plus 1999.
RESULTADOS
Peso Vivo y Condición Corporal
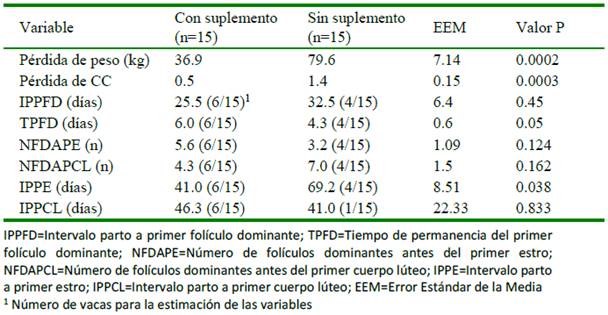
El peso promedio al parto de las vacas de los grupos con y sin suplemento fue de 481.0 y 467.0 kg, respectivamente, y la condición corporal al parto (CCP) fue de 4.7 y 4.6, respectivamente, sin diferencia significativa entre grupos. En ambos grupos de vacas se registró pérdida de peso y CC durante los 98 días posparto, siendo mayor en las vacas sin suplemento (-79 vs. -37 kg de peso vivo y -1.4 vs -0.5 de condición corporal) (p<0.05; Cuadro 1). Es importante resaltar que la pérdida de peso vivo y de condición corporal del grupo suplementado fue hasta los 42 días posparto, mientras que en las va- cas sin suplementación fue hasta el final del estudio (figuras 1 y 2).
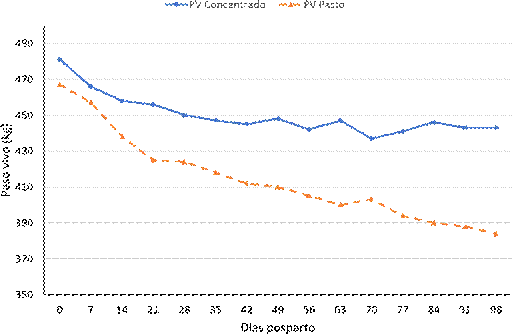
Figura 1 Cambios de peso vivo de vacas posparto Bos taurus x Bos indicus bajo dos esquemas de alimentación (solo pastoreo y pastoreo con suplementación)
Actividad Ovárica y Comportamiento Reproductivo
Se observó una pobre actividad ovárica posparto en ambos grupos de vacas (Cuadro 1). Los días de permanencia del folículo dominante fue mayor y el intervalo parto primer estro menor en las vacas con suplementación posparto (p<0.05; Cuadro 1).
Las vacas suplementadas presentaron un intervalo parto primer folículo dominante menor de siete días en comparación con las vacas no suplementadas, pero dicha diferencia no fue estadísticamente significativa (p=0.45). Tampoco resultaron significativas las diferencias para el número de folículos grandes (dominantes) antes del primer estro y antes del primer cuerpo lúteo posparto (Cuadro 1).
El intervalo parto primer cuerpo lúteo fue similar en las vacas de los dos tratamientos (p> 0.833); sin embargo, seis de 15 vacas en el grupo suplementado presentaron CL mientras que solamente una de las 15 vacas del grupo sin suplemento presento CL durante la prueba. Por otra parte, la tasa de gestación bue baja, siendo de 27% en el grupo suplementado y de 7% en el grupo sin suplementación (p>0.05).
DISCUSIÓN
Ambos grupos de vacas (sin y con suplemento) perdieron peso y condición corporal, aunque las pérdidas fueron mayores para las del grupo sin suplemento (17 vs 7.6% de peso y 1.4 vs 0.5 de CC), ya que los animales requieren usar las reservas corporales para proporcionar la energía necesaria para la producción de leche, dado que la capacidad de consumo se encuentra disminuida (Butler, 2003; Freetly et al., 2006; Wathes et al., 2007; Aguilar-Pérez et al., 2009; Kawashima et al., 2012).
Las vacas de doble propósito manejadas bajo pastoreo de gramíneas y leguminosas y con acceso de suplementos de cereales tienden a mantener tanto el peso vivo como la condición corporal, mientras que las que no reciben el suplemento experimentan mayor balance energético negativo (Laflamme y Connor 1992; Pérez-Aguilar et al., 2009; Tinoco-Magaña et al., 2012; Peniche- González et al., 2014), observándose la misma tendencia en el presente estudio.
Los cambios de peso vivo, CC y el balance energético negativo durante los primeros 20 días posparto afectan grandemente las funciones reproductivas de las vacas lecheras (Butler, 2003; Crowe, 2008; Tanaka et al., 2008;). No obstante, las vacas de doble propósito con buena CC durante el posparto temprano presentaron un mayor número de folículos grandes (>10 mm) que las vacas con pobre CC, mientras que la frecuencia de cuerpos lúteos no fue diferente por efecto de la CC (Domínguez et al., 2007). A este respecto, Diskin et al. (2003) señalaron que en el posparto temprano de vacas de carne y de leche no suplementadas no afecta notoriamente el crecimiento de folículos pequeños y medianos a diferencia del crecimiento de folículos de mayor tamaño, impidiendo una posible ovulación, como se observó en el presente estudio (Cuadro 1).
