











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Los animales se adaptan a un medio ambiente a través de cambios genéticos producidos a lo largo de los siglos (FAO, 1996). Las condiciones fisiológicas y patológicas, así como las condiciones ambientales y de manejo pueden influir en el tamaño de los animales. El tamaño contribuye a la estructura, la armonía y el equilibrio del cuerpo, pero también a las características fisiológicas y a los mecanismos de adaptación al medio ambiente (Wainwright et al., 1976).
Una herramienta para cuantificar y analizar la variación morfológica es la morfometría, la cual se refiere al estudio cuantitativo de la variación de las formas biológicas utilizando variables lineales como distancias, ángulos y proporciones que son analizadas por métodos estadísticos multivariados (Marcus, 1990; Bookstein, 1996; Tornese y Nabar, 2013), variables no lineales, como superficies, e incluso, variables de forma, en caso de recurrir a la morfometría geométrica por landmarks o elípticas de Fourier. La producción científica en este campo se ha incrementado en las últimas décadas (Benítez y Püschel, 2014; Parés-Casanova, 2017). Los estudios morfométricos indican si las variables de la forma de la estructura en estudio son o no distintas en las distintas etapas y en qué zona específica se encontrarían esas di- ferencias (Toro et al., 2010).
La caracterización racial basada en la morfometría y morfología mediante la interpretación de datos obtenidos desde diferentes enfoques es una de las primeras fases para la conservación, mantenimiento y conocimiento de los recursos zoogenéticos locales (FAO, 1997). En los animales, las medidas corporales brindan información significativa sobre la estructura morfológica y las habilidades de desarrollo, y representan también los factores más influyentes para determinar la estructura más adecuada de los animales para una eficiencia deseada (Sañudo, 2009; Francesch et al., 2011). Los estudios morfométricos contribuyen al conocimiento en la definición de especie frente a la dispersión geográfica y en estudios de diferenciación sexual (Benítez y Püschel, 2014). A partir de las variables morfológicas cuantitativas se calculan índices que permiten analizar las formas de las regiones para evidenciar la aptitud productiva, funcional y etnológica de las razas y el dimorfismo sexual (Parés-Casanova, 2007a; Gómez et al., 2010), para el desarrollo de programas de mejora genética (Da Costa et al., 2014) y para establecer comparaciones fenotípicas entre animales de distintas razas y a nivel de finca (Salako, 2006).
Los recursos zoogenéticos localmente adaptados constituyen un patrimonio cultural, de investigación, genético y sociocultural de una región. Iberoamérica cuenta con una gran cantidad de animales totalmente adaptados a las condiciones locales cuya producción es sostenible y ecológica en todas sus fases, y cuyo producto tiene un alto valor nutricional (Linares et al., 2011). En las últimas décadas se ha creado una mayor conciencia sobre la importancia de preservar los recursos zoogenéticos, aunque las iniciativas de investigación aún no están totalmente extendidas en la mayor parte del continente americano (Revidatti, 2009; Revidatti et al., 2014).
Los cerdos criollos forman parte de la tradición y cultura de las unidades familiares campesinas (Benítez y Sánchez, 2002). Los cerdos criollos han pasado por cientos de años de selección natural sobre la base de la adaptación a las características ambientales locales (Cortés et al., 2016), que incluyeron la supervivencia y reproducción en áreas geográficas de climas tropicales (Linares et al., 2011).
El cerdo criollo araucano habita en el ecosistema de sabanas inundables de Arauca (Oriente de Colombia). Se cree que descienden de cerdos traídos de España en el siglo XV y que fueron seleccionados para adaptarse a las condiciones locales (Salamanca et al., 2015). Conocidos localmente como «Sabaneros», se manejan en condiciones extensivas recorriendo grandes distancias en condiciones adversas, recibiendo escasos cuidados y prosperando de forma semi-silvestre. Estos cerdos están expuestos permanentemente a perturbaciones ambientales y suministro irregular de alimentos. Poco se sabe sobre su producción y distribución geográfica, lo que merece una atención especial para su estudio. Dada la importancia de es- tos cerdos para la región, y a la escasa información disponible, el objetivo del presente estudio fue contribuir a la caracterización morfométrica para conocer algunas de sus características funcionales y etnológicas que sirvan para elaborar estrategia para su conservación.
