











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
INTRODUCCIÓN
La población mundial de llamas es cercana a tres millones de cabezas (Quispe et al., 2009), siendo Bolivia el primer productor con el 65% de la población mundial, seguido por Perú con más de 740 mil cabezas (INEI, 2013), población que disminuyó en cerca del 50% en comparación con la población existente 15 años atrás, lo que se debería a la disminución de su uso como animal de transporte e intercambio (Iñiguez y Alem, 1996), así como al poco hábito de consumo de su carne, al cambio climático y a la baja valoración de su fibra.
La variedad K’ara (conocida también como Ccara, Q´ara, Cara-Sullo, Pelada o Carguera) y la variedad Chaku (conocida también como T’ampulli, Choco, Lanuda o Tapada) son dos ecotipos de llamas (Bonacic, 1991; Stemmer et al., 2005) bien diferenciados, aunque el cruce entre ambos produce un tipo de llama conocido como «intermedio» (Stemmer et al., 2005). La llama K’ara se caracteriza por estar desprovista de fibra o pelo en la cara, cuello, extremidades y barriga, es utilizada como animal de carga, produce escasa cantidad de fibra gruesa y su principal finalidad es la producción de carne, mientras que la llama Chaku tiene fibras que cubren todo el cuerpo, con un vellón muy denso y su mayor uso es para producción de fibra. Por otro lado, se tiene las llamas de Argentina que exhiben características particulares en torno a su morfología, color de capa y tipos de vellón (Hick et al., 2014), y las llamas de Marcapomacocha, Perú, que se caracterizan por ser muy grandes, manteniendo homogéneamente el color del guanaco (Cano et al., 2012).
Las llamas son generalmente animales de doble capa de fibras. La capa externa conformada por fibras gruesas (llamadas también «cerdas») y la capa interna formada por fibras finas (conocidas como «down») que son de buena calidad (Rodríguez, 2006; Quispe et al., 2015); por tanto, su uso para la confección de prendas finas dependería del descerdado (Sacchero et al., 2017), mientras que las fibras gruesas tendrían un uso potencial en la elaboración de prendas artesanales (Pinares, 2014).
Estudios sobre fibra de llamas han sido realizados principalmente en el ecotipo Chaku. Así, Martinez et al. (1997) en Bolivia encontraron una media de diámetro de fibra (MDF) de 25.5 y 40.8 µm y una longitud de mecha (LM) de 5.4 y 9.0 cm para las fibras finas y gruesas, respectivamente, mientras que Laime et al. (2016) reportaron una MDF de 21.8 µm en fibras finas después del descerdado. Asimismo, Quispe-Ccasa et al. (2020), sin diferenciar fibras finas y gruesas encontraron una MDF de 24.3 µm y LM de 9.5 cm, en tanto que Sacchero et al. (2017) reportaron una MDF de 22.6 µm y LM de 13.2 cm para llamas de Rio Negro, Argentina, y Cóndor et al. (2022) encontraron una MDF bastante gruesa (39.2 y 37.3 µm para machos y hembras, respectivamente). En este sentido, diversos autores reportan resultados variables para llamas (Rossanigo et al., 1997; Quispe-Huanca, 2014; Mancilla, 2017).
En el crecimiento de fibra, Ayala et al. (2018), sin diferenciar fibras finas y gruesas, encontraron una LM al nacimiento de 2.7 y 3.0 cm, creciendo hasta el año de edad a 13.6 y 12.2 cm, para llamas K’aras y Chaku, respectivamente, lo cual significaría una tasa de crecimiento mensual de 0.9 y 0.8 cm, respectivamente; resultado similar a las llamas de Catamarca, Argentina (Mueller et al., 2010). Por otro lado, en referencia al rendimiento al descerdado se ha registrado 68.6±2.5% (Quispe et al., 2015) y cerca del 80.0% (Cochi, 1999).
Las llamas K’ara han sido evaluadas principalmente en torno a la producción de carne (Mamani-Linares et al., 2014) y como medio de transporte (Iñiguez y Alem, 1996). En relación a la fibra, Vilá (1996) refiere que carecen casi por completo de interés textil, debido a que no posee mechas diferenciables, aunque abundantes fibras primarias. Sin embargo, Pari (1993), Mancilla, 2017) y Cóndor et al. (2022) determinaron en forma general el diámetro medio de las fibras indicando que la fibra de llamas K’ara tienen similares diámetros que las llamas Ch’acu, mientras que Alendez (2021) y Poma (2018) evaluaron el diámetro de las fibras antes y después del descerdado. Sin embargo, estudios sobre las características y relaciones de diámetro, longitud y rinde al descerdado de fibras finas y gruesas son escasos, no obstante que las fibras finas podrían tener gran potencial textil, (Iñiguez et al., 1998).
Con base a lo expuesto, los objetivos del presente trabajo fueron conocer las características de calidad, crecimiento y rinde al descerdado de fibras finas y gruesas, así como, determinar el efecto de diversos factores sobre dichas características de las fibras de llamas K´ara criadas en condiciones altoandinas de Perú.
