











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
En el Perú, una antigua corriente de evaluaciones del componente arbóreo de los bosques, comprendida bajo el término “inventario forestal”, desarrolladas mayormente entre las décadas 1970 - 2000, estuvo enfocada en reflejar, de manera central, los volúmenes maderables presentes en áreas boscosas, considerando sobre todo los árboles de maderas valiosas y diámetros mayores, por ejemplo, aquellos superiores a 35 o 40 cm de Diámetro a la Altura del Pecho (DAP) (Malleux, 1982; MINAG, 2012). Estos inventarios obviaron la identificación científica verificable de las especies, encarándola mayormente con una metodología de reconocimiento de árboles en el campo bajo sus nombres comunes por personal local no capacitado en taxonomía, y sin el respaldo de colecciones botánicas e identificaciones científicas comprobables (Vásquez & Gentry, 1987).
Con el paso del tiempo, se ha hecho evidente el valor significativo de la biodiversidad, así como el de la diversidad de productos forestales no maderables, y también el hecho de que éstos juegan un papel fundamental en los ciclos ecológicos de los cuales depende la supervivencia del hombre y otros organismos (Dinerstein et al., 2020). Por estas razones, las evaluaciones actuales de la vegetación arbórea incluyen cada vez más, a las especies de árboles que tienen tamaños menores, pero pueden ser importantes en la perspectiva mencionada.
En este contexto debe considerarse la extraordinaria diversidad biológica albergada en territorio peruano. Los notables niveles de diversidad de árboles existentes en los bosques del país no fueron revelados en detalle hasta estudios conducidos a partir de la década de los 80s, cuando evaluaciones desarrolladas en Parcelas Permanentes (PP), con metodologías de identificación taxonómica verificable, evidenciaron que en espacios como la llanura amazónica peruana, la diversidad arbórea (Diversidad Alfa o número de especies por unidad de área) puede alcanzar récords de hasta 300 especies por hectárea (Gentry, 1988, 1989, 1995; Ter Steege et al., 2003).
Las primeras cuantificaciones científicamente sustentadas de la cantidad total de especies de árboles existentes en el Perú evidenciaron, desde inicios de los 90s, que el país posee vastos espacios en los cuales las especies de árboles existentes son desconocidas (Honorio & Reynel, 2003). Estimaciones actuales sugieren que el 20% - 30% de la flora arbórea peruana no está descrita, catalogada ni conocida por la ciencia; particularmente, existen áreas que son auténticos vacíos de conocimiento en tal sentido (Honorio & Reynel, 2003; Joppa et al., 2011).
La necesidad actual, de evaluar la biodiversidad de los bosques, reconoce la importancia del estrato arbóreo como templete sobre el cual conjuntos de otros organismos encuentran sus nichos habitables; por ello, la cuantificación de esta diversidad es empleada actualmente como una herramienta para conocer el valor y estado de conservación de las áreas forestales (Antón & Reynel, 2004).
Los levantamientos de unidades muestrales para el estudio de áreas de bosque en el Neotrópico, y de modo particular en el Perú, establecidas con metodologías apropiadas para la identificación de las especies de flora, han ido en aumento en décadas recientes (Antón & Reynel, 2004; Marcelo & Reynel, 2014; De Rutte & Reynel, 2016). En ese contexto, es relevante la actividad del consorcio interinstitucional RAINFOR, que lidera el levantamiento de PP a nivel de los trópicos de todo el mundo, con énfasis en el Neotrópico (RAINFOR, 2000). No obstante, uno de los aspectos que no ha sido documentado para las formaciones boscosas del país es la manera como el diámetro mínimo, considerado en una evaluación, podría influir en la diversidad de especies hallada.
En un estudio conducido en un bosque húmedo tropical de tierra firme en la Guayana Francesa (Bordenave, 1998), de un total de 203 especies de plantas vasculares ≥ 2.5 cm DAP registradas, 40% de las especies tuvieron ≥ 2.5 cm de DAP; 21% tuvieron ≥ 10 cm DAP y solamente 8% tuvieron ≥ 25 cm DAP. Una investigación llevada a cabo en un bosque húmedo tropical semideciduo en Uganda, África, para un total de 32 parcelas de 0.5 ha, se tuvo un registro total de 269 especies de plantas vasculares ≥ 2 cm DAP, y la cantidad de especies ≥ 10 cm DAP disminuyó a 216 (Mwavu & Witkowski, 2015). Estos estudios muestran que cuando el diámetro limitante en la evaluación es mayor, el número de especies disminuye considerablemente.
El objetivo del presente trabajo fue mostrar la relación existente entre los diámetros mínimos considerados en una evaluación de la diversidad arbórea y la cantidad de especies que dicha evaluación podría registrar. Para ello, se analizaron los datos de áreas de bosque en dos diferentes formaciones ecológicas en la selva central peruana: los bosques húmedos montanos nublados y los bosques húmedos premontanos. La hipótesis es que cuando el diámetro limitante de la evaluación es mayor, el número de especies y familias disminuye considerablemente.
