Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología Aplicada
versión impresa ISSN 1726-2216
Ecol. apl. v.3 n.1-2 Lima ene./dic. 2004
ARTÍCULO ORIGINAL
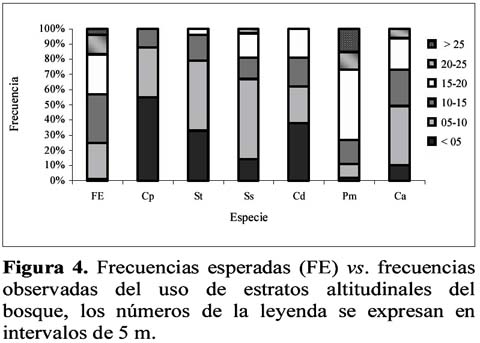
Preferencia de hábitat de seis primates simpátricos del Yasuní, Ecuador
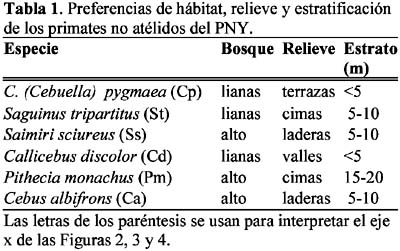
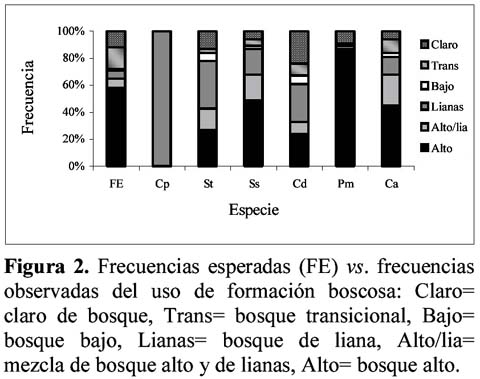
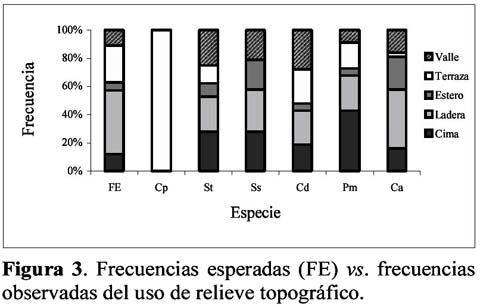
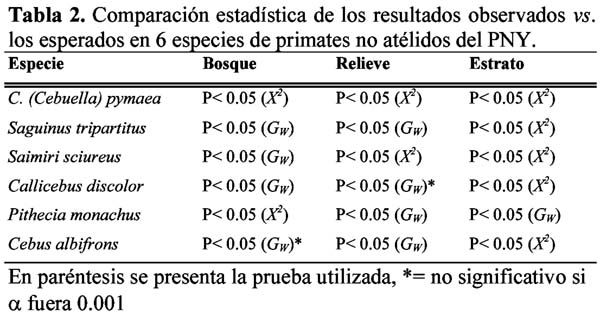
Habitat preferences of six non-atelidae primates of Yasuni, Ecuador Wilmer E. Pozo R.1 1 D.C.B. Programa de Doctorado en Ciencias Biológicas, Escuela de Biología, Universidad Central del Ecuador, Ciudadela Universitaria, Quito – Ecuador. Dirección actual: Escuela Politécnica del Ejército, Facultad de Ciencias Agropecuarias, Centro de Investigaciones del IASA, Laboratorio de Zoología. Av. El Progreso s/n, PO Box: 231-B, Sangolquí – Ecuador, E-mail: wepozor@hotmail.com Resumen Los bosques lluviosos tropicales albergan una gran cantidad de especies que viven en simpatría. El conocimiento de sus formas de uso de hábitat, permite entender cómo se reduce la competencia por espacio entre dichas especies. En este estudio se reportan datos sobre uso de hábitat de primates no atélidos del Parque Nacional Yasuní, Ecuador. Los bosques altos de las laderas y cimas y el estrato ubicado bajo el dosel son los hábitats más utilizados por la comunidad de primates estudiada, aunque lo hacen en proporciones distintas a las disponibles en el bosque y en formas diferentemente significativas entre especies. Esto permite demostrar que los primates no atélidos del Yasuní, evitan competencia mediante distintas forma de uso de hábitat. Palabras clave: monos no atélidos, nicho ecológico, competencia, Ecuador. Abstract The tropical rain forest contains great numbers of species living sympatrically. The knowledge of the uses of the habitat permits the understanding of how the competition for space is reduced between species. This study reports data on habitat use of non-atelidae primates found in the Yasuni National Park, Ecuador. The high forests on the mountain slopes and ridges and the layer below the canopy are the habitat most utilized by the primate community, but they do so in different frequencies with respect to those in the forest, and in significantly different ways from the other sympatric species. This demonstrates that the non-atelidae primates at Yasuní reduce competition through various methods of habitat use. Key words: non-atelidae primates, ecological niche, competition, Ecuador. Introducción Los bosques tropicales son los más diversos del mundo, por lo que varias especies de animales comparten sus espacios y compiten por otros recursos. Muchas especies de un área determinada no se ven afectadas por la presencia o ausencia de otras (Krebs, 1985) e incluso, las diferencias de conducta entre las especies, posibilita su coexistencia (Gausse, 1935). En ese entorno, los estudios sinecológicos permiten conocer cómo se relaciona un grupo de organismos de varias especies con su ambiente (Krebs, 1985) para poder identificar cómo las poblaciones y comunidades usan su hábitat. En 1994 se instauró, en el Ecuador, el Proyecto Primates Ecuador (PPE) con el objetivo principal de realizar estudios sistemáticos de largo tiempo con los grandes primates (Atelidae) del Parque Nacional Yasuní (PNY). A la par que se realizó estudios del comportamiento ecológico y posicional de los mencionados primates (Alouatta seniculus, Ateles belzebuth y Lagothrix poeppiggi), se decidió observar el comportamiento de la comunidad primatológica del sitio de