











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
El cuy doméstico Cavia porcellus (Linnaeus 1738) es una especie considerada como una fuente principal de proteína en los Andes (Dillard et al., 1972; Pascual et al., 2017). Estos animales han sido criados tradicionalmente por gran parte de los pueblos andinos como fuente de alimentos e ingresos económicos por su alta fecundidad, tasas de crecimiento rápidas, requerimientos de mantenimiento simples y carne con alto contenido proteico (Atter, 1998).
Se han venido realizando investigaciones en esta especie en áreas como la nutrición, sanidad, manejo, genética y sistemas de producción entre otros (Portocarrero e Hidalgo, 2015; Casart y Falconí, 2016; Pascual et al., 2017; Cruz et al., 2019). A pesar de ello, existe una tendencia a usar individuos con bajo potencial genético como reproductores, lo cual ocasiona que se generen respuestas variables en los aspectos productivos y reproductivos. Es por ello necesario adecuar los programas de mejoramiento genético y el conocimiento en las estimaciones de los parámetros genéticos (Olesen et al., 2000).
Parámetros genéticos como la heredabilidad y correlaciones sirven para la implementación de planes de mejoramiento genético que buscan mantener las líneas y mejorarlas constantemente en caracteres definidos (Jiang et al., 1999). Las diversas metodologías para la estimación de la heredabilidad y correlaciones genéticas hacen uso de registros productivos y reproductivos de los animales. Así mismo, utilizan la información genealógica para establecer relaciones de parentesco entre animales. La heredabilidad, como parámetro genético, ayuda a explicar el grado en que los genes controlan la expresión del rasgo, siendo además utilizado para evaluaciones genéticas basadas en el uso de modelos mixtos y genómicos (Gianola, s,f.; Gutiérrez, 2010; Mrode, 2014). Las estimaciones de heredabilidad para un rasgo pueden diferir entre especies y razas o pueden cambiar lentamente con el tiempo (Cassell, 2009). Por otro lado, las correlaciones genéticas indican el grado de relación o asociación genética entre dos rasgos originado por fenómenos como la pleiotropía y desequilibrio de ligamiento (Roff y Roff, 1997; van Rheenen et al., 2019) y su importancia radica en que pueden mejorar o retardar la respuesta de la selección (Etterson, 2016).
En la Estación Experimental Agraria Santa Ana se viene realizando la selección de cuyes desde hace más de 20 años. Las líneas genéticas se originaron con colectas de animales de zonas locales, luego se realizaron apareamientos y selección por características productivas y morfológicas como las mencionadas por Rivas y Rico (2014). El propósito fue generar líneas prolíficas y precoces en condiciones ambientales de la sierra central del Perú. Desde entonces se ha logrado uniformizar diversos rasgos morfológicos y algunos rasgos productivos y reproductivos (Reyes, 2015; Navarro, 2016); sin embargo, se desconocen los parámetros genéticos. El presente trabajo tuvo como objetivo estimar la heredabilidad y correlaciones genéticas de los rasgos productivos de cuyes de las líneas Saños y Mantaro de la Estación Experimental Agraria Santa Ana.
MATERIALES Y MÉTODOS
Registros
La información recopilada provino de cuyes de la Estación Experimental Agraria Santa Ana del Instituto Nacional de Innovación Agraria (INIA), ubicada en el distrito de El Tambo, Junín (Perú), a una altitud de 3320 msnm.
La información constó de registros de peso al nacimiento (PN), peso al destete (15 días de vida; PD), peso a la cuarta semana (P4), peso a la octava semana (P8), peso a la décimo tercera semana (P13), sexo (S) y total de nacidos (TN) de 1622 y 1874 cuyes de las líneas Saños y Mantaro, respectivamente, desde abril de 2017 hasta marzo de 2020. Además, se incluyeron registros del número de parto hasta el quinto parto (NP), temporada de parto (doce trimestres) (Tp) de 343 y 357 madres Saños y Mantaro, respectivamente. El número de datos de rasgos productivos de las dos líneas, así como el número de camadas consideradas para la estima de la varianza del ambiente materno (c2) se presentan en el Cuadro 1.