Materiales y métodos
Área de estudio; sus características ecológicas y de población humana
El ámbito de estudio comprendió bosques emplazados a cierta distancia del eje carretero que discurre desde Lima / Perú hacia el Este, en la gradiente elevacional del flanco oriental andino, con acceso desde las ciudades de Tarma, San Ramón, La Merced y Satipo. Políticamente, corresponde a las provincias de Chanchamayo y Satipo del Departamento de Junín / Perú. Este espacio, que históricamente ha constituido el paisaje forestal más prontamente colonizado desde la capital, Lima, se encuentra actualmente muy alterado por la presencia humana, sobre todo en las partes más accesibles, de pendiente suave o moderada. En las zonas con menos accesibilidad, existen remanentes de bosques más o menos extensos, incluyendo extensiones que han sido priorizadas para la conservación. Las actividades económicas más importantes para la población asentada son la producción de frutas tropicales, para las cuales los valles allí existentes constituyen una despensa frutícola primaria para la capital; la producción de café y, también, el turismo local (INEI, 1993).
Las localizaciones estudiadas corresponden a bosques húmedos montanos nublados, en el tercio alto de la cuenca del río Perené, entre 2 000 - 2 800 msnm, y progresivamente hacia el Este, bosques húmedos premontanos situados entre 600 - 2 000 msnm. De acuerdo con el Sistema de Clasificación Ecológica por Zonas de Vida de Holdridge (Holdridge, 1978; INRENA, 1995), están ubicadas principalmente en Bosque húmedo Premontano Tropical y Bosque muy húmedo Premontano Tropical. Las localizaciones de estudio tienen acceso desde las ciudades de San Ramón y La Merced, en el valle de Chanchamayo, y Satipo, en el tercio medio del río Perené. El trabajo ha estado centrado en tres estaciones de investigación: Puyu Sacha (APRODES), Fundo Génova y Fundo Santa Teresa; las dos últimas pertenecen al Instituto Regional de Desarrollo (IRD-Selva) de la UNALM (Figura 1).
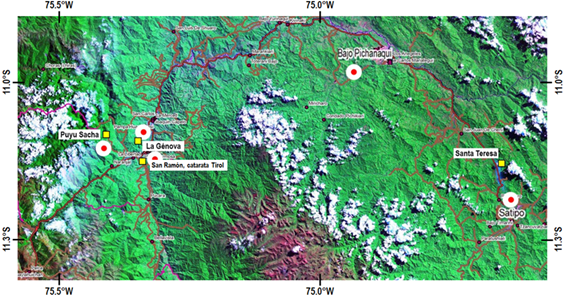
Unidades de muestra consideradas
Ocho Parcelas Permanentes (PP) de 1 ha.
Establecidas previamente (Antón & Reynel, 2004; Caro et al., 2004; Reynel & Honorio, 2004; Marcelo & Reynel, 2014; De Rutte & Reynel, 2016), éstas son parcelas mayormente de forma cuadrada, de 100 × 100 m; en pocos casos rectangulares (Tabla 1). Están subdivididas en 25 subparcelas de 20 × 20 m empleando la metodología del Consorcio de Investigación RAINFOR (Phillips et al., 2009), y son mantenidas para estudios vinculados a la dinámica de los bosques. En cada PP se han codificado y evaluado todos los árboles ≥ 10 cm de DAP. Para cada árbol, fue registrado el DAP y la altura total, y fue colectada una muestra botánica, que preparada y acondicionada de acuerdo a la metodología estándar (Bridson & Forman, 1999; Maden, 2004), fue depositada en el Herbario de la Facultad de Ciencias Forestales de la Universidad Nacional Agraria - La Molina, Lima, donde fue determinada por los dos primeros autores y botánicos especialistas que han revisado las colecciones del herbario. Se ha empleado el sistema de clasificación taxonómico por familias del Angiosperm Phylogeny Group (APG, 2016).
Ocho Transectos RAP de 0.1 ha.
Estos fueron transectos de 2 × 500 m, subdivididos en tramos de 50 m; los posicionamientos de sus puntos iniciales fueron registrados, así como su orientación geográfica; más, los transectos, no son unidades mantenidas permanentemente en el tiempo. En cada Transecto RAP fueron registradas todas las plantas ≥ 2.5 cm de DAP; cada planta fue numerada de manera sucesiva, consignando el tramo del Transecto RAP al que pertenecía (Phillips & Miller, 2002; Phillips et al., 2003). Al igual que para las PP, fue colectada para cada una de ellas una muestra botánica, que fue preparada, acondicionada, depositada y determinada en el herbario de modo similar. Todos los Transectos RAP corresponden al estrato de bosque húmedo premontano y sus características se muestran en la Tabla 2.