estudio, para así obtener información sobre el uso del hábitat de los otros integrantes de la comunidad primatológica. Siguiendo Groves (2001), las especies que conforman la lista primatológica del PNY son: Callithrix (Cebuella) pymaea (Spix), Saguinus fucsicollis (Spix)1 S. tripartitus (Milne-Edwards), Saimiri sciureus (Linnaeus), Callicebus cupreus (Spix)2 , Aotus vociferans (Spix), Pithecia aequatorialis Hershkovitz, P. monachus (Geoffroy), Cebus albifrons (Humboldt), Lagothrix poeppiggi Schinz, Alouatta seniculus (Linnaeus) y Ateles belzebuth (Geoffroy). Normalmente los estudios de preferencia de hábitat, se basan en el mayor número de especies censadas (Mittermeier & van Roosmalen, 1981; Peres, 1997) o en la mayor cantidad de avistamientos de una determinada especie en ciertos sistemas ecológicos (de la Torre et al., 1995; Jiménez, 1995). Son raros los estudios que comparan la disponibilidad de características del hábitat con su uso (Thielen et al., 1997; Pozo, 2001, en prensa); además es escasa la aplicación de las pruebas de frecuencias observadas vs. esperadas para determinar el uso preferencial de hábitat (da Fonseca, 1985; Pozo, en prensa). En este artículo, se presenta el uso preferencial de tres parámetros ecológicos de los primates no atélidos que viven simpátricamente en el hábitat de terra firme del PNY, en la provincia Oriental de Orellana, Ecuador. Materiales y métodos Área de estudio Los datos fueron colectados en el PNY, entre Diciembre de 1994 y Septiembre de 1996, en un área ubicada en el km 47 de la carretera Pompeya Sur – Iro (00°4201´´ S, 76°2805 O., Figura 1), la misma que presentó una extensión aproximada de 400 ha y altitudes que oscilaron entre los 280 y 330 msnm La precipitación y la temperatura media mensual fueron de 246 mm y de 27.71°C, respectivamente (Pozo, 2001). A pesar de existir una buena producción de frutos durante todo el año, se puede definir dos picos de productividad de frutos maduros en el área de estudio (Pozo, 2003). Estructura del hábitat En tres senderos lineales de 1000 m de longitud por 10 m de ancho, se midió el DAP y la altura de 493 árboles; y, en 303 puntos de observación de los mismos senderos, se determinó el tipo de relieve (cima, ladera, terraza, valle y riachuelo) y el tipo de bosque (bosque alto, bosque alto con lianas, bosque de lianas, bosque bajo, bosque transicional y claro de bosque). Datos obtenidos Al momento del contacto con alguna especie de primate se tomó la siguiente información (First Sight Data -FSD-): hora del día, especie contactada, sendero y ubicación, tipo de bosque, tipo de relieve y altura de ubicación de la especie en el bosque. Se registró 270 FSD para primates no atélidos los que se distribuyeron así: Callithrix (Cebuella) pygmaea 18, Callicebus discolor 24, Cebus albifrons 47, Saimiri sciureus 55, Pithecia monachus 57 y Saguinus tripartitus 69. Los relieves topográficos más frecuentes fueron laderas (45%) y terrazas (26%) seguidos por cimas (12%), valles (11%) y riachuelos (6%); el tipo de bosque más disponible fue el bosque alto (58%), seguido por el bosque transicional (16%), los claros de bosque (12%), el bosque alto con lianas (7%), el bosque de lianas (6%) y el bosque bajo (1%). La altura arbórea más frecuente osciló en un rango de 10 a 15 m (32%) seguida por árboles de 15 a 20 m (26%), de 5 a 10 m (24%), de 20 a 25 m (13%), los mayores a 25 m (4%) y los menores a 5 m (1%). Análisis estadístico Los resultados del análisis de estructura de hábitat se llamarán frecuencias disponibles ya que son las características ecológicas que el sitio dispone para el uso