El intervalo de la aparición del primer folículo dominante después del parto es variable. En vacas de carne con cría al pie se ha observado folículos >8 mm de diámetro desde el día 7 y aumentando hasta el día 42 (Spicer et al., 1986) y la presencia de folículos dominantes el día 11 posparto, con un rango entre 5 y 39 días (Roche y Boland, 1991). En vacas lecheras, Kawashima et al. (2012) señalaron que la primera onda folicular ocurre alrededor del día 5 posparto y el primer folículo dominante el día 10, aunque solamente cerca del 50% llega a ovular. En el presente estudio, el intervalo de la aparición del primer folículo dominante fue de 25 y 32 días en los animales con y sin suplemento, respectivamente. Ambos valores son mayores al promedio de días de aparición del folículo dominante mencionado por los otros autores, aunque dentro de los rangos citados (Cuadro 1).
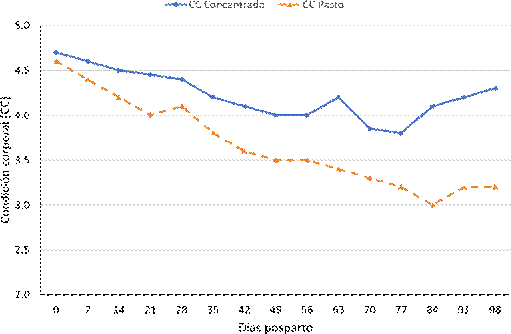
Figura 2 Cambios de condición corporal de vacas posparto Bos taurus x Bos indicus bajo dos esquemas de alimentación (solo pastoreo y pastoreo con suplementación)
La concentración de FSH circulante se incrementan en la primera semana posparto en vacas lecheras, independientemente de la dieta suministrada, iniciando el desarrollo de la primera onda folicular (Gong et al., 2002), lo cual puede haber ocurrido en el presente estudio ya que no se observó diferencias en el intervalo al primer folículo entre grupos. Por otra parte, se ha observado que el balance energético negativo es generalmente mayor durante el periodo posparto temprano, aunque aparentemente no afecta la población de folículos de 3-5 o 6-9 mm presentes en los ovarios entre el día 8 y 25 posparto (Beam y Butler, 1997; Lucy et al., 1999). Los resultados del presente estudio parecen estar de acuerdo con lo mencionado, ya que no se observó diferencias significativas entre los grupos en cuanto al intervalo parto primer folículo dominante, pero con solamente el 40 y 26% de las vacas suplementadas y no suplementadas, respectivamente.
El tiempo de permanencia del primer folículo dominante fue de 6 días en promedio en el grupo suplementado y de 4 días en el grupo con solo pastoreo (p<0.05). La restricción de la dieta provoca decremento gradual de la tasa de crecimiento, del diámetro máximo y la permanencia del folículo dominante (Murphy et al., 1991; Rhodes et al., 1996).
Stephen et al. (1997) con vacas Holstein alimentadas con dietas con baja, media y sin adición de grasa reportaron que la permanencia del folículo de la primera onda que ovuló tuvo un rango de 3 a 19 días, pero fue más larga en promedio que el folículo de la primera onda que no ovuló (10.8 ± 1.1. vs. ± 1.1 días). Los resultados del presente trabajo coinciden con los resultados de Stephen et al. (1997), en particular con los del folículo de la primera onda que no ovuló.
Se puede especular que la permanencia corta del folículo de la primera onda que no ovula se debe en parte a su falta de capacidad esteroidogénica, ya que se ha observado diferencias en la producción de estradiol entre los folículos que ovulan y los que no ovulan (Stephen et al., 1997), o bien a la insulina, ya que se conoce su efecto sobre la función de las células foliculares de varias especies, incluyendo el bovino o a los factores de crecimiento parecidos a la insulina IGF- 1 (Spicer y Stewart, 1996). La permanencia de un folículo dominante representa la fase durante la cual dicho folículo aparece y se mantiene funcionando o deja de crecer, en espera de la señal que determinará su destino final: la maduración seguida de ovulación, o bien la atresia y su eventual desaparición.
No se observaron diferencias significativas entre grupos (p> 0.05) con respecto al número de folículos grandes antes del primer estro, situación similar al reporte de Domínguez et al. (2007) con vacas suple- mentadas antes y después del parto. Es bien conocido el hecho de que las vacas de carne que amamantan tienen un intervalo más largo del parto a la primera ovulación (Roche y Boland, 1991; Wettemann et al., 2003), situación que concuerda con los resultados del presente estudio, ya que las vacas de ambos grupos presentaron varios folículos dominantes antes del primer estro y la aparición del primer CL posparto.