Tabla 2 Ubicación de los Transectos RAP de 0.1 ha establecidos en bosques húmedos del estrato premontano en la Estación Fundo La Génova UNALM en la Provincia de Chanchamayo / Departamento de Junín / Perú.
| Código Transecto | Coordenadas | Elevación (msnm) | ||
|---|---|---|---|---|
| Zona | Este | Sur | ||
| T1-30 | 18 L | 460284 | 8773083 | 1 280 |
| T2-30 | 18 L | 460367 | 8773166 | 1 240 |
| T1-40 | 18 L | 460437 | 8773458 | 1 260 |
| T2-40 | 18 L | 460465 | 8773384 | 1 230 |
| T3-40 | 18 L | 460613 | 8773484 | 1 290 |
| T1>50 | 18 L | 460823 | 8772416 | 1 116 |
| T2>50 | 18 L | 460841 | 8772586 | 1 152 |
| T3>50 | 18 L | 460881 | 8772617 | 1 136 |
Análisis efectuados
Número de especies (Diversidad Alfa)
Se ha calculado la Diversidad Alfa, en este caso expresada como el número de especies por hectárea. Se establecieron clases diamétricas cada 5 cm para analizar el número de especies (Diversidad Alfa). Se analizaron las PP por separado y, adicionalmente, el conjunto de éstas por estrato (bosque premontano y bosque montano) (Tabla 3).
Tabla 3 Clases diamétricas establecidas para el análisis de la Diversidad Alfa en las Parcelas Permanentes y Transectos RAP del presente estudio.
| Clase diamétrica | Intervalos (cm) | |
|---|---|---|
| Parcelas Permanentes | Transectos RAP | |
| 1 | 10 - 14.99 | 2.5 - 4.99 |
| 2 | 15 - 19.99 | 5 - 9.99 |
| 3 | 20 - 24.99 | 10 - 14.99 |
| 4 | 25 - 29.99 | 15 - 19.99 |
| 5 | 30 - 34.99 | 20 - 24.99 |
| 6 | 35 - 39.99 | 25 - 29.99 |
| 7 | > 40 | 30 - 34.99 |
| 8 | - | 35 - 39.99 |
| 9 | - | > 40 |
Curvas de acumulación de especies
Usando el programa EstimateS 9.1 (Colwell, 2013), se elaboraron curvas de acumulación de especies basadas en la Diversidad Alfa registrada para cada PP y Transecto RAP en cada estrato elevacional (premontano y montano). Adicionalmente, con el mismo programa se evaluó el desempeño de las curvas correspondientes al número de especies representadas por un individuo (“singletons”) y al número de especies representadas solamente en una subparcela (“uniques”). Se hizo esto para poder contrastarlas con la diversidad promediada (S Mean) y analizar cuán completo fue el inventario obtenido. Existe cierto consenso sobre que las curvas de “singletons” y “uniques” descendentes o asintóticas indicarían que el muestreo florístico realizado ha sido eficaz (Villareal et al., 2004; Cava et al., 2015).
Alpha Fisher
Se calculó el índice de Diversidad Alpha Fisher para las PP mediante el programa PAST versión 1.91 (Hammer et al., 2001). Estos cálculos se realizaron para todas las especies e individuos mayores a 10, 20, 30 y 40 cm de DAP en el estrato premontano y montano.
Resultados
Diversidad Alfa por clase diamétrica
Parcelas Permanentes - Diversidad Alfa por clase diamétrica en el estrato premontano
En las Tabla 4 se muestran los resultados de números de individuos y especies hallados en PP en bosques premontanos (elevaciones entre 600 - 1 800 msnm). En todos los casos, se trata de bosques húmedos maduros, y sin intervención humana.
Tabla 4 Número (y porcentaje) de individuos y especies por clase diamétrica en las Parcelas Permanentes evaluadas en el estrato premontano de la Selva Central del Perú.
| Clase Diam. | P-GC | P-GL | P-SRL | P-SPI | ||||
| Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | |
| 1 | 189 (37.6) | 71 (65.1) | 165 (38.7) | 49 (75.4) | 180 (39.1) | 70 (56.9) | 245 (31.9) | 94 (55.0) |
| 2 | 132 (26.2) | 62 (56.9) | 115 (27.0) | 33 (50.8) | 96 (20.9) | 54 (43.9) | 172 (22.4) | 75 (43.9) |
| 3 | 57 (11.3) | 33 (30.3) | 57 (13.4) | 25 (38.5) | 77 (16.7) | 38 (30.9) | 145 (18.9) | 69 (40.4) |
| 4 | 49 (9.7) | 31 (28.4) | 22 (5.2) | 15 (23.1) | 37 (8.0) | 21 (17.1) | 74 (9.6) | 45 (26.3) |
| 5 | 29 (5.8) | 23 (21.1) | 15 (3.5) | 12 (18.5) | 22 (4.8) | 16 (13.0) | 60 (7.8) | 36 (21.1) |
| 6 | 16 (3.2) | 12 (11.0) | 11 (2.6) | 10 (15.4) | 13 (2.8) | 7 (5.7) | 33 (4.3) | 26 (15.2) |
| 7 | 31 (6.2) | 21 (19.3) | 41 (9.6) | 29 (44.6) | 35 (7.6) | 17 (13.8) | 38 (5.0) | 24 (14.0) |
La observación de los datos de las PP en el estrato premontano muestra que, en todos los casos, la primera clase diamétrica, 10 - 14.99 cm de diámetro, registra porcentajes más altos del total de individuos (promedio = 36.8%), y del total de especies (promedio = 63.1%). Dichos porcentajes corresponden, en todos los casos, a más de la mitad y hasta más de dos tercios del total de la Diversidad Alfa hallada. La siguiente clase diamétrica, 15 - 19.99 cm, también contiene un porcentaje elevado del total de individuos (promedio = 24.1%) y del total de especies (promedio = 48.87%). En contraste, en las clases diamétricas mayores, a partir de 40 cm, se observa una disminución drástica del número de individuos y su porcentaje en relación al total (promedio = 7.1%), y de especies registradas y su porcentaje en relación al total (promedio = 22.9%) (Figura 2).