MATERIALES Y MÉTODOS
Localización, Animales y Registros
Obtuvieron los registros de 69 llamas K´ara, siendo 49 del Centro de Investigación de Camélidos Sudamericanos Lachocc (CICS-Lachocc) de la Universidad Nacional de Huancavelica (UNH) y 20 de Huaccoto, Comunidad Campesina de San Miguel de Mestizas (CC Mestizas), distrito de Cotaruse, departamento de Apurímac, Perú. El CICS-Lachocc está ubicado a una altitud de 4490 msnm, mientras que la CC Mestizas se encontraban a una altitud de 4400 msnm. La región tiene temperaturas que varían desde 5 hasta 18 °C y una precipitación pluvial de 752 mm/año.
En llamas del CICS-Lacchoc se tomaron muestras de fibra de 5 g de tres zonas corporales de cada animal (Paleta = P, Costillar = C y Nalga = N), mediante corte de las fibras utilizando una tijera. Las muestras se colocaron en bolsas de papel donde anotando la zona corporal (P, C, N), edad (1 a 12 años) y sexo (macho, hembra). También se registró el peso vivo de 35 llamas con una balanza de plataforma (± 50 g). Asimismo, se logró esquilar a las 49 llamas por primera vez (anteriormente no habían sido esquilados), en forma manual, con tijera conocida como «lapiaco». Los vellones fueron pesadas con una balanza tipo reloj a fin de registrar el peso de vellón.
En las llamas CC Mestizas se evaluó la longitud de fibras finas («down») y fibras gruesas («cerdas») mediante una regla milimétrica metálica, considerando cinco mediciones en cada animal y tipo de fibra (finas y gruesas). Lu ego se tomaron muestras de fibras (22.9 ± 2.5 g, promedio ± error estándar) cortándolas con una tijera a nivel de la piel, registrando la fecha de muestreo, sexo, color y categoría etaria (cría, juvenil y adulto). A los 106 días de volvió a evaluar la longitud de las fibras finas y gruesas, siguiendo el mismo procedimiento y de la misma zona corporal a fin de obtener la tasa de crecimiento mensual.
Evaluación de Fibras Finas y Gruesas
Las muestras de fibras de las llamas del CICS Lachocc fueron descerdadas de acuerdo con el procedimiento descrito por Frank et al. (2012), el cual está basado en la utilización de los dedos en lugar de fórceps, con 5 a 8 movimientos de descerdado por mechón. Además, se determinó la media del diámetro de fibra (MDF), el coeficiente de variación de la MDF (CVMDF) y el porcentaje de fibras menores a 30 micras (Fib<30mic), tanto de fibras finas como de gruesas, utilizando el FIBER EC de acuerdo a la metodología indicada por Quispe et al. (2017).
Las muestras de la CC Mestizas también fueron sometidas al proceso del descerdado manual, de acuerdo con el procedimiento anteriormente indicado. Para una mejor separación de las cerdas se colocó como base un papel contrastante con el color de fibra a descerdar (negro para fibra blanca y blanco para fibra de color) encima de la mesa. De cada muestra descerdada se obtuvieron tres componentes: fibra fina, gruesa e impurezas (esta última compuesta por fibras muy cortas, restos vegetales y tierra), los cuales fueron pesados en una balanza analítica.
Análisis Estadístico
Los datos obtenidos se procesaron mediante el paquete estadístico R v. 3.1.1. (R Core Team, 2020). Se inició con un análisis exploratorio univariado, pasando luego por la prueba de Shapiro y de Barlett para evaluar la normalidad de los residuos estandarizados y la homogeneidad de varianzas de los datos. Datos correspondientes a Fib<30mic fueron sometidos a una transformación trigonométrica; pero para la presentación de resultados se realizaron las reconversiones respectivas.
Para evaluar las características de calidad de la fibra fina y gruesa, así como del peso vivo y peso de vellón se calcularon los principales estadísticos mediante el paquete «psych» de R. Para la evaluación del efecto sexo, zona de muestreo y su interacción, así como de las covariables edad, peso vivo y peso de vellón sobre las características de calidad de la fibra fina y fibra gruesa se utilizó un modelo aditivo lineal con covariables. Asimismo, para evaluar el efecto del sexo, color de vellón y categoría etaria (crías, tuis y adultos) sobre características del crecimiento mensual de los dos tipos de fibras y sobre el rinde al descerdado de fibras finas se usó un modelo aditivo lineal, haciendo posteriormente un análisis de varianza de tipo III por tratarse de datos desbalanceados. Las diferencias de medianas se visualizaron mediante gráficos de cajas y bigotes, mientras para las interacciones se utilizaron gráficos de medias. Adicionalmente, para evaluar las relaciones entre dos variables (que resultaron significativos en el modelo aditivo lineal) se elaboraron gráficos de puntos, mostrando sus correlaciones de Pearson y ecuaciones de regresión simples, dentro de los gráficos.