de los animales y se utilizaron como resultados esperados en las pruebas estadísticas. Las frecuencias de los resultados esperados (también se llamarán frecuencias disponibles) y de los resultados obtenidos (provenientes de FSD) fueron dispuestas en tablas de contingencia 5 x 2 (para uso de relieve) y 6 x 2 (para uso de formación boscosa y estrato altitudinal), se utilizó la prueba G con la corrección de Williams (GW) y, de no ser aplicable, la prueba X2 considerándose significativos los valores de P iguales o menores a 0.05. Resultados Sinecología de Primates del PNY En la Tabla 1 se presenta el comportamiento sinecológico de los primates sujetos de estudio. La mencionada Tabla se realizó usando el porcentaje de observación más alto y las especies se ordenaron de acuerdo con el incremento de masa corporal de un adulto (Peres, 1997). Callithrix (Cebuella) pygmaea Se observó este diminuto primate exclusivamente en los bosques de liana (Figura 2); usó solamente las terrazas (Figura 3); sus actividades las realizó en 3 estratos altitudinales: <5 m (55%), 5 a 10 m (33%) y 10 a 15 m (18%) (Figura 4). Estos resultados demostraron diferencias significativas entre el uso y la disponibilidad de formación boscosa, relieve y estrato (Tabla 2) y además este comportamiento fue significativamente diferente al de los otros miembros de la comunidad no atélida de primates del PNY (Tabla 3). Saguinus tripartitus La especie fue registrada en los bosques de lianas (35%) y en los bosques altos (27%), fue encontrado también en los bosques altos con lianas (16%), bordeando los claros de bosque (13%) y muy rara vez en el resto de formaciones boscosas (Figura 2); usó todas las formaciones topográficas, aunque fue raro encontrarla cerca de los riachuelos (Figura 3); fue común en estratos inferiores a 20 (Figura 4). Las proporciones de uso fueron diferentemente significativas a las disponibles (Tabla 2) y este comportamiento fue tan solo similar con el uso de formación boscosa de Callicebus discolor y con el uso topográfico de Cebus albifrons (Tabla 3). Saimiri sciureus Este animal usó comúnmente los bosques altos (49%), pero también los bosques altos con lianas, los bosques de lianas (19% c/u) y, aunque en menor frecuencia, el resto de formaciones boscosas del sitio (Figura 2); las laderas (30%) y cimas (28%) fueron los relieves más frecuentados (Figura 3); la mayoría de sus actividades, las realizó en estratos inferiores a 25 m (Figura 4). Los datos obtenidos, para esta especie, fueron significativamente diferentes de los esperados (Tabla 2) y con los de los otros miembros de la comunidad, pero no fueron significativos con los de Cebus albifrons (Tabla 3). Callicebus discolor Este mono fue avistado en los bosques de lianas (28%), en los claros de bosque (24%) y en los bosques altos (24%) aunque también fue encontrado en otras formaciones boscosas disponibles en el sitio de estudio (Figura 2); fue registrado en los valles, laderas, terrazas y cimas (28%, 24%, 24% y 19%, respectivamente) pero también fue visto cerca a los riachuelos (5%) (Figura 3); los estratos altitudinales que usó fueron inferiores a 20 m (Figura 4). Estos primates usaron las formaciones boscosas, el relieve topográfico y los estratos altitudinales en forma significativamente diferentes a las disponibles (Tabla 2) y tan solo las formaciones boscosas uso en forma similar a Saguinus tripartitus (Tabla 3). Pithecia monachus Fue muy común en los bosques altos (87%) aunque frecuentó también los claros de bosque (9%), los bosques altos con lianas y los bosques transicionales (2% c/u) (Figura 2); usó las