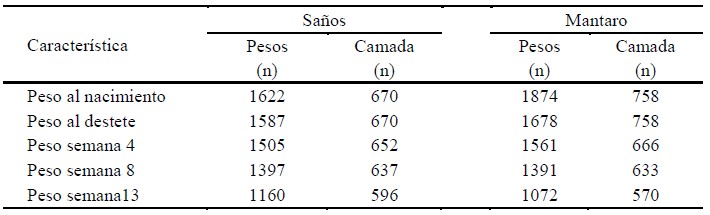
Sistema de Producción
La granja tiene áreas de cultivo para su propio abastecimiento de forraje. Los animales reciben una alimentación con base a forrajes y, en ocasiones, residuos de cosecha. El sistema de reproducción es el empadre continuo. Las instalaciones son pozas con divisiones de madera. Se manejan en promedio 440 animales de la línea Mantaro (154 madres y 33 reproductores) y 570 de la línea Saños (185 madres y 35 reproductores). En los registros diarios se anota información de características productivas y reproductivas de los animales, los cuales se encuentran identificados con aretes metálicos. Los datos para el presente trabajo corresponden a cuatro años e incluyen cinco y cuatro generaciones solapadas de las líneas Saños y Mantaro, respectivamente.
Análisis Estadístico
El cálculo del coeficiente de consanguinidad y la estimación de los componentes de varianza y las correlaciones fenotípicas y genéticas se realizaron por línea genética. El coeficiente de consanguinidad fue calculado con el uso del software VCE en las genera- ciones evaluadas de la línea Saños y Mantaro.
Los modelos ajustados para PN y PD (1) y para P4, P8 y P13 (2), basado en los modelos mixtos propuestos por Henderson (1984) y Mrode (2014), fueron y = xβ + Wc+ Zµ + Mm + e (1) y = xβ + Wc + Zµ + e (2), donde y es el vector de las observaciones de PN, PD, P4, P8 y P13; β es el vector de efectos fijos (sexo de la cría, número de parto, total de nacidos, temporada); µ es el vector de los efectos genéticos directos como aleatorio; c es el vector de efecto aleatorio de camada; m es el vector de efecto genético materno como aleatorio; e es el vector de los errores aleatorios asociados a cada observación, y X, Z, W y M son matrices de incidencia que relacionan registros con efectos ambientales fijos, animales y genéticos maternos.
Los componentes de varianza de todos los caracteres fueron estimados aplicando modelos univariados para cada carácter. Los efectos fijos que fueron incluidos en los modelos aplicados a cada parámetro fueron los que mostraron un efecto significativo. Los análisis de significancia de los efectos fijos se realizaron con la función nlme (Pinheiro et al., 2021) del software R v. 4.0.2 (R Core Team, 2020).
La heredabilidad fue calculada como la ratio de la varianza aditiva y la varianza fenotípica, obtenida como la suma de la varianza aditiva, la varianza debida a la camada y la varianza residual. Para la estimación de las correlaciones genéticas y fenotípicas se formularon modelos bivariados entre los rasgos productivos. Los componentes de varianzas y covarianzas y las correlaciones fenotípicas y genéticas se obtuvieron mediante el modelo animal del programa VCE6 v. 6.0.2 (Groeneveld, 2010), que estima componentes de varianzas por metodología REML con sus respectivos errores estándar.
RESULTADOS
La consanguinidad media poblacional fue 0.67 y 0.18% para las líneas Saños y Mantaro, respectivamente. La consanguinidad media en la línea Saños de cuyes consanguíneos fue 7.46% con un máximo de 12.50%, mientras que en la línea Mantaro la consanguinidad media de cuyes consanguíneos fue 8.72% con un valor máximo de 25%.