RESULTADOS
Peso vivo, peso de vellón, fibra fina, fibra gruesa, crecimiento de fibra/mes y rinde al descerdado de llamas K´ara
Las características productivas y tecnológicas de la fibra se muestran en el Cuadro 1. El peso vivo promedio fue relativamente alto 87.9±4.4 kg (promedio ± error estándar), encontrándose inclusive llamas por encima de los 150 kg; sin embargo, la producción de fibra fue baja (1175.1±96.6 g), incluyendo vellones menores a 500 g. Por otro lado, las fibras finas presentaron una buena calidad, reflejado en su diámetro (21.3±0.3 µm), coeficiente de variación (18.9±0.2%) y Fib<30mic (95.7±0.5%), encontrando animales con MDF menores a 18 µm, con baja variabilidad entre las fibras (<16%) y con Fib<30mic cercanos al 100%.
Cuadro 1. Estadísticos del peso vivo, peso de vellón y características tecnológicas de la fibra de llamas K´ara (Apurímac, Perú)
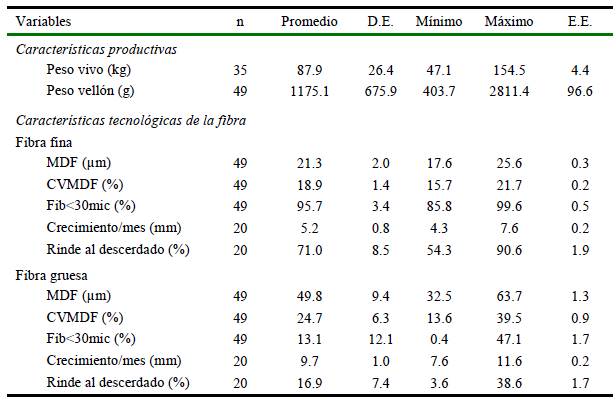
MDF: Media del diámetro de fibra; CVMDF: Coeficiente de variación de la MDF; Fib<30mic: Porcentaje de fibras menores a 30 micras; D.E.= desviación estándar; E.E. = error estándar
También se encontró que, aunque las fibras finas crecen más lentamente que las fibras gruesas (5.2±0.2 y 9.7±0.2 mm/mes, respectivamente), tienen un buen rendimiento al descerdado (71.0±1.9), habiéndose detectado inclusive muestras con un rendimiento mayor al 90%, aunque también se presentaron vellones con rendimiento de fibras finas cercanas al 50%. Alrededor del 13% corresponde a impurezas y materias extrañas (principalmente tierra) en el vellón, que son obtenidos como productos del proceso manual de descerdado.
Edad, peso vivo, peso de vellón, sexo, zona e interacción zona x sexo de las fibras finas y gruesas sobre la MDF, CVMDF y Fib<30mic de llamas K´ara
El efecto de la edad, peso vivo, peso de vellón, sexo, zona e interacción sexo x zona de las fibras finas y gruesas sobre las características tecnológicas de la fibra de llamas K´ara se presenta en el Cuadro 2. En las fibras finas no hubo efecto de las variables e interacciones sobre la MDF, CVMDF y Fib<30 micras, a excepción del peso vivo (p<0.001) sobre MDF y sobre Fib<30 micras (<0.001). En fibras gruesas tampoco se observaron efectos de la edad, sexo, zona corporal e interacción sexo*zona sobre el MDF, CVMDF y Fib<30mic; sin embargo, el peso vivo tuvo efecto significativo (<0.001) sobre la MDF, CVMDF y %Fibras<30mic (<0.001), y el peso de vellón sobre la MDF.
Cuadro 2. Efectos de la edad, peso vivo, peso de vellón, sexo, zona e interacción sexo x zona de fibras finas y gruesas sobre las características tecnológicas de la fibra de llamas K´ara criadas en CICS-Lachos (n=49)
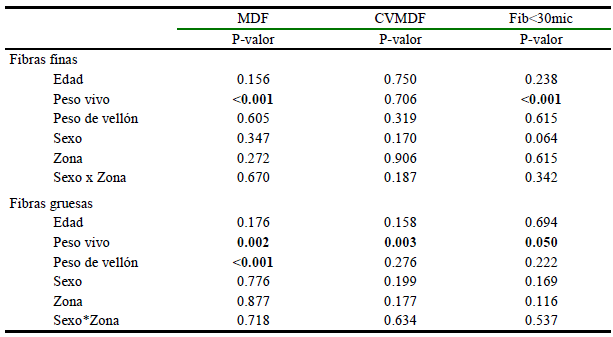
MDF: Media del diámetro de fibra; CVMDF: Coeficiente de variación de la MDF; Fib<30mic: Porcentaje de fibras menores a 30 micras
Regresión del peso vivo y peso de vellón de fibras finas y gruesas sobre sobre la MDF, CVMDF y Fib<30mic de llamas K´ara
Los estadísticos de las regresiones peso vivo y peso de vellón sobre la MDF, CVMDF y Fib<30mic de fibras finas y gruesas de llamas K´ara se muestran en el Cuadro 3. El peso vivo tiene una relación lineal directa y significativa (p<0.001) con la MDF, pero inversa con el Fib<30mic para el caso de las fibras finas; sin embargo, los coeficientes de regresión (bxy) son bajos (0.034 µm y -0.061 µm, respectivamente). Las demás evaluaciones no mostraron relaciones significativas (p>0.05). En el caso de las fibras gruesas solo se encontró que el peso de vellón tiene relación inversa con la MDF y directa con el Fib<30mic; sin embargo, los (bxy) son igualmente muy bajos (-0.006 µm y 0.007 µm, respectivamente). Estos resultados indican que, por cada 10 kg de incremento del peso vivo de las llamas, la MDF se incrementaría apenas en 0.34±0.08 µm, mientras que el Fib<30mic disminuiría en 0.61%, en las fibras finas de las llamas; en tanto que por cada 100 g de incremento del peso de vellón, la MDF de las cerdas disminuiría entre 0.99 y 0.28 µm, pero incrementaría el Fib<30mic entre 0.20 y 1.2%.