cimas, laderas y terrazas (43%, 25% y 18%, respectivamente) pero fue visto también en los valles y al borde de los riachuelos (9% y 5%, respectivamente) (Figura 3); los estratos del bosque más frecuentados oscilaron desde los 10 m hasta alturas mayores a 25 m, en cambio que los estratos inferiores a 10 m los usó con poca frecuencia (Figura 4). También presentó diferencias significativas con las características disponibles del hábitat (Tabla 2) y además diferencias significativas con el mismo tipo de comportamiento frente a los demás miembros de la comunidad de primates no atélidos, pero, en cuanto al uso de formación boscosa, sus proporciones no fueron comparables con las de C. (Cebuella) pygmaea (Tabla 3). Cebus albifrons Se encontró con mayor frecuencia en los bosques altos (45%), pero usó también los bosques altos con lianas, bosques de lianas y los bosques transicionales (23%, 13% y 10%, respectivamente), las otras formaciones boscosas del sitio fueron usadas con menor frecuencia (Figura 1); los relieves utilizados por la especie fueron las laderas (42%), riachuelos (23%), cimas (16%), valles (16%) y terrazas (3%) (Figura 2); usó todos los estratos del bosque excepto los mayores a 25 m (Figura 3). Sus frecuencias observadas presentaron diferencias significativas con las frecuencias disponibles (Tabla 2) y este tipo de comportamiento fue tan solo similar al de Saimiri sciureus (Tabla 3). Discusión En marmosas, la selección del microhábitat no está influenciada por las variables ambientales sino por el conocimiento previo que los animales tienen del sitio (Thielen et al., 1997); los primates por su parte prefieren ciertos tipos boscosos, con características ecológicas propias, donde son mayormente avistados (da Fonseca, 1985) e incluso donde prefieren dormir (Pozo, en prensa). El uso de formación boscosa fue diferente al esperado y la comparación interespecífica mostró similitudes solo entre (1) Saguinus y Callicebus y entre (2) Cebus y Saimiri, el primer caso se debe a que se registró independientemente a ambas especies pero siempre en el mismo sitio (este del área de estudio), el segundo caso, en cambio, tiene su explicación en que tropas de ambas especies casi siempre viajan juntas (obs. pers.). No se pudo comparar el uso de formación boscosa entre C. (Cebuella) y Pithecia debido a las exigencias de las pruebas estadísticas; igual que en otros sitios (Mittermeier & van Roosmalen, 1981), en el PNY, la mayoría de especies primatológicas utilizaron los bosque altos en forma preferencial. En el PNY, todos los relieves topográficos fueron utilizados pero cimas y laderas fueron más preferidos por la comunidad estudiada. En el río Duda, Colombia, los relieves topográficos mayormente usados son: tierras altas, lomas y planicies (Yoneda, 1988). Las diferencias significativas, entre uso de relieve y sus formaciones disponibles en el hábitat, demostraron ser preferenciales; además este parámetro ecológico se usó en forma diferente por la mayoría de primates, sujeto de estudio, menos entre Saguinus y Cebus y entre Saimiri y Cebus por las mismas razones anotadas en el párrafo anterior. Nótese que C. (Cebuella) fue avistado solamente en las terrazas y en los bosques altos, esto se debe a las características ecológicas del ámbito hogareño del grupo estudiado y no necesariamente reflejan que todos los bosques de lianas se distribuyen en las terrazas del sector (Obs. pers.) Se ha comprobado que las especies pequeñas de primates del PNY ocupan estratos bajos del bosque, mientras que especies de mayor tamaño ocupan estratos altos (Youlatos, 1999a). La