Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología Aplicada
versión impresa ISSN 1726-2216
Ecol. apl. v.3 n.1-2 Lima ene./dic. 2004
ARTÍCULO ORIGINAL
Invasión del matorral por Genista hystrix en una dehesa salmantina (España)
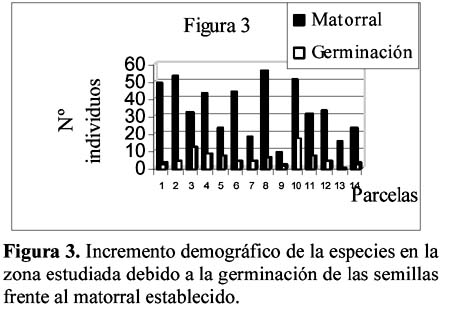
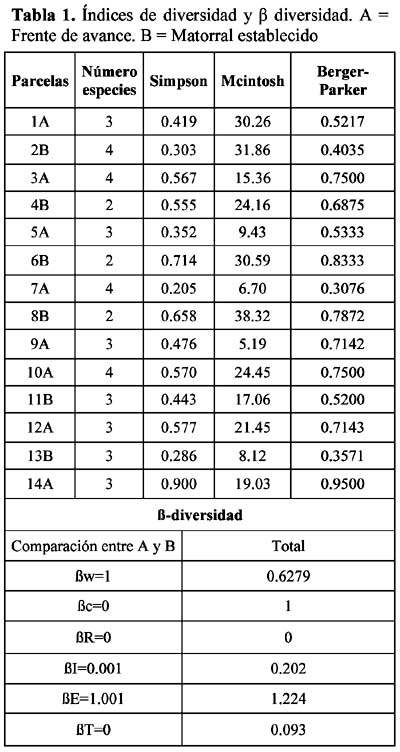
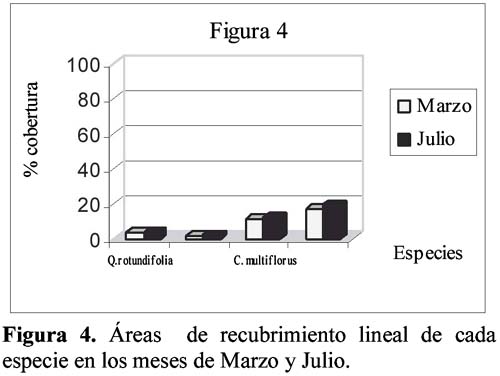
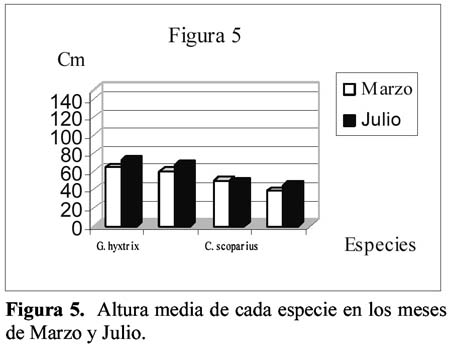
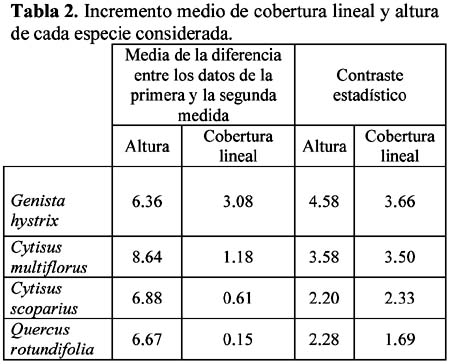
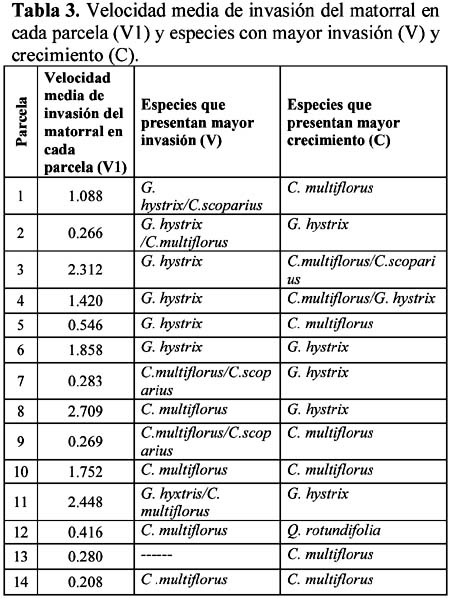
Study of a shurb invasion in a salamancan dehesa Francisco Caro Domínguez1; María Ángeles Sánchez Anta2 y Teresa Tarazona Lafarga3 1 Departamento de Botánica. Facultad de Biología. C/ Licenciado Méndez Nieto, s/n. Campus Miguel de Unamuno. Universidad de Salamanca 37007. Salamanca. (España). Correo electrónico: fcarodomínguez@hotmail.com Resumen Se ha realizado un estudio de la invasión del matorral de Genisto hystricis-Cytisetum multiflori en una dehesa salmantina (bosque de quercíneas de baja densidad) mediante la metodología de transectos y cuadrados permanentes. Los resultados nos indican que, de todas las especies de este matorral, Genista hystrix Lge es, por su velocidad de crecimiento y fenomorfología, la especie invasora predominante en dicha formación esclerófila. Esta especie está invadiendo el pastizal de la dehesa, aproximadamente, 2.26 m2 por año. Palabras clave: Dinamismo, esclerófilo, matorral, dehesa, Salamanca, especies invasivas, Genista hystrix Lge., Cytisus multiflorus (L'Hèr.) Sweet., Cytisus scoparius (L.) Link, Quercus rotundifolia Lam. Abstract The invasion of Genisto hystricis-Cytisetum multiflori shrubs in a Salamancan "dehesa" (Quercus forest with low density), was studied using permanent quadrat and transect methodology. The results indicate that, of all shrub species, Genista hystrix Lge. is the predominant invading species in this sclerophyllous formation, due to its growth rate and phenomorphology. The overall invasion rate of this shrub in the "dehesa" has been calculated to be approximately 2.26 m2 per year. Key words: Dynamics, sclerophyllous, shrubs, "dehesa", Salamanca, invasive species, Genista hystrix Lge., Cytisus multiflorus (L'Hèr.) Sweet., Cytisus scoparius (L.) Link, Quercus rotundifolia Lam. Introducción Se puede definir una dehesa como un bosque de quercíneas de baja densidad, con un suelo relativamente limpio de matorral, donde crece el pastizal. En la dehesa salmantina es Quercus rotundifolia Lam. la especie predominante. Esta especie ocupa lugares caracterizados por un clima continental seco, con frecuentes heladas tardías que se corresponde al piso supramediterráneo. Como señalan Huntsinger & Bartolome (1992) la dehesa como sistema de explotación agrícola y ganadera, ha persistido desde los últimos 1000 años. La diversidad de la producción y vegetación contribuyen tanto a la estabilidad ecológica como a la estabilidad económica del propietario. El sistema de explotación de la dehesa necesita transformar el matorral en bosque abierto (Montoya Oliver, 1989). Sin embargo, es un hecho que las dehesas se están poblando de monte bajo debido, fundamentalmente, a la disminución y encarecimiento de la mano de obra para realizar labores de desbrozado, a la escasa rentabilidad en ellas de los cultivos de cereales y al aprovechamiento por una o a lo sumo por dos especies ganaderos, vacuno y lanar. (Gómez Gutiérrez et al., 1991) Para mantener espacios abiertos es necesario controlar las especies arbustivas y para ello, la roturación es la práctica más habitual. Las tierras deben ser roturadas cada 4 ó 5 