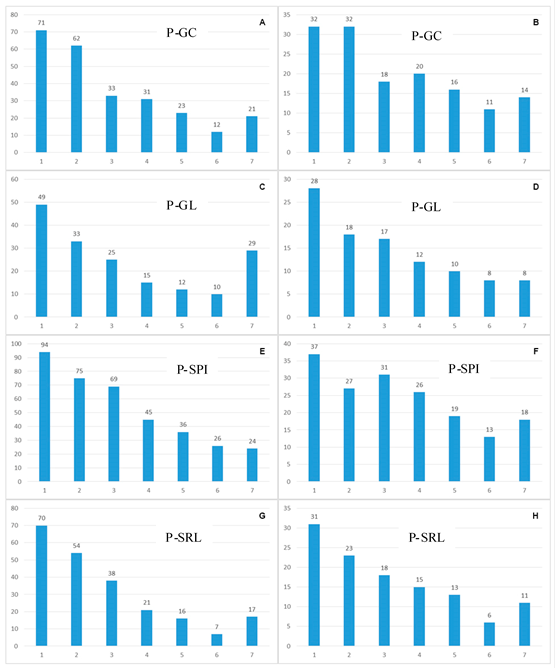
Parcelas Permanentes - Diversidad Alfa en el estrato montano
En la Tabla 5 se muestran los resultados del número de individuos y especies halladas en PP de bosques montanos (elevaciones entre 1 800 - 2 800 msnm). En todos los casos, se trata de bosques húmedos maduros, y sin intervención humana.
En cuanto al estrato montano, los datos de las PP muestran, de manera similar, que en todos los casos la primera clase diamétrica (10 - 14.99 cm de diámetro) registra porcentajes más altos del total de individuos (promedio = 38.5%), y del total de especies (promedio = 68.8%). Esos porcentajes corresponden, en todos los casos, desde más de la mitad hasta dos tercios del total de la Diversidad Alfa hallada. La siguiente clase diamétrica (15 - 19.99 cm) también contiene un porcentaje elevado del total de individuos (promedio = 21.27%), y del total de especies (promedio = 54.0%). En contraste, en las clases diamétricas mayores (a partir de 40 cm) se observa una disminución drástica del número de individuos y su porcentaje en relación al total (promedio = 7.2%), y de especies registradas y su porcentaje en relación al total (promedio = 23%) (Figura 3). Los datos reflejan, entonces, que una evaluación que incluya solo árboles de diámetros mayores a 40 cm, al igual que en los bosques premontanos, tendría una resolución muy baja del componente de Diversidad Alfa en el espacio estudiado.
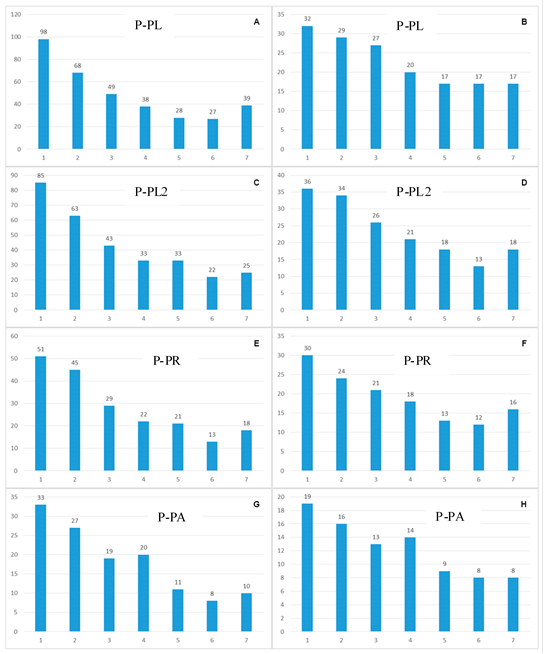
Figura 3 Número de especies y familias en función de las clases diamétricas, para las Parcelas Permanentes ubicadas en el estrato montano.