Los estadísticos descriptivos de las dos líneas de cuyes se presentan en el Cuadro 2. Se aprecia que los ejemplares de la línea Saños tienden a ser más pesados que los de la línea Mantaro, llegando a obtener un peso al beneficio o a las 13 semanas de 34.6 g más que en la línea Mantaro. Esto se evidencia igualmente en las distintas etapas de vida (PN, PD, P4, P8 y P13).
Cuadro 2 Valor promedio del rasgo (𝑥), desviación estándar (D.S), mínimo (Min.) y máximo (Max.) de las líneas de cuyes Saños y Mantaro (Junín, Perú)
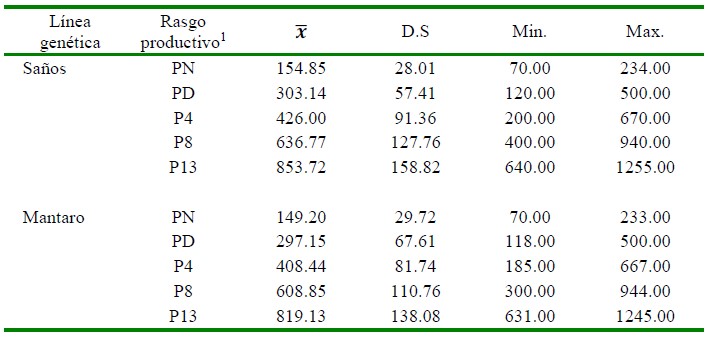
1 PN, PD, P4, P8, P13: pesos (g) al nacimiento, destete (15 d) y semanas 4, 8 y 13 de edad
Las heredabilidades directas de PN y PD y de los rasgos posdestete de los cuyes de la línea Saños se presentan en el Cuadro 3. La heredabilidad materna del peso al nacimiento y peso al destete (PNm y PDm) fue de 0.09 ± 0.02 y 0.05 ± 0.01, respectivamente. Las correlaciones genéticas fueron positivas y entre moderadas a altas, con excepción de la correlación entre PN y P13 de 0.28 ± 0.15. La correlación entre el valor genético directo de peso al nacimiento y el valor genético materno de peso al nacimiento fue negativa y alta (-0.77 ± 0.08). Del mismo modo, se encontró una correlación genética negativa (-0.22 ± 0.19) entre el valor genético de peso al destete directo y materno. Por otro lado, se observan correlaciones fenotípicas altas y positivas, siendo esta correlación mayor entre medidas próximas. La correlación entre PN y PD fue de 0.65, las correlaciones consecutivas entre PD, P4, P8 y P13 fueron 0.71,0.70 y 0.75, tal como se muestra en el Cuadro 3.
Cuadro 3 Heredabilidades en la diagonal y en negrita, correlaciones genéticas sobre la diagonal y correlaciones fenotípicas debajo de la diagonal de rasgos productivos1 de la línea de cuyes Saños (Junín, Perú)
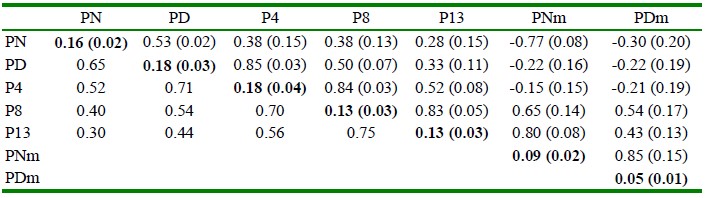
1 Peso al nacimiento (PN), peso al destete (PD, 15 d), peso a la cuarta semana (P4), peso a la octava semana (P8). peso a la décimo tercera semana (P13), efecto genético materno de peso al nacimiento (PNm), efecto materno de peso al destete (PDm)
Error estándar entre paréntesis
Las heredabilidades directas de PN y PD y de los rasgos posdestete de los cuyes de la línea Saños se presentan en el Cuadro 4. Las heredabilidades maternas de PNm y PDm fueron 0.04 ± 0.01 y 0.05 ± 0.02, respectivamente. Las correlaciones genéticas fueron positivas y entre moderadas a altas con valores mínimos de 0.46 y valores máximos de 0.93. La correlación entre el valor genético directo de peso al nacimiento y el valor genético materno de peso al nacimiento fue negativa (-0.14 ± 0.15). Así mismo, la correlación genética entre el valor genético directo y materno de peso al destete fue de -0.58 ± 0.15. Las correlaciones fenotípicas fueron altas y positivas; la correlación fenotípica entre PN y PD fue de 0.63, mientras que las correlaciones fenotípicas entre PD y P4, P4 y P8, P8 y P13, tomaron valores dentro del rango de 0.66 a 0.76.