años, bien como único modo de controlar el matorral o bien como parte de un periodo de producción de un cultivo. La combinación de los dos métodos puede utilizarse como control de producción de un cultivo, al tiempo que una intensa actividad ganadera puede retrasar la necesidad de la próxima roturación. Como señalan Huntsinger & W. Bartolome l.c. se pueden definir cuatro estados de vegetación en las dehesas españolas: - estado I: pastizal anual. Los procesos y las condiciones que afectan a los diferentes pasos serían considerados hipotéticos, pero la invasión del matorral es rápida, entre 2-5 años en ausencia de fuego y/ó cultivos. Los estados II y III son los mas estables y el mantenimiento de estados con alta densidad de pasto (estado I y IV) requiere comúnmente la intervención humana. En la figura 1 se dan las posiciones dinámicas de los diferentes estados de vegetación en la dehesa de Salamanca. Diversos autores vienen estudiando las variables que actúan en la invasión de especies. El objetivo de este trabajo es estudiar el paso de: Quercus/Pastizal ↔ Quercus/Matorral Crawley (1987) establece que la mayoría de los modelos de comunidades dinámicas presentan las siguientes barreras de invasión: competencia con especies nativas establecidas. pérdidas causadas por enemigos naturales. falta de mutualistas para la dispersión del polen u otras vías facilitan el invasor. efectos nocivos de baja densidad operando sobre el propio invasor. Godron (1981) señala que para poder integrar el conjunto de interacciones entre los individuos de una comunidad, se deben buscar las fuerzas macroscópicas que controlan las variaciones; la primera de estas fuerzas es el poder ascensional que produce la estratificación vertical de la vegetación; la segunda de las tendencias es el aumento general de la metastabilidad de los sistemas ecológicos que no son nunca verdaderamente estables. Allier & Lacoste (1981) señalan que el papel dinámico esencial es jugado por un número reducido de especies del matorral mediterráneo que constituyen la más importante parte del recubrimiento o de la biomasa. El mantenimiento del clímax se realiza, por tanto, por regresiones locales naturales, que permiten la reinstalación de especies arbustivas características de estados seriales jóvenes. Barbero et al. (1990) hace referencia a lugares de clima subhúmedo y húmedo del sur de España y norte de África donde las especies esclerófilas preforestales están adaptadas a la permanente perturbación antropozoogénica a que son sometidos. El objetivo, por tanto, de este trabajo es conocer el funcionamiento y los mecanismos utilizados para la invasión por las diferentes especies de matorral en la dehesa y si en el proceso de invasión existe alguna especie más agresiva que se pueda considerar como invasora. Zona de estudio La Finca "El Campillo" (29TQF463413) (Figura 2), es una dehesa típica poblada de encinas, encuadrada dentro del Sector Salmantino de la Provincia Carpetano-Ibérico-Leonesa (Rivas Martínez,1987). Su altitud es de 841 msnm y se encuentra situada en el término municipal de Sando. Geológicamente está constituida por esquistos del Cámbrico y los suelos corresponden a cambisoles dístricos. Las precipitaciones oscilan entre 460 y 572 mm y la temperatura media anual es de 11.3ºC (Caro, 1992). Corresponden estas dehesas a la serie supramediterránea de vegetación Genisto hystricis-Quercetum rotundifoliae y sus etapas seriales constituyentes son escobales referibles a Genisto hystricis-Cytisetum multiflori Rivas Martinez (in Rivas Martinez et al., 1984). En su composición