Tabla 5 Número (y porcentaje) de individuos y especies por clase diamétrica en las Parcelas Permanentes evaluadas en el estrato montano de la Selva central del Perú.
| Clase Diam | P-PL | P-PL2 | P-PR | P-PA | ||||
|---|---|---|---|---|---|---|---|---|
| Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | |
| 1 | 252 (34.3) | 98 (62.8) | 249 (35.9) | 85 (70.8) | 277 (48.2) | 51 (68.0) | 170 (35.9) | 33 (73.3) |
| 2 | 154 (21) | 68 (43.6) | 150 (21.6) | 63 (52.5) | 118 (20.5) | 45 (60.0) | 104 (22) | 27 (60.0) |
| 3 | 105 (14.3) | 49 (31.4) | 88 (12.7) | 43 (35.8) | 58 (10.1) | 29 (38.7) | 62 (13.1) | 19 (42.2) |
| 4 | 70 (9.5) | 38 (24.4) | 68 (9.8) | 33 (27.5) | 37 (6.4) | 22 (29.3) | 54 (11.4) | 20 (44.4) |
| 5 | 44 (6) | 28 (17.9) | 56 (8.1) | 33 (27.5) | 35 (6.1) | 21 (28.0) | 33 (7) | 11 (24.4) |
| 6 | 36 (4.9) | 27 (17.3) | 37 (5.3) | 22 (18.3) | 17 (3) | 13 (17.3) | 18 (3.8) | 8 (17.8) |
| 7 | 73 (9.9) | 39 (25) | 45 (6.5) | 25 (20.8) | 33 (5.7) | 18 (24) | 32 (6.8) | 10 (22.2) |
Síntesis de Parcelas Permanentes - Número de especies y familias de acuerdo al diámetro mínimo de evaluación en ambos estratos (premontano y montano)
Los datos hallados muestran que cuando las evaluaciones del componente arbóreo del bosque son realizadas con DAP mínimos de evaluación progresivamente mayores, la Diversidad Alfa registrada disminuye considerablemente. Esto se puede observar en las Tablas 6 y 7. En el estrato premontano, si se considera un DAP mínimo de 40 cm, la cantidad de especies registradas disminuye a 13.8 - 44.6% (promedio 29.2%) del total registrado para un DAP mínimo de 10 cm. Para el estrato montano, la Diversidad Alfa registrada disminuye a 20.8 - 25% (promedio 22.9%) del total registrado para un DAP mínimo de 10 cm. Asimismo, el comportamiento del índice Alpha de Fisher es coincidente con lo mencionado. Dicho índice corrige la distorsión en los valores de diversidad originada por las diferencias en el número de individuos entre los datos (Tabla 6).
Tabla 6 Número (y porcentaje) de especies y Alpha Fisher en función del diámetro mínimo de evaluación en las Parcelas Permanentes en el estrato premontano y montano de la Selva Central del Perú.
| Código de parcela | Estrato | Número total de especies (y porcentaje); Alpha Fisher | |||
|---|---|---|---|---|---|
| > 10 cm DAP | > 20 cm DAP | > 30 cm DAP | > 40 cm DAP | ||
| P-GC | Premontano | 109 (100); 42.83 | 65 (59.6); 36.17 | 45 (41.3); 46.38 | 21 (19.3); 28.6 |
| P-GL | Premontano | 65 (100); 21.37 | 49 (75.4); 25.88 | 36 (55.4); 31.69 | 29 (44.6); 44.16 |
| P-SRL | Premontano | 123 (100); 54.98 | 67 (54.5); 37.92 | 32 (26); 22.79 | 17 (13.8); 13.03 |
| P-SPI | Premontano | 171 (100); 68.29 | 116 (67.8); 60.65 | 63 (36.8); 47.7 | 24 (14); 27.97 |
| P-PL | Montano | 156 (100); 60.63 | 96 (61.5); 45.67 | 64 (41); 41.36 | 39 (25); 34.04 |
| P-PL2 | Montano | 120 (100); 41.89 | 74 (61.7); 31.81 | 48 (40); 26.11 | 25 (20.8); 23.16 |
| P-PR | Montano | 75 (100); 23.03 | 47 (62.7); 20.68 | 33 (44); 19.81 | 18 (24); 16.21 |
| P-PA | Montano | 45 (100); 12.22 | 30 (66.7); 9.81 | 19 (42.2); 7.71 | 10 (22.2); 4.99 |
Tabla 7 Número (y porcentaje) de familias en función del diámetro mínimo de evaluación en las Parcelas Permanentes en el estrato premontano y montano de la Selva Central del Perú.
| Código de Parcela | Estrato | Número total de familias APG (2016) (y porcentaje) | |||
|---|---|---|---|---|---|
| > 10 cm DAP | > 20 cm DAP | > 30 cm DAP | > 40 cm DAP | ||
| P-GC | Premontano | 41 (100) | 33 (80.5) | 28 (68.3) | 14 (34.1) |
| P-GL | Premontano | 28 (100) | 23 (82.1) | 16 (57.1) | 14 (50) |
| P-SRL | Premontano | 39 (100) | 29 (74.4) | 18 (46.2) | 11 (28.2) |
| P-SPI | Premontano | 47 (100) | 39 (83) | 29 (61.7) | 18 (38.3) |
| P-PL | Montano | 41 (100) | 36 (87.8) | 28 (68.3) | 17 (41.5) |
| P-PL2 | Montano | 48 (100) | 39 (81.3) | 27 (56.3) | 18 (37.5) |
| P-PR | Montano | 38 (100) | 29 (76.3) | 22 (57.9) | 16 (42.1) |
| P-PA | Montano | 22 (100) | 18 (81.8) | 13 (59.1) | 8 (36.4) |
Curvas de acumulación
Hemos elaborado las curvas de acumulación de especies basadas en la Diversidad Alfa registrada para cada PP en cada estrato elevacional (premontano y montano). Para el estrato premontano se pudo observar que las curvas de “singletons” y “uniques” son asintóticas sólo para las parcelas P-GC y P-GL (ambas en La Génova), lo que indicaría que en las PP restantes en ese estrato el muestreo fue sólo relativamente eficaz y podría no haber capturado el grueso de la Diversidad Alfa estimada. Por otro lado, en las PP del estrato montano las curvas de “singletons” y “uniques” sí presentaron tendencias asintóticas, indicando que las unidades de muestra levantadas habrían capturado de manera bastante completa la Diversidad Alfa allí existente (Figuras 4 y 5).