Cuadro 3. Regresiones del peso vivo y peso de vellón sobre la MDF, CVMDF y Fib<30mic de fibras finas y gruesas de llamas K´ara, criadas en CICS-Lachoc
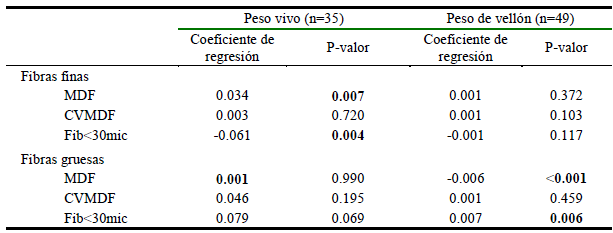
MDF: Media del diámetro de fibra; CVMDF: Coeficiente de variación de la MDF; Fib<30mic: Porcentaje de fibras menores a 30 micras
Efecto del sexo, color, categoría etaria e interacciones sobre el crecimiento mensual de fibras finas y gruesas, y rinde de fibras finas al descerdado
Los efectos del sexo, color, categoría etaria y las interacciones entre los factores en estudio, sobre el crecimiento mensual de fibras finas y gruesas y sobre el rinde de fibras finas al descerdado se muestran en el Cuadro 4. En las fibras finas solo el sexo tuvo efecto sobre el crecimiento mensual (p=0.026) y el rinde al descerdado (p=0.039), observándose que las hembras tienen mayor tasa de crecimiento de fibras finas, pero a la vez menor rinde al descerdado de fibras finas en comparación a los machos (Figura 1, parte superior). De otro lado, el sexo (p=0.002), la interacción sexo x categoría (p=0.017) y sexo x categoría (p=0.004) tuvo efecto sobre el crecimiento de las fibras gruesas. En la Figura 1 (parte inferior) se observa que en llamas con fibras de colores claros los machos tendrían un menor crecimiento comparado