florística participan Genista hystrix Lge, Cytisus multiflorus (L'Her.) Sweet y Cytisus scoparius (L.) Link, acompañadas de Quercus rotundifolia Lam. y Lavandula stoechas L. subsp. sampaioana Rozeira. Su distribución es orensano-sanabriense, trasosmontana y salmantina, ocupando suelos silíceos en el piso supramediterráneo, bajo ombroclima seco-subhúmedo-húmedo (Rivas Martinez & Belmonte, 1987) Material y métodos Una zona del Campillo se valló con el fin de evitar agresiones por parte del ganado. Dentro de esta zona se eligieron al azar 14 parcelas contiguas con matorral (cuadrados permanentes), de las que 8 corresponden a lo que denominamos frente de avance, es decir zona de contacto entre el pastizal y el matorral con una superficie de recubrimiento lineal de hasta un 50% y las 6 restantes corresponden a lo que denominamos matorral establecido con un recubrimiento lineal del matorral de más del 50% y con desaparición casi total del pasto. El tamaño de las parcelas fue de 100 m2 (10m x 10m), medida ésta que corresponde al área mínima necesaria para que la diversidad del matorral esté bien representada (Gounot, 1969; Guinochet, 1973). En cada una de ellas se marcaron transectos a lo largo de los cuales se colocaron placas numeradas cada 20 centímetros; se establecieron de esta forma 536 m de transectos y un total de 2 680 puntos de muestreo, donde se contabilizó la presencia o ausencia del matorral y su altura, con objeto de detectar los cambios en recubrimiento lineal, altura y variabilidad de las especies fruticosas siguiendo la metodología de cuadrados permanentes (Austin, 1981; Brewer et al., 1990; Mesleard et al., 1991). Centramos nuestro interés en las especies que forman el matorral y que son Genista hystrix Lge. Cytisus multiflorus (L'Hèr.) Sweet, Cytisus scoparius (L.) Link y Quercus rotundifolia. Lam. Para cada una de ellas se estudiaron los siguientes parámetros: recubrimiento lineal global del matorral, incremento del recubrimiento lineal global, velocidad media de invasión, velocidad de invasión para cada especie, altura media del matorral, incremento de la altura en el periodo estudiado y velocidad de crecimiento para cada una de las especies. La velocidad de invasión (V) en cada uno de los cuadrados permanentes y para cada especie se calculó mediante la siguiente fórmula: V = ∆Ei/D La velocidad media de invasión de una especie (V) se calcula del siguiente modo: V = ∑Vi/N El significado de los términos utilizados es el siguiente: ∆E = incremento del recubrimiento lineal de la especie en la parcela en el periodo D La actividad de los matorrales esclerófilos de los ecosistemas mediterráneos es inhibida por dos condiciones de stress: la limitación de agua en verano (periodo seco) y las bajas temperaturas del invierno (Mitrakos, 1980). Como ocurre en otras floras mediterráneas (Arroyo, 1988) las restricciones climáticas, en cuanto a temperatura, se dan antes y después de la floración y los meses durante los que ésta se desarrolla es el periodo del año donde no existen restricciones de agua; por esto, los datos necesarios para hacer los cálculos se tomaron durante los meses de Marzo a Julio, coincidiendo con el inicio del crecimiento y la parada estival. Con los datos obtenidos se realizó un tratamiento estadístico mediante un contraste de igualdad de medias para datos apareados. Se aplicaron también diferentes índices de diversidad así como la diversidad β con objeto de comparar grado de cambio en la diversidad de especies arbustivas de los diferentes cuadrados permanentes y si hay alguna variación