MATERIALES Y MÉTODOS
Área de Estudio
La investigación se realizó en el municipio de Arauca, departamento de Arauca, al oriente de Colombia (latitud: 07-04N, longitud: 070-44W, altitud). Es una región de topografía plana y sabana inundable, con una extensión de 587 000 ha, dividida en 56 vere- das (centros de división territorial). La precipitación anual es inferior a 1500 mm, con una época seca o verano (noviembre-abril), y una época lluviosa o invierno (mayo-octubre). La zona presenta una temperatura ambiental de 35 °C en marzo hasta 19 °C en enero, humedad relativa de 65% en marzo y 85% en junio-julio y se encuentra a una altitud de 128 msnm (Arauca, 2020).
Muestra
Se estudiaron 58 cerdos criollos araucanos (27 hembras y 31 machos), con edades entre 4 a 48 meses, localizados en las veredas de Las Monas, Merecure y La Saya. Los cerdos se manejan de forma extensiva tradicional. De cada individuo, se obtuvieron 11 medidas corporales y tres índices, siguiendo procedimientos estándar (Hurtado et al., 2005; Parés-Casanova, 2009; Castro et al., 2012; FAO, 2012):
Perímetro torácico (PT): desde el punto más declive de la cruz para pasar por la región esternal, en el punto situado inmediatamente detrás del codo, y llegar nuevamente al punto de origen o salida, medido con cinta métrica inextensible.
Longitudinal corporal (LCO): desde el punto más craneal y lateral de la articulación del húmero (encuentro) hasta el punto más caudal de la nalga (ilion-isquiático), medido con regla métrica.
Alzada a la cruz (ALC): desde el punto más alto de la cruz hasta el suelo en vertical, medido con regla métrica.
Alzada al esternón (ALES): desde el punto más ventral del cuerpo del esternón a nivel de la cinchera hasta el suelo, medido con regla métrica.
Perímetro rodilla (PRO): perímetro máximo del plano trasversal del carpo, entre el antebrazo y la caña, medido con cinta métrica inextensible
Longitud de cabeza (LCA): desde el punto más culminante del occipital (nuca) hasta el hocico, medido con regla métrica.
Anchura de cabeza (ACA): entre los puntos más salientes lateralmente de los arcos zigomáticos, medido con regla métrica.
Largo de oreja (LORE): desde base de la oreja hasta su vértice, medido con cinta métrica inextensible.
Ancho de oreja (ANORE): desde el punto medio del borde craneal hasta el punto medio del borde caudal, medido con cinta métrica inextensible.
Perímetro caña posterior (PCA): rodeando el tercio medio y el superior del metatarso, medido con cinta métrica inextensible.
Alzada al corvejón (ACO): desde el suelo hasta la porción más alta del corvejón.
Con estas medidas se construyeron tres índices, siendo dos de interés etnológico y uno de interés productivo:
Análisis Estadístico
Se realizó un análisis de estadística descriptiva por sexo que incluían las medidas de tendencia central (media); y la desviación estándar (DE), el coeficiente de variación (CV), máximo (Máx.) y mínimo (Mín.) como medidas de dispersión. De igual forma, para evaluar la relación entre variables morfométricas, se estimaron correlaciones de Spearman.
Con el propósito de medir el efecto ambiental sobre las medidas corporales e índices se realizaron análisis univariados a través de modelos mixtos. La normalidad de las variables se evaluó a través del estadístico de Shapiro-Wilk. Aquellas variables que no se distribuyeron normalmente, fueron transformadas a su log10 para mejorar el ajuste. En el modelo de análisis se consideraron los efectos fijos de vereda, sexo, y la interacción sexo*vereda, mientras que el afecto aleatorio fue el animal dentro de cada vereda. Dado que no todos los animales tenían la misma edad, se incluyó como covariable. Cuando alguno de los efectos fijos presentó diferencias estadísticas, se usó la prueba DMS para la diferenciación de medias con un nivel de significancia del 5%. Todos los análisis se realizaron en el programa SAS v. 9.4 (SAS, 2013).