Se ha señalado que los cambios de peso vivo y de CC de vacas de carne durante el periodo posparto afectan tanto la duración del anestro como la tasa de gestación (Montiel y Ahuja, 2005; Crowe, 2008). En el presente estudio, las vacas suplementadas presentaron el intervalo parto primer estro en un menor tiempo (41 días posparto) en comparación con las vacas del grupo solo pastoreo (69 días posparto) (p<0.05). Estos intervalos, sin embargo, se encuentran dentro de los rangos establecidos en el ganado de carne (Mukasa-Mugerwa et al., 1991: Bolaños et al., 1996; Delgado et al., 2004). En vacas multíparas, la CC al parto se ha identificado como el principal factor que afecta el intervalo del primer estro posparto y la tasa de gestación (Selk et al., 1988). Short et al. (1990) argumentan que, si bien las pérdidas de CC después del parto prolongan la duración del anestro, sus efectos son amortigua- dos por una buena alimentación que permita llegar al parto con una CC adecuada. En el presente estudio fue evidente que la mayor pérdida de peso y CC en el grupo de solo pastoreo influyó marcadamente en el alargamiento del primer estro posparto y en el número de animales que presentaron estro antes de los 98 días posparto. Esto conlleva al intervalo de 550 días entre partos señalados en el trópico mexicano (Rojo-Rubio et al., 2009).
La proporción de animales suplementados que presentaron CL fue de 40% en comparación con el 6% del grupo de solo pastoreo, con un intervalo parto primer CL de 46 y 41 días respectivamente (p>0.05; Cuadro 1). En este sentido, Aguilar-Pérez et al. (2009) reportaron 58 y 30% de vacas con CL en los grupos suplementados y no suplementado, similar al estudio de Tinoco-Magaña et al. (2012) con 50% en vacas suplementadas y 33% en vacas no suplementadas. Sin embargo, Ruiz et al. (2008) y Domínguez et al. (2007) no encontraron este nivel de respuesta en vacas Holstein x Brahman o mestizas, respectivamente. En el presente caso, esta falta de significancia se debe tomar con reserva debido al pequeño número de animales que tuvieron CL. Asimismo, es probable que la pérdida de peso de las vacas de solo pastoreo (17%) no haya sido suficiente para provocar un retraso en la aparición del primer CL posparto. Diskin et al. (2003) señalan que los animales comienzan a presentar un esta- do de anestro cuando pierden un promedio de 22 a 24% de su peso inicial.
La tasa de gestación anual del ganado es uno de los indicadores más importantes para medir la eficiencia reproductiva en una explotación. Este indicador está condicionado por intervalo entre partos. En el trópico se reportan tasas de preñez de 37-40% a los 120 días posparto (Delgado et al., 2004; Aban et al., 2008), pudiendo mencionarse el estudio de Osorio-Arce y Segura-Correa (2002). En sistemas de producción de doble propósito reportaron a 120 días postparto evaluando más de 5000 vacas en diversas unidades de producción una tasa de gestación de 51.2% incluyendo debido principalmente a una alta incidencia de anestro posparto (Mukasa- Murgerwa et al., 1991; Bolaños et al., 1996). En un estudio en sistemas de producción de doble propósito en Yucatán, México con más de 5000 vacas, Osorio-Arce y Segura-Correa (2002) reportaron una tasa de gestación de 51.2% a los 120 días posparto. Asimismo, Aguilar-Pérez et al. (2009) Tinoco Magaña et al. (2012) encontraron grandes diferencias en tasas de gestación entre animales suplementados y no suplementados. En el presente trabajo, las vacas suplementadas presentaron 37% de gestación en comparación al 7/% en el grupo no suplementado, aunque sin diferencia significativa.
Vacas que pierden un punto más de CC durante la lactancia temprana tienen mayor riesgo de baja fertilidad, retraso en el primer estro y la primera ovulación (Montiel y Ahuja., 2005; Lents et al., 2008; Cavestany et al., 2009). En el presente estudio, las vacas suplementadas solamente perdieron en promedio 0.5 puntos de CC mientras que las no suplementadas perdieron 1.5 puntos, aunque esta diferencia no fue significativa sobre los intervalos del parto al primer cuerpo lúteo y tasa de concepción si sobre el intervalo parto a primer estro.
CONCLUSIONES
La suplementación ayudó a mitigar las pérdidas de peso vivo y condición corporal posparto de vacas Bos taurus x Bos indicus al pastoreo sin afectar significativamente la actividad ovárica en términos del intervalo parto primer folículo dominante, número de folículo dominantes antes del primer estro y número de folículos dominante antes del primer cuerpo lúteo, así como mejorando el comportamiento reproductivo en términos del tiempo de permanencia del primer folículo dominante, intervalo parto primer estro, pero no la tasa de preñez.