entre las parcelas correspondientes al frente de avance y los del matorral establecido. Resultados y discusión La invasión del pasto por el matorral en la dehesa de Salamanca se realiza, fundamentalmente, por un incremento del recubrimiento lineal y altura de las especies arbustivas, teniendo prácticamente poca importancia el crecimiento demográfico de las especies en la zona estudiada, con un 14% de germinaciones (Figura 3). El estudio de los índices de diversidad así como la diversidad β (Tabla 1) de los 14 cuadrados permanentes estudiados, nos demuestra que no hay un comportamiento diferente entre el frente de avance y el matorral establecido, aunque en todos los casos Genista hystrix Lge. juega un papel invasor claro. Se exponen los porcentajes de recubrimiento lineal de cada especie, en los meses de Marzo y Julio, calculados con las mediciones efectuadas en las parcelas, cada 20 cm, a lo largo de los 2 680 puntos señalados en los transectos (Figura 3); como se observa, Genista hystrix Lge. es la especie que tiene mayor recubrimiento lineal, seguida de Cytisus multiflorus (L'Hèr.) Sweet, Quercus rotundifolia Lam. y Cytisus scoparius (L.) Link De la misma manera se calcularon las alturas medias globales para las cuatro especies que componen el matorral (Figura 5); Genista hystrix Lge. presenta la mayor altura, con un valor muy parecido - aunque algo superior - al que muestra Cytisus multiflorus (L'Hèr.) Sweet seguidos de Cytisus scoparius (L.) Link y Quercus rotundifolia Lam. La media del incremento del recubrimiento y altura en cada una de las especies consideradas se reflejan en la Tabla 2. De estos resultados deducimos que Genista hystrix Lge. y Cytisus multiflorus (L'Hèr.) Sweet son las especies fruticosas predominantes en el matorral, ya que presentan los valores más altos en porcentaje de recubrimiento y en altura media global; y podemos asegurar, con una confianza del 95%, que las cuatro especies estudiadas muestran una diferencia significativa entre las alturas presentadas durante los meses de Marzo y Julio respectivamente. En cuanto al recubrimiento lineal, la única especie que no muestra variación es Quercus rotundifolia Lam. (Figura 4). En la tabla 3 se dan, por parcelas, la especie o especies con mayor velocidad de invasión y velocidad de crecimiento. Conclusiones Desde el punto de vista de la invasión global del matorral (Tabla 3), se deduce que la velocidad media de invasión es de 1.13+0.62 cm /día; considerando que el periodo vegetativo medio es de 75 días, se puede decir que el matorral de Genisto hystricis-Cytisetum multiflori Rivas Martinez (in Rivas Marinez et al., 1984) está invadiendo el pastizal de esta dehesa, aproximadamente, 2.26 m por año. En el estudio de los modelos de fenofases vegetativa-generativa realizado para Genista hystrix Lge., Cytisus multiflorus (L'Hèr.) Sweet y Cytisus scoparius (L.) Link (Caro, 1992, 1994) se observa que Genista hystrix Lge. tiene una fenofase vegetativa más amplia que las otras dos especies. Este aspecto junto con su mayor velocidad de crecimiento y su morfología espinosa que dificulta el aprovechamiento por el ganado, hace que la espécie endemica ibérico Genista hystrix Lge., de distribución salmantina-orensano-sanabriense y trasosmontana, sea la especie invasora en la dehesa El Campillo. Agradecimientos Este estudio se ha llevado a cabo con cargo al proyecto El matorral como invasor de la dehesa salmantina finanziado por el I.N.I.A. de Salamanca. Literatura citada Allier C. & Lacoste A. 1981. Processus dynamiques de reconstitution dans la série du Quercus ilex en Conse. Vegetatio. 