RESULTADOS Y DISCUSIÓN
Estadísticos Descriptivos
Los resultados de los estadísticos descriptivos por sexo se presentan en el Cuadro 1. Se estima que si en una población de estudio las variables morfológicas presentan un CV inferior al 5% se considera de escasa variabilidad, indicando una gran homogeneidad de la población; si el CV está entre el 5 y 15%, la población presenta un grado de uniformidad medio, y si es superior al 15%, se debe considerar una elevada variabilidad de la muestra estudiada (Herrera et al., 1996; Parés-Casanova, 2007b; Bravo y Sepúlveda, 2010). En este estudio, el CV general se encontró dentro de un rango de 10.9 y 19.9% para las variables LCA y PCA, respectivamente y de 8.9% para IP y 14.7% para ICE, por lo que se puede deducir que la población de cerdos criollos araucanos examinada presenta una uniformidad morfológica media considerable.
El ICE, considerado como el principal índice de diagnóstico racial (Sañudo, 2009) presentó una variabilidad media en el presente estudio. Este índice clasifica a los porcinos en tres tipos étnicos: los troncos asiático y céltico (braquicéfalos, cabezas anchas), y el tronco ibérico (dolicocéfalo, cabezas largas) (Castro et al., 2012; Hernández-Baca et al., 2017). La importancia etnológica de este índice se explica porque su variación no está influenciada por los factores ambientales ni por el manejo dado a los animales (Herrera y Luque, 2009). Según Aparicio (1960), el ICE es muy constante en cada raza y si es inferior a 50 el animal es similar al tipo ibérico. El valor de 37.5% contenido en el estudio puede clasificar a la población de cerdos criollos araucanos como dolicocéfalos o de cabeza alargada, presentando diferencia con los cerdos Pampa Rocha (Montenegro et al., 2014) y de Ecuador (Espinosa, 2016), que son re- portados como mesocéfalos o de cabeza redondeada.
El IP determina la forma (rectangularidad) del cuerpo del animal y requiere para su interpretación un alto nivel de intuición, dado que, a menor valor, el animal se aproxima más a un rectángulo, forma predominante en animales de aptitud cárnica (Sañudo, 2009). El valor encontrado en el presente estudio (83.3%) fue superior al obtenido para el cerdo criollo de Nicaragua (Hernández- Baca et al., 2017) y para el cedro criollo del Perú (Pujada et al., 2018). Lo anterior sugiere que el cerdo criollo araucano se aproximaría a un animal de aptitud cárnica, aunque el reconocimiento por parte de los producto- res es de ser un animal de «alto engrasamiento» por lo que es muy utilizado en la producción de «manteca» (aceite) para el consumo humano.
El ICO estima la conformación de la raza, permitiendo clasificar a los animales en brevilíneos (d»85), mesolíneos (entre 86 y 88) y longilíneos (e»90) (Sañudo, 2009). De acuerdo con estos criterios y según los resultados obtenidos (86.9%), el cerdo criollo araucano se podría clasificar como mesolíneo, con va- lores de longitud y altura relativamente iguales. Los valores del ICE y de ICO fueron inferiores a los calculados para el cerdo criollo de Nicaragua (Hernández-Baca et al., 2017) y para el cedro criollo del Perú (Pujada et al., 2018).
Efectos Ambientales
Las variables de medidas corporales para cada efecto ambiental se presentan en el Cuadro 2. La vereda fue el único efecto que influyó sobre el comportamiento medio de las variables LCO, ALC, ALES, PRO y LCA (p<0.05). De igual forma, la covariable «edad» influyó (p<0.05) para PT, LCO, ALC, ALES, PRO, LCA y ACO, efecto esperado dado el rango de edades evaluadas.
Se observa una amplia variabilidad de algunas medidas corporales e índices en las tres veredas estudiadas (Cuadro 3). Los va- lores medios para la mayoría de las variables estudiadas fueron mayores en la vereda Las Monas (excepción PRO, que fue mayor en Merecure), seguida de La Saya y Merecure. Aunque todas las medidas tienen importancia morfométrica, se resalta las diferencias (p<0.05) en LCO, ALC, ALES, PRO y LCA.