relación entre tamaño y uso de estrato altitudinal se ha registrado también en ardillas (Youlatos, 1999a). En Surinam (Mittermeier & van Roosmalen, 1981), Perú (Terborgh & Janson, 1983) y Colombia (Yoneda, 1988), se ha visto que los estratos medios son más utilizados por los primates; lo que se corroboró en el PNY, donde se observó que los estratos menores a 10 m fueron los más frecuentados por la comunidad primatológica de no atélidos, con la excepción de Callicebus que usó mayormente los estratos superiores a 15 m, sin embargo de ello, se demostró que este uso es diferente a la disponibilidad de estratos altitudinales y entre la mayoría de las especies estudiadas, exceptuando entre Saimiri y Cebus. La distribución y uso de hábitat, en primates neotropicales, son consideradas como mecanismos para reducir competencia (Eisenberg, 1979); el comportamiento locomotor tiene relación directa con el tamaño corporal y parámetros ecológicos como estratificación, tipo de bosque y dieta (Fleagle & Mittermeier, 1980); diferencias entre comportamiento alimenticio, dieta y uso de hábitat arbóreo juegan un papel importante en la separación de nichos ecológicos (Tomblin & Cranford, 1994); las diferencias de uso de formación boscosa, topográfica y estrato con la disponibilidad de dichos parámetros en el bosque, además de sus frecuencias de uso diferentemente significativas entre especies, indican que existió preferencia de hábitat en las 6 especies simpátricas estudiadas, por lo que estos resultados soportan la hipótesis que varias especies pueden convivir en un mismo hábitat utilizando diferente parámetros ecológicos del mismo a fin de evitar competencia. Conclusiones Los primates no atélidos del PNY presentaron preferencia de hábitat. Casi todas las especies estudiadas usan en forma diferente a las formaciones boscosas, prefieren diferentes tipos topográficos y viven en distintos estratos altitudinales, lo que les permite evitar competencia, por espacio y otros recursos, entre ellas. Agradecimientos Deseo agradecer a la National Science Foundation (Grant NSF # SBR 9222526) y al Dr. J.G.H. Cant, Departamento de Anatomía de la Escuela de Medicina de la Universidad de Puerto Rico, por financiar mi participación en el Proyecto Primates Ecuador. Gracias al Dr. P.S. Rodman de la Universidad de California (Davis), por su aceptación de mi trabajo en el área de estudio. Deseo agradecer al Dr. L. Albuja V. y al Dr. R. Barriga del Departamento de Ciencias Biológicas de la Escuela Politécnica Nacional por asistirme en la obtención de los permisos de investigación otorgados por el INEFAN. Debo reconocer a la Dra. L. Arcos Terán y al personal de la Estación Científica Yasuní de la Pontificia Universidad Católica del Ecuador por el apoyo logístico brindado. Expreso mi gratitud al Departamento de Relaciones Comunitarias de la Maxus – YPF del Ecuador, a la ONHAE y a los grupos Huaoranis de la Carretera Pompeya Sur – Iro, por facilitar mi trabajo en el campo. Agradezco la camaradería y la ayuda en la toma de datos del Dr. Denis Youlatos y de mis compañeros de campo: J.G.H. Cant, P.S. Rodman, M.D. Rose, A. Di Fiore, J.L. Dew, K. Phillips y a mis asistentes Huaoraris: Oro Nenquimo, Boyo Orengo y Mipo Huira. Finalmente expreso mi agradecimiento a Rosie Reid por revisar y corregir el resumen en inglés de este artículo. Literatura citada Albuja V.L. 1991. Lista de vertebrados del Ecuador, Mamíferos. Politécnica, 16(3; Biología 3): 163-203. da Fonseca, G.A.B. 1985. Observation on the ecology of the muriqui (Brachyteles arachnoides E. Geofroy 1806): implication for its conservation. Primate Conservation. 