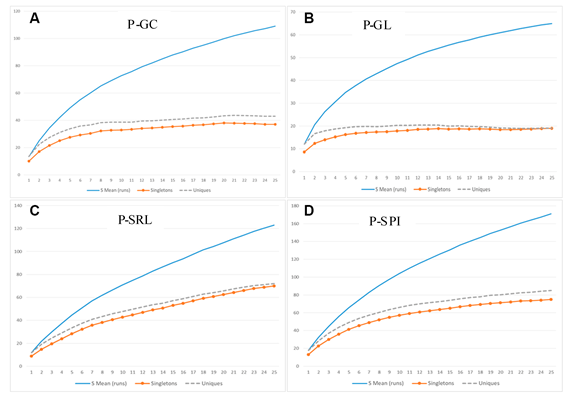
Figura 4 Curvas de acumulación de especies para las Parcelas Permanentes ubicadas en el estrato premontano.
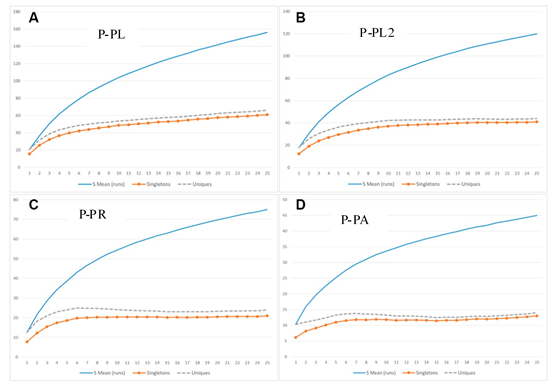
Figura 5 Curvas de acumulación de especies para las Parcelas Permanentes ubicadas en el estrato montano.
Muchos de los géneros registrados en los bosques premontanos también están presentes en los bosques montanos. Sin embargo, algunos están restringidos a estratos superiores, como los géneros Gordonia (Theaceae), Podocarpus (Podocarpaceae), Symplocos (Symplocaceae) o Weinmannia (Cunoniaceae), característicos de los bosques montanos nublados (Tablas 8 y 9).
Tabla 8 Géneros con individuos con un diámetro (DAP) mayor a 40 cm en las Parcelas Permanentes evaluadas en el estrat o premontano de la Selva Central del Perú.
| P-GC | P-GL | P-SRL | P-SPI |
|---|---|---|---|
| Alchornea, Aspidosperma, Batocarpus, Cariniana, Celtis, Clarisia, Dendropanax, Endlicheria, Huertea, Inga, Licaria, Margaritaria, Mauria, Meliosma, Myrcianthes, Nectandra, Ocotea, Pseudolmedia, Terminalia | Alchornea, Aniba, Batocarpus, Cecropia, Celtis, Chimarrhis, Chrysophyllum, Clarisia, Erythrina, Ficus, Garcinia, Hymenolobium, Licaria, Maclura, Margaritaria, Mauria, Nectandra, Otoba, Pourouma, Pseudobombax, Pseudolmedia, Trichilia, Urera, Vatairea, Virola | Alchornea, Bixa, Cecropia, Ceiba, Cosmibuena, Ficus, Ladenbergia, Macbrideina, Machaerium, Nectandra, Ochroma, Pourouma, Sciodaphyllum, Tachigali, Trichilia, Trophis, Virola | Alchornea, Cecropia, Clarisia, Dendropanax, Drypetes, Endlicheria, Eschweilera, Eugenia, Geonoma, Miconia, Myrsine, Oenocarpus, Oreopanax, Ormosia, Pleuranthodendron, Pourouma, Pouteria, Sciodaphyllum, Senefeldera, Simira, Sloanea, Trattinnickia, Virola |
Tabla 9 Géneros con individuos de un diámetro (DAP) mayor a 40 cm en las Parcelas Permanentes evaluadas en el estrato montano de la Selva Central del Perú.