con las hembras, pero que en colores oscuros esta diferencia es bastante menor. Asimismo, en crías y animales adultos, los machos tendrían un menor crecimiento de las fibras gruesas comparado a las hembras, ocurriendo lo contrario en animales jóvenes.
Cuadro 4. Efecto del sexo, color, categoría etaria e interacciones sobre el crecimiento mensual de fibras finas, gruesas y sobre el rendimiento al descerdado manual de fibras en llamas K´ara
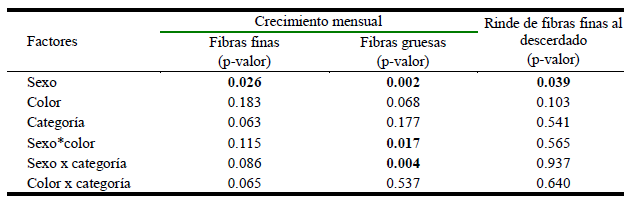
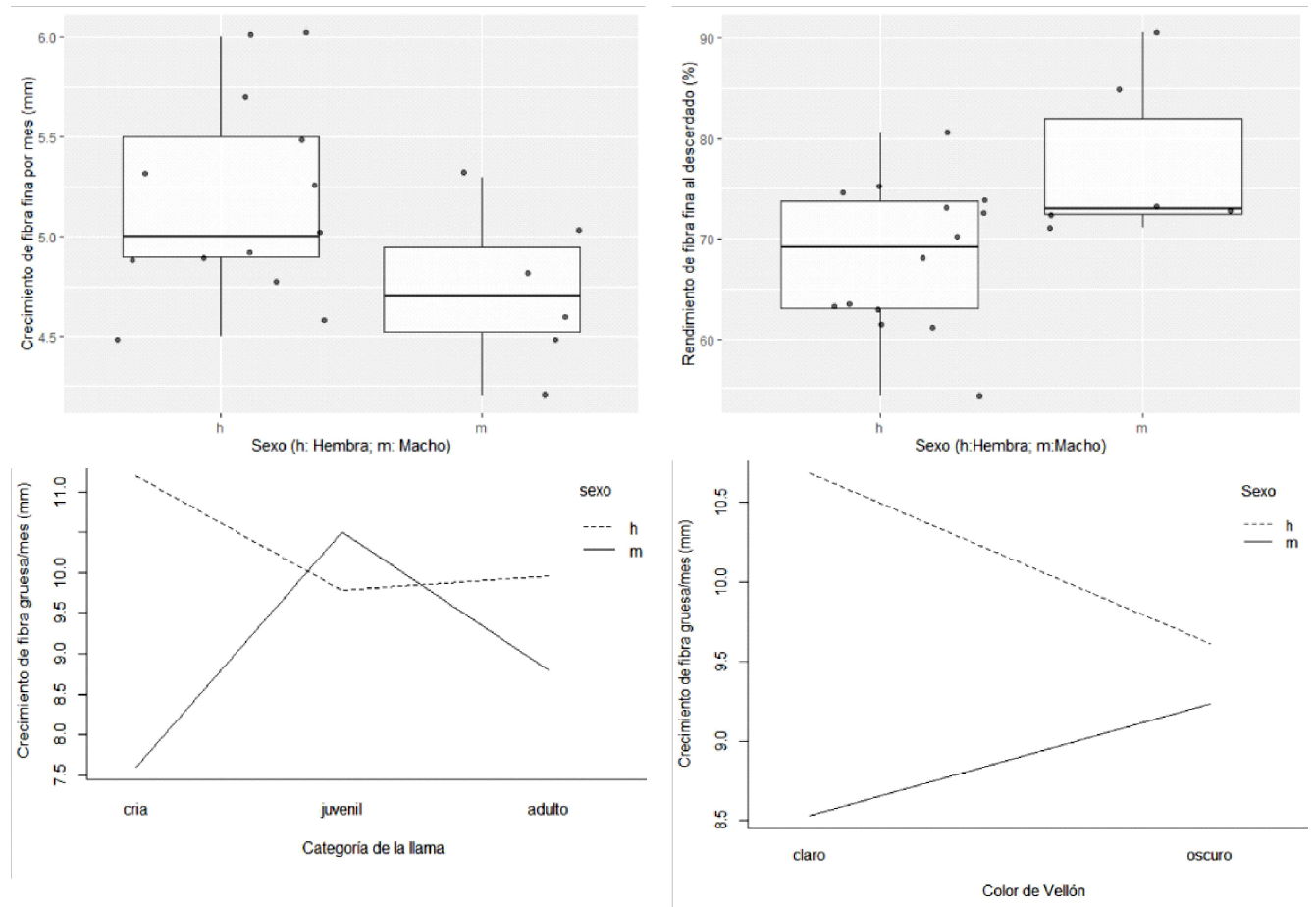
Figura 1. Parte superior: Gráfico de cajas y bigotes por sexo, del crecimiento por mes de fibras finas y rendimiento de fibras finas al descerdado. Parte inferior: Gráfico de medias mostrando interacción entre sexo con color de vellón y categoría, con respecto al crecimiento de fibras gruesas de llamas K’ara
Relaciones entre características productivas y de calidad de fibras de llamas K´ara
La Figura 2 muestra las relaciones lineales bivariadas de peso vivo y peso de vellón con la MDF, CVMDF y Fib<30mic en fibras finas y gruesas. El peso vivo tuvo relación directa con la MDF e inversa con el arcoseno del Fib<30mic en fibras finas, con una correlación mediana de 0.50 entre ellos para ambos casos. Las correlaciones para las demás relaciones bivariadas no fueron significativas.