Las diferencias pueden estar asociadas al entorno donde se han venido desenvolviendo los cerdos, vinculados a mecanismos de movilidad para recorrer grandes extensiones por terrenos agrestes y particulares en cada división territorial. El LCA fue otra variable que presentó fluctuaciones entre veredas (p<0.05), particularidad de adaptación que le concede al cerdo la habilidad para la adquisición de alimentos a través del proceso denominado «hociqueo».
Cuadro 1. Estadísticosdescriptivos por sexo de variables corporales (cm) e índices (%) en cerdos criollos Araucanos (Colombia).
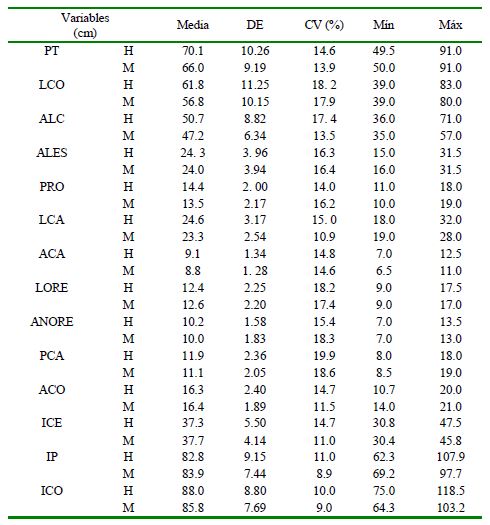
PT: Perímetro torácico; LCO: Longitudinal corporal; ALC: Alzada a la cruz; ALES: Alzada al esternón; PRO: Perímetro rodilla; LCA: Longitud de cabeza; ACA: Anchura de cabeza; LORE: Largo de oreja; ANORE: Ancho de oreja; PCA: Perímetro caña posterior; ACO: Alzada al corvejón; ICE: Índice cefálico; IP: Índice de proporcionalidad; ICO: Índice corporal
Cuadro 2. Influencia de vereda, sexo y edad sobre variables morfológicas en cerdos criollos araucanos
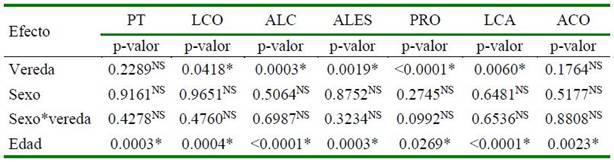
PT: Perímetro torácico; LCO: Longitudinal corporal; ALC: Alzada a la cruz; ALES: Alzada al esternón; PRO: Perímetro rodilla; LCA: Longitud de cabeza; ACO: Alzada al corvejón
*(p<0,05); NS= No significativo
Cuadro 3. Valor medio de las variables corporales (cm) e índices (%) según fuente de variación vereda en cerdos criollos araucanos (Colombia)
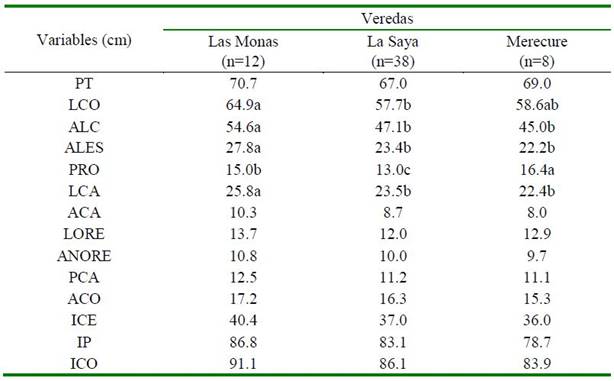
PT: Perímetro torácico; LCO: Longitudinal corporal; ALC: Alzada a la cruz; ALES: Alzada al esternón; PRO: Perímetro rodilla; LCA: Longitud de cabeza; ACA: Anchura de cabeza; LORE: Largo de oreja; ANORE: Ancho de oreja; PCA: Perímetro caña posterior; ACO: Alzada al corvejón; ICE: Índice cefálico; IP: Índice de proporcionalidad; ICO: Índice corporal
Cuadro 4 Valor medio de variables corporales (cm) e índices morfológicos (%) según fuente de variación y sexo en cerdos criollos araucanos (Colombia)
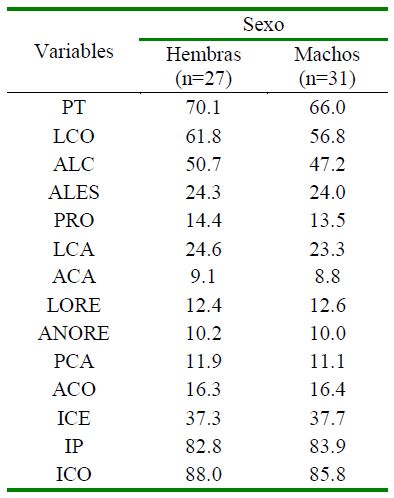
PT: Perímetro torácico; LCO: Longitudinal corporal; ALC: Alzada a la cruz; ALES: Alzada al esternón; PRO: Perímetro rodilla; LCA: Longitud de cabeza; ACA: Anchura de cabeza; LORE: Largo de oreja; ANORE: Ancho de oreja; PCA: Perímetro caña posterior; ACO: Alzada al corvejón; ICE: Índice cefálico; IP: Índice de proporcionalidad; ICO: Índice corporal