Cuadro 4 Heredabilidades en la diagonal y en negrita, correlaciones genéticas sobre la diagonal y correlaciones fenotípicas debajo de la diagonal de rasgos productivos1 de la línea de cuyes Mantaro (Junín, Perú)
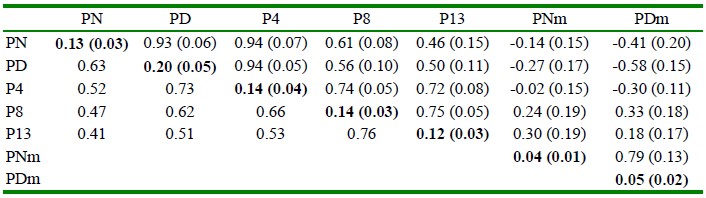
1 Peso al nacimiento (PN), peso al destete (PD, 15 d), peso a la cuarta semana (P4), peso a la octava semana (P8). peso a la décimo tercera semana (P13), efecto genético materno de peso al nacimiento (PNm), efecto materno de peso al destete (PDm)
Error estándar entre paréntesis
Parte de los componentes de varianza mostrados en el Cuadro 5 especifican valores de varianzas de PN debida a la camada de 295.40 y 299.96 para las líneas Saños y Mantaro, respectivamente, y varianzas de PD debidas a la camada de 2050.20 y 2083.02 para las líneas Saños y Mantaro, respectiva- mente. Por lo general, las varianzas debidas a la camada de rasgos posdestete fueron mayores en la línea Saños que en la línea Mantaro. La proporción de la varianza ambiental materna (c2), en ambas líneas fue mayor al peso al destete, con contribuciones del 61 y 64% sobre el fenotipo total de la línea Saños y Mantaro, respectivamente.
Cuadro 5 Componentes de varianza1, ratio de la varianza debida a la camada con respecto a la varianza fenotípica2 de los rasgos productivos3 de las líneas de cuyes Saños y Mantaro (Junín, Perú)
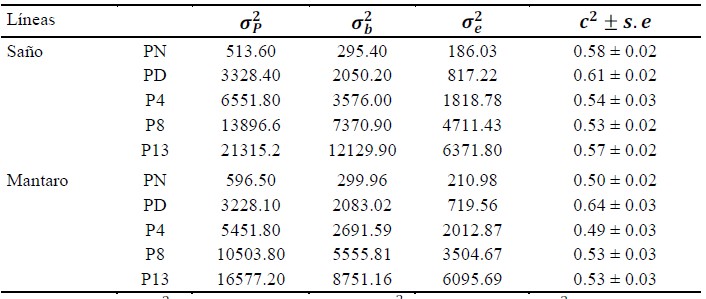
1 Varianza fenotípica (𝜎𝑃 2), varianza debida a la camada (𝜎𝑏 2) y varianza residual (𝜎𝑒 2)
2 Proporción de la varianza ambiental materna (𝑐2)
3 Peso al nacimiento (PN), peso al destete (PD), peso a la cuarta semana (P4), peso a la octava semana (P8), peso a la décimo tercera semana (P13)
DISCUSIÓN
Las consanguinidades media poblacional de la línea Saños (0.67%) y Mantaro (0.18%) para las líneas Saños y Mantaro fueron menores al 2.011% indicado por Solarte et al. (2002), pese a la falta de un esquema de apareamiento adecuado y la reducida población de cada línea Los autores mencionados utilizaron un esquema de apareamiento Poilley adoptado para cuyes por Ortegón y Morales (2002).