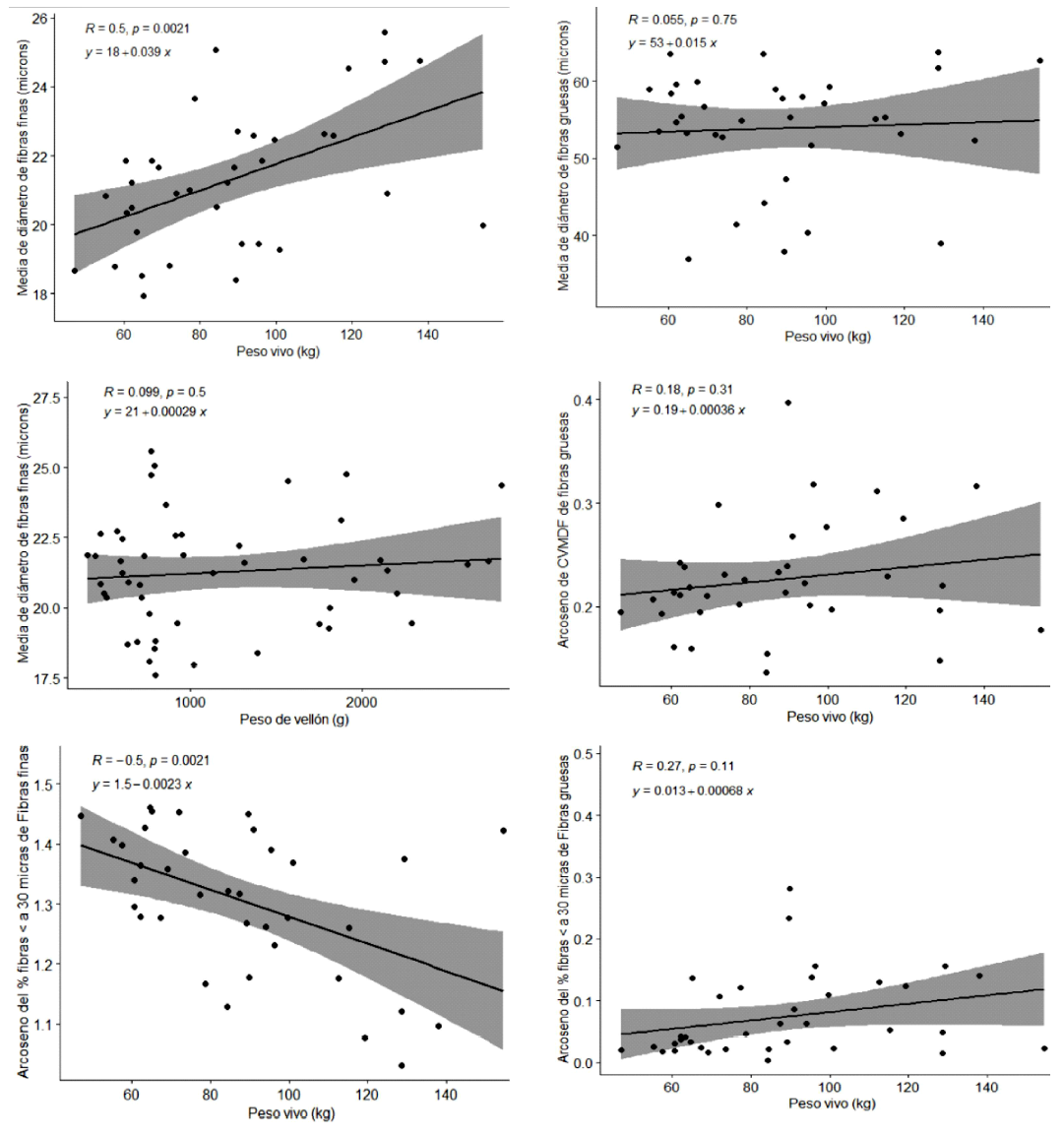
Figura 2. Gráfico de puntos del peso vivo con media del diámetro de fibra (MDF); MDF de fibras finas, MDF de fibras gruesas, arcoseno del Coeficiente de variación de la MDF (CVMDF) de fibras gruesas, arcoseno del porcentaje de fibras menor a 30 µm de fibras finas y gruesas, y gráfico entre el peso de vellón y MDF de fibras finas de llamas K’ara. Se presenta, además. los coeficientes de correlación de Pearson y ecuaciones de regresión
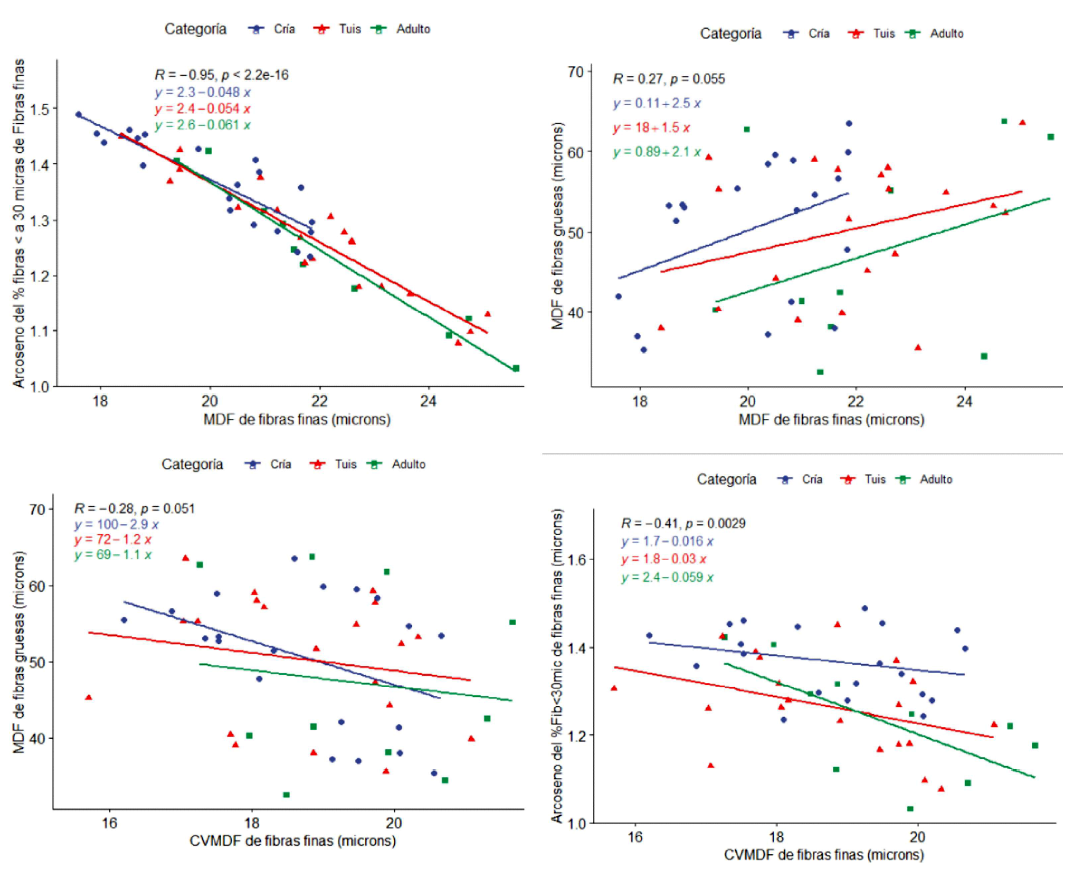
Las relaciones bivariadas entre características de calidad de fibras finas y gruesas se muestran en la Figura 3. Se observa que las pendientes resultan ser muy similares para crías, tuis y adultos cuando se relacionan la MDF y el arcoseno del Fib<30mic, encontrándose una correlación alta (r=0.95); sin embargo, cuando se relacionan la MDF de fibras finas y gruesas, las pendientes para crías y animales adultos son más altas que para tuis, mientras que cuando se relaciona el CVMDF de fibras finas con la MDF de fibras gruesas, la relación es de tipo inversa, obteniéndose una pendiente más pronunciada en crías (coeficiente de regresión = -2.9), en comparación a tuis y animales adultos (coeficientes de regresión = -1.2 y 1.1, respectivamente). Finalmente, cuando se relaciona el CVMDF y arcoseno de Fib<30mic, la mayor pendiente se obtiene en animales adultos (coeficiente de regresión = 0.06), con un coeficiente de correlación de 0.41.
DISCUSIÓN
El peso de vellón obtenido para las llamas K´ara (1175.1±96.6 g) es menor (1.71 kg) a lo reportado por Ayala et al. (2018) para llamas de 21 meses de edad, no habiendo otros valores en la literatura con fines de comparación. Si se considera el peso vivo (87.9±26.4 kg), se debería esperar mayor peso de vellón, tal como se observa en alpacas y ovinos que producen mayor cantidad de fibra con relación al peso vivo. La baja producción de fibra podría deberse a que la especialidad productiva de estas llamas es la carne y la fibra queda como un producto secundario (Bonacic, 1991; Stemmer et al., 2005).