Las diferencias morfométricas encontradas entre veredas pueden estar asociadas por el mecanismo de la selección natural que ha ocurrido en cada división territorial, produciendo animales con rasgos morfológicos adaptativos diferentes. Dado que en cada di- visión territorial (vereda) del ecosistema de sabana inundable existen diferentes unidades fisiográficas, este es un factor determinante que contribuye en la diversidad de recursos vegetales para la alimentación y crecimiento de los cerdos. Por ejemplo, en la vereda Las Monas y Merecure existen posiciones fisiográficas denominadas bancos, banquetas, bajo y esteros, y en la Saya predominan bosques de galería; consecuentemente, el mate- rial vegetal disponible para la alimentación de los cerdos es disímil, pues básicamente está dado por raíces de plantas como el Boro (Euchornia crassipes), frutos de Palma Real (Attalea butyracea) y Palma corozo (Acrocomia aculeata), frutos de árboles nativos como el Saman (Pithecellobium saman), Cañafistola (Cassia moschata), Caro Caro (Enterolobium cyclocarpum), Algarrobo (Hymenaea courbaril), Guacimo (Guazuma ulmifolia), y algunos anélidos terrestres (Lumbricus terrestris), entre otros.
Los cerdos en las tres veredas son considerados como dolicocéfalos (ICE). Los cerdos de Merecure tienden a ser más de aptitud cárnica (IP), mientras que los cerdos de las Monas tienden a ser más longilíneos (ICO). Estos valores contrastan con los encontrados por Espinosa (2016) en cerdos criollos ecuatorianos. Dada la variabilidad de las medidas entre veredas se podría inferir que estos cerdos presentan biotipos distintos.
El sexo ni la interacción vereda*sexo fueron fuente de variación significativa (p>005) para las variables analizadas; sin embargo, se observa una tendencia a meno- res valores en machos que en hembras, a excepción del ACA y LORE (Cuadro 4). Estos resultados difieren a lo reportado para el cerdo criollo Alto Andino del Perú donde los machos mostraron tendencias a ser más grandes (Pujada et al., 2018); para el cerdo criollo de Venezuela donde los machos son más altos y las hembras presentan mayor PT (Hurtado, 2006) y con el cerdo criollo Pampa Rocha que presentan indicios de dimorfismo sexual, siendo las hembras mesolíneas (Castro et al., 2012). Esta diferencia es propia de especies poligínicas y puede estar dado a condiciones propias de cada división territorial y con el desempeño reproductivo y materno, puesto que las hembras requieren mayor tamaño para alojar varias crías y asegurar la supervivencia de estas.