Los pesos evaluados y corregidos por efectos fijos como el tamaño de camada, número de parto y otros, en las mismas líneas genéticas son detallados y discutidos en el estudio publicado por Cruz et al. (2021). En el análisis estadístico de los datos no se encontraron diferencias significativas entre las dos líneas para PD y P13, con excepción de mejores valores para PN, P4 y P8 en la línea Saños que en la línea Mantaro, aunque se debe destacar que las diferencias no fueron relevantes.
Las heredabilidades de PN y PD de las dos líneas de cuyes fueron similares a las halladas por Vargas (2015) quien reportó heredabilidades de 0.158 ± 0.036 y 0.24 ± 0.05 para PN y PD de cuyes del genotipo Cieneguilla, respectivamente. Del mismo modo, Rodríguez (2013) reportó heredabilidades de 0.17 ± 0.05 y 0.26 ± 0.07 para PN y PD en el mismo genotipo. Por otro lado, Solarte et al. (2002) obtuvieron heredabilidades de 0.12 ± 0.05 y 0.13 ± 0.05 para PN y PD, respectivamente. Los resultados muestran que las heredabilidades de PN y PD son similares entre ambas líneas. Asimismo, según la escala dada a conocer por Cardellino y Rovira (1987), las heredabilidades de PN de ambas líneas se- rían bajas, mientras que las heredabilidades de PD entre bajas a moderadas.
Las heredabilidades de los tres caracteres posdestete (P4, P8, P13) fueron similares entre ambas líneas y se encuentran en el rango de bajo a intermedio según la escala de Cardellino y Rovira (1987). Solarte et al. (2002), reportaron heredabilidades de 0.32 ± 0.04 y 0.34 ± 0.04 en pesos de cuyes de 4 y 8 semanas, valores superiores a los de esta investigación; posiblemente debido a la inclusión de la varianza ambiental materna dentro de la evaluación para la estima de las heredabilidades de los rasgos mencionados. Las heredabilidades estimadas de P13 para cuyes Saños y Mantaro fueron 0.13 ± 0.03 y 0.12 ± 0.03, respectivamente, y similares al 0.17 ± 0.05 encontrado por Chauca (1997).
Por lo general, heredabilidades altas aseguran el progreso genético; sin embargo, se debe considerar que las correlaciones genéticas podrían afectar este progreso (Falconer y Mackay, 1996). En el presente estudio se encontró que la influencia del efecto de camada sobre los pesos post destete no demuestra reducción de los efectos no aditivos con el incremento de la edad, tal como lo menciona Solarte et al. (2002); quienes mencionaron que es más preciso el uso de las heredabilidades de pesos de posteriores semanas asumiendo que hay reducción de efectos no aditivos en función al incremento de la edad del animal. Por otro lado, las heredabilidades maternas de PN y PD fueron menores a los valores entre 0.11 hasta 0.14 hallados por Cedano et al. (2021) en otras líneas genéticas de cuyes, por lo que es preferible el uso del efecto genético directo (heredabilidades directas) en vez de un efecto genético materno (heredabilidad materna).