46: 83-91. [ Links ] Arroyo J. 1988. Fenología de la floración en especies del matorral del sur de España. Lagascalia (Extra).: 593-606. [ Links ] Austin M.P. 1981. Permanents quadrats: An interface for theory and practice. Vegetatio. 46: 1-10. [ Links ] Barbero M., Bonin G., Loisel R. & Querel P. 1990. Changes and disturbans of forest ecosystems caused by human activities in the western part of mediterranean basin. Vegetatio. 87: 151-173. [ Links ] Brewer J.S. & Grace J.B. 1990. Plant community structure in an oligohaline tidal marsh. Vegetatio. 90: 93-107. [ Links ] Caro Domínguez F. 1992. Fenomorfología, reproducción y dinamismo de la vegetación de la dehesa de "El Campillo". Memoria de Licenciatura. Facultad de Biología. Universidad de Salamanca. [ Links ] Caro Domínguez F., Sánchez Anta M.A., Gallego Martín F. & Navarro Andrés F. 1994. Phenomorphological aspects of three species in a shrubland on Genisto hystricis-Cytisetum multiflori Rivas Martínez in Rivas Martínez et al. in a dehesa of Salamanca. Boletim da Sociedade Broteriana. Vol LXVI (2ª serie).: 303-316. [ Links ] Crawley M.J. 1987. What makes a community invasible in colonization, succesion and stability?. Ed. A. J. Gray, M.J.Crawley and P.J. Edwards Blackwell scientific Publications. Oxford. [ Links ] Godron M. 1981. Preface in vegetation dynamics in grasslands, heathlands and mediterranean ligneous formations. Edited by Poissonet, F. Romane, M.A. Austin, E. van der Maarel & W. Schmidt. Dr. W. Junk Publisher. The Hague. [ Links ] Gómez Gutiérrez J.M. 1991. Coordinador científico. El libro de las dehesas salmantinas. Junta de Castilla y León. Consejería de Medio Ambiente y Ordenación del Territorio. Salamanca. [ Links ] Gounot M. 1969. Méthodes d'étude quantitative de la végétation. Masson. París. [ Links ] Guinochet M. 1973. Phytosociology. Ed. Masson. Paris. [ Links ] Mesleard F., Grillas P. & Lepart J. 1991. Plant succession in coastal wetland after abandonment of cultivation: the example of the Rhone delta. Vegetatio. 94: 35-45. [ Links ] Mitrakos K. 1980. A theory for Mediterranean plant life. Acta Ecológica. Ecol. Plant. 1 (15): 245-252. [ Links ] Montoya Oliver. J.M.1989. Encinas y encinares. Edic. Mundi Prensa. Madrid. [ Links ] Rivas Martínez S., Díaz González T.E., Fernández Prieto J.A., Loidi J. & Penas Merino A. 1984. La vegetación de la alta montaña cantábrica. Los picos de Europa. Ed. Leonesas. León. [ Links ] Rivas Martinez S. & Belmonte D. 1987. Sinopsis de la clase Cytisetea-scopario-striati. VII. Jornadas de Fitosociología, multicopia. Salamanca. [ Links ]
2 Departamento de Botánica. Facultad de Biología. C/ Licenciado Méndez Nieto, s/n. Campus Miguel de Unamuno. Universidad de Salamanca 37007. Salamanca. (España). Correo electrónico: asanta@aida.usal.es
3 Servicio Territorial de Medio Ambiente. Sección de Espacios Naturales. Junta de Castilla y León. C/ Villar y Macias, 1. 37002. Salamanca (España): Correo electrónico: tarlafma@jcyl.es
- estado II: matorral.
- estado III: dominancia de Quercus con baja densidad de matorral.
- estado IV: bosque abierto de Quercus con baja densidad de herbáceas anuales. 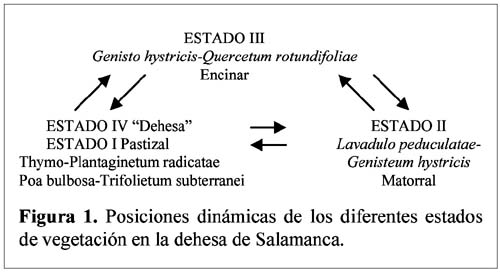
La velocidad de crecimiento (C) de cada especie en cada parcela se calcula meditante la siguiente fórmula: C =∆Hi/D
De este modo, la velocidad media de crecimiento (C) de una especie es la siguiente: C =∑Ci/N
i = parcelas 1 a 14
D = número de días transcurridos entre las dos mediciones (117)
N = número de parcelas total (14)
∆H = incremento de la altura en el periodo D