5: 48-52. [ Links ] de la Torre S., Campos F. & de Vries T. 1995. Home range and birth seasonality of Saguinus nigricollis graellsi in Ecuadorian Amazonia. Am. J. Primatol. 37: 39-56. [ Links ] Eisenberg J.F. 1979. Habitat, economy and society: some correlations, and hypotheses for the Neotropical primates. En: I.S. Bernsteind y O.E. Smith (editores) Primates ecology and human origins: ecologycal influences on social organization: 215-262. Granland STPM Press. NY y London. [ Links ] Fleagle J.G. & Mitermeier R.A. 1980. Locomotor behavior, body size, and comparative ecology of seven Surinam monkeys. Am. J. Phys. Anthropol. 52: 301-314. [ Links ] Gausse G.F. 1935. Experimental demostration of Volterras periodic oscillation in the numbers of animal. J. Exp. Biol. 12: 44-48. [ Links ] Groves C. 2001. Primates taxonomy. Smithsonian Institution Press. Washington. [ Links ] Jiménez-P. P.J. 1995. Area de vida y uso preferencial del hábitat de Cebus albifrons (Primates: Cebidae) en Cuyabeno, Amazonía Ecuatoriana. Tesis de Licenciatura, PUCE. Depto. de Ciencias Biológicas. Quito. [ Links ] Krebs C.H. 1985. Ecología: estudio de la distribución y abundancia. Segunda edición. Harla. México. [ Links ] Mittermeier R.A. & van Roosmalen M.G.M. 1981. Preliminary observation on habitat utilization and diet in eight Surinam monkeys. Folia Primatol. 36: 1-39. [ Links ] Peres C.A. 1997. Primates community structure at twenty western Amazonian flooded and unflooded forest. J. Trop. Ecol. 13: 381-405. [ Links ] Pozo-R. W.E. 2001. Composición social y costumbres alimenticias del mono araña oriental (Ateles belzebuth belzebuth) en el Parque Nacional Yasuní, Ecuador. Disertación Doctoral. Escuela de Biología, Facultad de Filosofía, Universidad Central del Ecuador. Quito. [ Links ] ---------- 2003. Producción de frutos silvestres de un sector de la baja Amazonía Ecuatoriana. Ciencia. 6(1): 33-47. [ Links ] ---------- (en prensa). Caracterización del uso de los dormideros de Ateles belzebuth en el Parque Nacional Yasuní de la Amazonía Ecuatoriana. Ciencia. 7(2). [ Links ] Terborgh J. & Janson C.H. 1983. Ecology of primates in Southeasten Peru. National Geographic Society Research Report. 15: 665-662. [ Links ] Thielen D.R., Arends A., Segnini S. & Fariñas M.R. 1997. Selección del microhábitat por Marmosa xerophila (Marsupialia: Didelphidae) en un ecosistema semiárido del norte de Venezuela. Ecotropicos. 10(1): 21-28. [ Links ] Tomblin D.C. & Cranford J.A. 1994. Ecological niche differences between Alouatta palliata and Cebus capucinus comparing feeding modes, branch use, and diet. Primates. 35(3): 265-274. [ Links ] van Roosmalen M.G.M., van Roosmalen, T. & Mittermeier R.A. 2002. A taxonomic review of the titi monkeys, genus Callicebus Thomas, 1903, with the description of two new species, Callicebus bernhardi and Callicebus stephennashi, from Brazilian Amazonia. Neotrop. Primates. 10(Suppl.): 1-52. [ Links ] Yoneda M. 1988. Habitat utilization of six species of monkeys in Rio Duda, Colombia. Field Studies of the New World Monkeys. 1: 39-45. [ Links ] Youlatos D. 1999a. Comparative locomotion of six sympatric primates in Ecuador. Annales des Sciences Naturalles. 20 (4): 161-168. [ Links ] ---------- 1999b. Locomotor and postural behavior of Sciurus igniventris and Microsciurus flaviventer (Rodentia, Sciuridae) in eastern Ecuador. Mammalia. 63 (4): 405-416. [ Links ] ---------- & Pozo-R. W.E. 1999. Preliminary observation on the songo-songo (Dusky titi monkey, Callicebus molloch) of northeastern of Ecuador. Neotrop. Primates. 7(2): 45-46. [ Links ]