Cuadro 5 Correlaciones de Spearman entre medidas corporales e índices (valores estimados y su significancia se encuentran por debajo y encima de la diagonal, respectivamente) en cerdos criollos araucanos (Colombia)
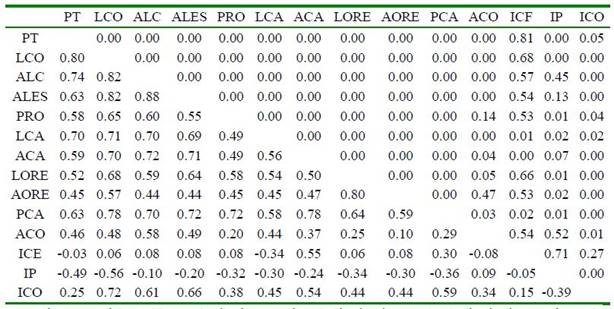
PT: Perímetro torácico; LCO: Longitudinal corporal; ALC: Alzada a la cruz; ALES: Alzada al esternón; PRO: Perímetro rodilla; LCA: Longitud de cabeza; ACA: Anchura de cabeza; LORE: Largo de oreja; AORE: Ancho de oreja; PCA: Perímetro caña posterior; ACO: Alzada al corvejón; ICE: Índice cefálico; IP: Índice de proporcionalidad; ICO: Índice corporal
El ICE fue similar en los machos y hembras (p>0.05), el IP fue mayor en machos y el ICO fue superior en las hembras. Los va- lores de estos índices son similares con los reportados para el cerdo criollo del Chocó (Arredondo et al., 2011), de Nicaragua (Hernández-Baca et al., 2017) y del Alto Andino Perú (Pujada et al., 2018) donde los índices IP e ICE fueron mayores en los machos y el ICO en hembras.
En general, se evidencia poco dimorfismo sexual de tamaño en cerdos criollos araucanos. Este fenómeno también ha sido reportado en cerdos criollos en Guatemala (Jáuregui et al., 2019), en el cerdo Mancha- do de Jabugo (Forero et al., 2000), y en el cerdo criollo de Nicaragua (García, 2016). Pese a que no hay una explicación de las causas del dimorfismo sexual de tamaño, se indica que podrían deberse a restricciones evolutivas, selección natural o selección sexual (Dale et al., 2007). Por otro lado, es un fenómeno poco comprendido respecto a sus causas ontogenéticas y evolutivas (Martínez et al., 2014).
Análisis de Correlación
Los resultados del análisis de correlación de Spearman se muestran en el Cuadro 5. El 47% de los valores (43/91) tuvieron una alta correlación (r>0.5) y con significancia estadística (p<0.05). Las variables más correlacionadas fueron ALC con ALES y LCO (p<0.05), y las menos correlacionadas ACO con AORE (p>0.05). Ninguna medida corporal presentó correlación negativa.
Parés-Casanova (2009), indica que cuando todas las variables están significativamente correlacionadas, una raza responde a un modelo armónico de la estructura, y cuando el número de correlaciones significativas entre las diversas variables ronda el 50%, responde a un modelo medianamente armónico. Para el caso del cerdo criollo araucano, más del 82% de las correlaciones (75/91) fueron estadísticamente significativas (p<0.05), lo que revela que el cerdo criollo araucano es un animal medianamente armónico en su estructura. El modelo armónico también es reportado para el cerdo criollo de Nicaragua (García, 2016), aunque no menciona el número de correlaciones significativas.
El IP presentó correlación negativa con todas las variables, el ICO se correlacionó negativamente con el IP y el ICE presentó correlación negativa con PT, LCA y ACO, lo que puede estar asociado con el aumento o disminución de las variables que los conforman.
CONCLUSIONES
Se observa una variabilidad media en las medidas corporales estudiadas que se asocia con una mediana armonía del ani- mal.
La amplia variabilidad de las medidas según las veredas podría indicar que en los cerdos araucanos se presentan biotipos distintos.
El dimorfismo sexual de tamaño no es significativo, aunque las hembras tienden a presentar valores mayores en las me- didas corporales.
La variabilidad morfométrica da indicios para establecer futuros programa de conservación.