| P-PL | P-PL2 | P-PR | P-PA |
|---|---|---|---|
| Alchornea, Alchorneopsis, Aniba, Beilschmiedia, Cecropia, Coussapoa, Cyathea, Elaeagia, Faramea, Ficus, Guatteria, Heliocarpus, Hieronyma, Huertea, Magnolia, Nectandra, Ocotea, Piper, Podocarpus, Protium, Prumnopitys, Pseudolmedia, Sapium, Simira, Tovomita, Weinmannia | Ampelocera, Aniba, Coussapoa, Eugenia, Ficus, Gordonia, Hieronyma, Huertea, Jacaratia, Juglans, Magnolia, Morus, Myrcia, Myrcianthes, Nectandra, Persea, Piper, Protium, Prumnopitys, Prunus, Styrax, Symplocos, Trema | Blakea, Brunellia, Cecropia, Croton, Cupania, Elaeagia, Euplassa, Ficus, Hieronyma, Inga, Magnolia, Meliosma, Nectandra, Ocotea, Ruagea, Weinmannia | Clusia, Gordonia, Miconia, Myrsine, Podocarpus, Symplocos, Ternstroemia, Weinmannia |
Transectos RAP
El conjunto de los ocho Transectos RAP analizados corresponden al estrato premontano. Los resultados del análisis de dichos transectos se muestran en las Tablas 10 y 11. La observación de los datos de los Transectos RAP correspondientes en el estrato premontano (Tabla 10) muestra que, en todos los casos, la primera clase diamétrica (2.5 - 4.99 cm) registra los porcentajes más altos del total de individuos (promedio = 44.6 %) y del total de especies (promedio = 63.1%). Dichos porcentajes corresponden, en todos los casos, a más de la mitad y hasta más de dos tercios del total de la Diversidad Alfa hallada. La siguiente clase diamétrica (15 - 19.99 cm) también contiene un porcentaje elevado del total de individuos (promedio = 28.1%) y del total de especies (promedio = 59.7%). En contraste, en las clases diamétricas mayores (a partir de 40 cm) se observa una disminución drástica del número de individuos y su porcentaje, en relación al total (promedio = 1.8%), y de especies registradas y su porcentaje en relación al total (promedio = 8.4%) (Figura 6). Empleando este tipo de Transectos, al igual que con PP como unidades de muestra, una evaluación inclusiva solamente de diámetros mayores a 40 cm tendría una resolución muy baja del componente de Diversidad Alfa arbórea en el bosque estudiado. Adicionalmente, en la Tabla 11 se pueden apreciar los géneros con individuos con un diámetro (DAP) mayor a 40 cm.
Tabla 10 Número (y porcentaje) de individuos y especies en Transectos RAP evaluados en el estrato premontano de la Selva Central del Perú.
| Clase Diam | T1-30 | T2-30 | T1-40 | T2-40 | T3-40 | T1>50 | T2>50 | T3>50 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | Núm. inds. (%) | Núm. spp. (%) | |
| 1 | 222 (36.9) | 35 (62.5) | 253 (42.2) | 38 (69.1) | 346 (50.7) | 58 (72.5) | 229 (39.6) | 42 (66.7) | 330 (51.9) | 45 (80.4) | 339 (47.8) | 46 (73) | 282 (41.2) | 45 (60) | 285 (46.8) | 46 (75.4) |
| 2 | 228 (37.9) | 41 (73.2) | 186 (31) | 33 (60) | 195 (28.6) | 48 (60) | 160 (27.7) | 36 (57.1) | 163 (25.6) | 37 (66.1) | 176 (24.8) | 35 (55.6) | 187 (27.3) | 34 (45.3) | 134 (22) | 37 (60.7) |
| 3 | 62 (10.3) | 24 (42.9) | 67 (11.2) | 26 (47.3) | 55 (8.1) | 22 (27.5) | 61 (10.6) | 24 (38.1) | 66 (10.4) | 21 (37.5) | 68 (9.6) | 23 (36.5) | 96 (14) | 17 (22.7) | 88 (14.4) | 24 (39.3) |
| 4 | 34 (5.7) | 16 (28.6) | 29 (4.8) | 15 (27.3) | 28 (4.1) | 18 (22.5) | 43 (7.4) | 16 (25.4) | 23 (3.6) | 14 (25) | 57 (8) | 23 (36.5) | 47 (6.9) | 20 (26.7) | 47 (7.7) | 26 (42.6) |
| 5 | 20 (3.3) | 11 (19.6) | 24 (4) | 16 (29.1) | 20 (2.9) | 16 (20) | 29 (5) | 16 (25.4) | 25 (3.9) | 8 (14.3) | 22 (3.1) | 15 (23.8) | 17 (2.5) | 11 (14.7) | 22 (3.6) | 10 (16.4) |
| 6 | 13 (2.2) | 9 (16.1) | 17 (2.8) | 8 (14.5) | 13 (1.9) | 9 (11.3) | 19 (3.3) | 9 (14.3) | 19 (3) | 5 (8.9) | 20 (2.8) | 14 (22.2) | 15 (2.2) | 12 (16) | 12 (2) | 10 (16.4) |
| 7 | 11 (1.8) | 9 (16.1) | 9 (1.5) | 4 (7.3) | 8 (1.2) | 6 (7.5) | 11 (1.9) | 6 (9.5) | 6 (0.9) | 2 (3.6) | 14 (2) | 10 (15.9) | 11 (1.6) | 8 (10.7) | 8 (1.3) | 7 (11.5) |
| 8 | 1 (0.2) | 1 (1.8) | 2 (0.3) | 2 (3.6) | 4 (0.6) | 3 (3.8) | 7 (1.2) | 6 (9.5) | 2 (0.3) | 1 (1.8) | 8 (1.1) | 7 (11.1) | 7 (1) | 5 (6.7) | 5 (0.8) | 5 (8.2) |
| 9 | 10 (1.7) | 7 (12.5) | 13 (2.2) | 5 (9.1) | 14 (2) | 7 (8.8) | 19 (3.3) | 9 (14.3) | 2 (0.3) | 2 (3.6) | 5 (0.7) | 4 (6.3) | 22 (3.2) | 6 (8) | 8 (1.3) | 3 (4.9) |
Tabla 11 Géneros con individuos de un diámetro (DAP) mayor a 40 cm en los Transectos RAP evaluados en el estrato premontano de la Selva Central del Perú.