La MDF (21.3±2 µm) registrada en el presente trabajo es inferior al encontrado por Condor et al. (2022), quiénes obtuvieron valores cercanos a 38 µm; así como a los valores reportados por Maquera (1991) y Martinez et al. (1997) con MDF para fibras finas de 24 y 25 µm. No obstante, Ayala et al. (2018), Poma (2018) y Alendez (2021) encontraron MDF de 21-22 µm, similares a los del presente estudio. La MDF, CVMDF y Fib<30mic encontradas en el estudio permite indicar que se podrían confeccionar prendas de vestir de excelente calidad en la industria textil. Por otro lado, el CVMDF (24.7±6.3%) encontrado en fibras gruesas es inferior al 48.4%, reportado por Mancilla (2017); sin embargo, esta enorme diferencia podría deberse a la evaluación de este parámetro utilizando solo fibras gruesas, lo que indica que, al evaluar en forma independiente fibras finas y gruesas, se obtienen fibras con buena homogeneidad, que resulta importante dentro de los procesos textiles.
El resultado en torno a la tasa de crecimiento de fibras gruesas es ligeramente superior al reportado por Mueller et al. (2010) en llamas argentinas y Ayala et al. (2018) en llamas K´aras bolivianas, quienes encontraron una tasa de crecimiento entre 9 y 8 mm/mes. Esta diferencia puede deberse al genotipo, alimentación y, en general, al medioambiente. Lamentablemente, no se pudo encontrar reportes sobre la tasa de crecimiento de fibras finas, pero considerando el resultado obtenido (5.2±0.8 mm/mes), se podría deducir que se tendría un crecimiento de más de 60 mm al año, valor que supera los 50 mm de longitud mínima necesaria para industrializar la fibra en el sistema de peinado, que es el mejor sistema para producir productos textiles finos y de alta calidad.