Las correlaciones fenotípicas son positivas de magnitud entre moderadas a altas según la escala de Mukaka (2012). En la literatura científica se encuentran correlaciones fenotípicas y genéticas como resultado de la aplicación de diversos modelos mixtos que hace que los resultados difieran entre ellos. Así se tiene que Solarte et al. (2002) reportaron correlaciones genéticas de 0.79 ± 0.13 entre el peso al nacimiento y el destete, en tanto que Chauca (1997) compiló resultados de investigaciones obteniendo correlaciones fenotípica y genética de 0.47 y 0.47 ± 0.1 entre PN y P4, respectivamente, así como valores variados de correlaciones fenotípicas y genéticas entre PN, PD y P13 obtenido de trabajos realizados en varias zonas geográficas del Perú. Por otro lado, Quijandría et al. (1983) reportaron correlaciones fenotípicas de 0.51, 0.38 y 0.46 y correlaciones genéticas de 0.50, 0.24 y 0.61 entre PN y PD, PN y P13 y, PD y P13, respectivamente; en tanto que Hualparuca (2017) en su estudio con cuyes de la línea Saños reportó valores de correlación fenotípicas con rango desde 0.34 hasta 0.81 en machos y desde 0.33 hasta 0.72 en hembras para los diferentes pesos.
La correlación fenotípica entre el PN y PD en ambas líneas fueron numéricamente menores a la correlación entre los demás pesos, posiblemente debido a que los pesos en etapas iniciales suelen ser afectados principalmente por factores exógenos como el tamaño de camada, número de parto de la madre o la competición por el alimento con los adultos a diferencia de etapas posteriores (Burgos et al. 2010; Pajares, 2009). Por lo general, los pesos están correlacionados a lo largo de la curva de crecimiento (Coyne et al., 2016), enunciado que concuerda con los valores de correlación obtenidas. Además, los resultados muestran que las correlaciones fenotípicas y genéticas suelen ser mayores entre medidas próximas y menores entre medidas lejanas.
Varianzas de 342.44 y 409.25 debidas al efecto azar de camada (σ2) para PN fueron reportados por Vargas (2015) y Rodríguez et al. (2013), respectivamente; en tanto que en este estudio fueron de 295.40 y 299.96 para las dos líneas. Así mismo, las varianzas atribuidas por estos autores al efecto al azar de camada para PD fueron de 2054.26 y 1635.40, respectivamente, mientras que en este trabajo se observaron varianzas de 2050.20 y 2083.02. Las proporciones de la varianza de la camada para PN de las líneas Saños y Mantaro representaron aproximadamente el 58 y 50% de la varianza total, mientras que autores como como Solarte et al. (2002), Rodríguez et al. (2013) y Vargas (2015) encontraron contribuciones del 47, 54 y 45.6%, respectivamente; asimismo, estos autores encontraron contribuciones ambientales maternas de 53, 60 y 49 sobre PD, respectivamente, mientras que en esta investigación no superaron el 60%. Los porcentajes obtenidos muestran que hubo una mayor contribución por parte de la varianza ambiental materna sobre PD que sobre PN en ambas líneas. Rodríguez et al. (2013) mencionan que los porcentajes altos de la contribución de la varianza ambiental materna puede ser debido al efecto ambiental temporal de la poza.
CONCLUSIONES
Los pesos vivos de cuyes de las líneas Saños y Mantaro tienen heredabilidades bajas a medias, y las correlaciones genéticas y fenotípicas entre los pesos son positivas y entre moderadas y altas.
La selección podría hacerse en cualquiera de los rasgos evaluados por estar correlacionados positivamente; sin embargo, el peso al destete es el que tiene mayor heredabilidad y podría ser utilizado con fines de evaluación genética.
Las heredabilidades directas de los rasgos evaluados fueron mayores a las heredabilidades maternas en ambas líneas de cuyes, por lo que se debería de considerar al momento de estimar los valores genéticos de los rasgos evaluados.
Las heredabilidades y la ratio de la varianza debida a la camada con respecto a la varianza fenotípica fueron similares entre ambas líneas de cuyes evaluadas.
Las contribuciones debidas a la varianza ambiental materna resultaron tener una alta influencia sobre los rasgos en estudio, con proporciones que van desde el 49 al 64%.