Por otro lado, se encontró un mayor crecimiento de las fibras gruesas (9.7 cm) que de las fibras finas (5.2 cm), lo cual se debería al gen FGF5 (factor de crecimiento de fibroblastos) que controla el ciclo y afecta la longitud del crecimiento de las fibras. El gen FGF5 tiene dos transcripciones: una de longitud completa y otra de longitud corta, que carece del exón 2. La forma completa induce la fase catágena, una función que es antagonizada por la forma corta que prioriza la fase anágena, de tal modo que mutaciones de pérdida de función en el gen FGF5 han sido asociados con fenotipos de pelo largo en algunas especies, incluidos los camélidos (Anello et al., 2022). De otro lado, es preciso indicar que la presencia de fibras gruesas en los vellones de las llamas representa una adaptación específica para que dichos animales puedan prosperar en ambientes duros, pues ellas son las encargadas de proteger la capa interna subyacente, reducir la pérdida de calor corporal y actuar como una barrera física contra el viento (Gerken, 2010).
A pesar de que las llamas K´ara no tienen la especialidad productiva de fibra como las llamas Chaku, el rendimiento al descerdado de fibras finas es relativamente alto (71%), lo cual se incrementaría a 81.6% si no se considera el contenido de las impurezas. Cochi (1999) y Ayala et al. (2018) encontraron, en forma similar, rendimientos de 80.0 y 81.8%, respectivamente, pero que resultan inferior al rendimiento en llamas Chaku. Sin embargo, con base a los resultados obtenidos se podría afirmar que las llamas K´ara tienen alta proporción de fibras finas de buena calidad textil, la cual no es aprovechada adecuadamente por la industria textil (Stemmer et al., 2005). Uno de los inconvenientes para obtener fibras finas de llamas es el alto costo económico del descerdado manual, cuya producción de fibra fina/h puede ser tan baja como de 10 g/persona/h (Quispe et al., 2015). Sin embargo, esta baja producción podría solucionarse a corto plazo mediante el descerdado utilizando máquinas Minimills (Sacchero et al., 2017) o en el largo plazo mediante selección genética.
La falta de efecto del sexo sobre MDF, CVMDF y Fib<30mic ha sido reportada (Martínez et al., 1997; Siguayro, 2009; Mancilla, 2017). De otro lado, la falta de efecto de la zona corporal (P, C y N) sobre las características de calidad de la fibra de llama resulta similar a los resultados encontrados en alpacas (Huanca et al., 2006), demostrándose homogeneidad de la MDF, incluyendo las variaciones en las distintas zonas corporales, lo cual favorecería enormemente la calidad textil de las prendas de vestir hechas con fibras finas de llamas K´ara. Estas podrían lograr un buen precio en el mercado por ser un producto exótico. Asimismo, el hecho de no encontrar efecto significativo de la edad sobre la MDF podría ser consecuencia que los animales no habían sido esquilados anteriormente.
El hallazgo del efecto del sexo sobre el crecimiento mensual tanto de las fibras finas como de gruesas y sobre el rinde de fibras finas al descerdado, resulta diferente a lo hallado por Poma (2018) quien encontró longitudes de fibra similares entre llamas (K’ara y Chaku) machos (8.9 cm) y hembras (8.7 cm), así como por Castellaro et al. (1998) y Quispe et al. (2014), quienes no encontraron efecto del sexo sobre la tasa de crecimiento de las fibras de alpaca.
El efecto y relación directa del peso vivo sobre la MDF encontrado indica que a mayor peso vivo mayor MDF de las fibras finas y gruesas; sin embargo, este efecto resultaría ser mayor en fibras finas que en fibras gruesas, pues el bxy para fibras finas es mayor y significativo, comparado al de fibras gruesas, cuyo bxy no es significativo. Estos resultados se deberían a la mayor superficie de piel que tienen los animales con mayor peso vivo, lo que permitiría un mayor incremento de nutrientes que repercuten en el incremento del diámetro de las fibras (Adams y Cronje, 1999; Wuliji et al., 2000).
La relación inversa entre la MDF de fibras gruesas y el peso de vellón encontrada resulta particular y contradictoria con lo reportado en llamas por Frank et al. (2011), quienes reportan una correlación positiva de 0.36 (p=0.04) entre estas variables. Asimismo, difiere los hallazgos de otros autores en fibras de alpacas (Wuliji et al., 2000; Quispe et al., 2009) quienes refieren que a mayor peso de vellón existe una mayor MDF. Estas diferencias podrían deberse a la metodología utilizada, pues Frank et al. (2011) no separaron fibras finas y fibras gruesas.
CONCLUSIONES
Las fibras finas de llamas K’ara s presentan una media del diámetro de fibra (MDF) de 21.3±0.3 µm que a la vez exhiben alta homogeneidad expresado con un coeficiente de variación de la MDF (CVMDF) de 18.9%, lo que refleja una buena calidad, favorable para la industria textil.
Las fibras finas tienen una menor tasa de crecimiento que las fibras gruesas (5.2±0.2 y 9.7±0.2 mm/mes, respectivamente), con un rendimiento al descerdado de más de 70%.