| Transectos RAP | Géneros | Transectos RAP | Géneros |
|---|---|---|---|
| T1-30 | Cecropia, Erythrina, Heliocarpus, Juglans, Maclura, Ocotea, Piptadenia | T3-40 | Hieronyma, Sapium |
| T2-30 | Cecropia, Erythrina, Guarea, Maclura, Schizolobium | T1>50 | Ceiba, Erythrina, Inga, Maclura |
| T1-40 | Erythrina, Ficus, Inga, Juglans, Ocotea, Schizolobium | T2>50 | Cecropia, Ficus, Neea, Ochroma, Sapium |
| T2-40 | Acacia, Cecropia, Erythrina, Inga, Juglans, Maclura, Ocotea, Piptadenia, Pleuranthodendron | T3>50 | Cecropia, Ceiba, Ficus |
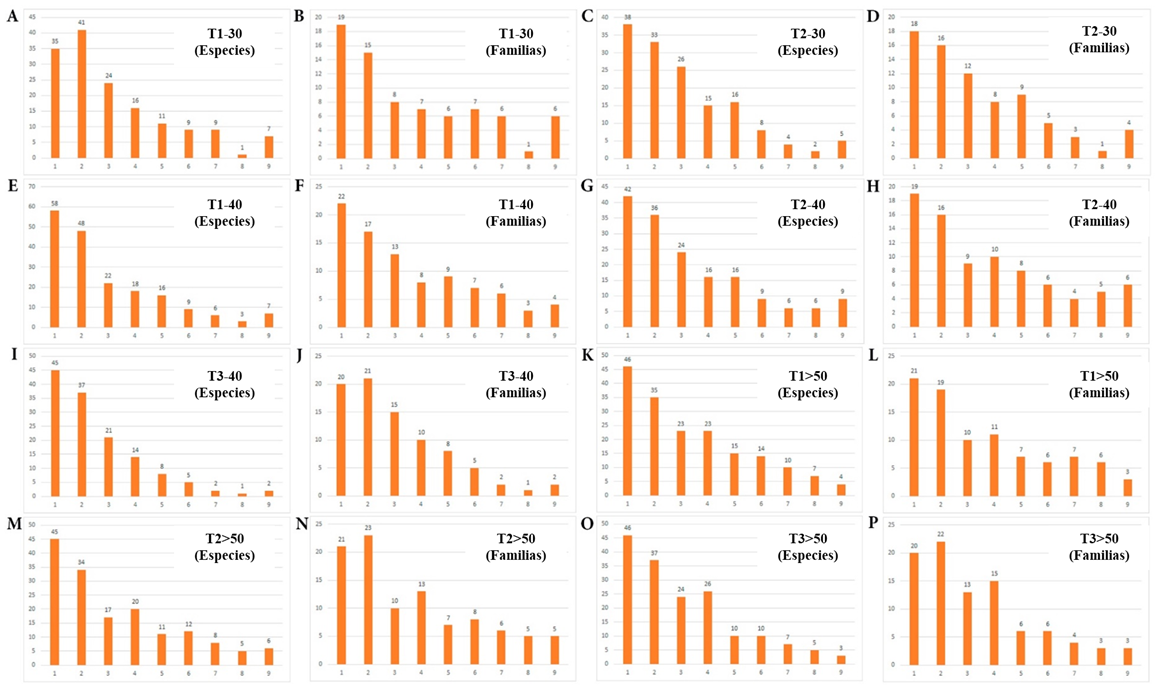
Figura 6. Número de especies y familias en función de las clases diamétricas, para los Transectos RAP ubicados en el estrato premontano.
Tabla 12 Número (y porcentaje) de especies en función del diámetro mínimo de evaluación en los Transectos RAP evaluados (estrato premontano) de la Selva Central del Perú.
| Código de Transecto | Número total de especies (Porcentaje) | ||||
|---|---|---|---|---|---|
| > 2.5 cm DAP | > 10 cm DAP | > 20 cm DAP | > 30 cm DAP | > 40 cm DAP | |
| T1-30 | 56 (100) | 33 (58.9) | 18 (32.1) | 12 (21.4) | 7 (12.5) |
| T2-30 | 55 (100) | 35 (63.6) | 20 (36.4) | 7 (12.7) | 5 (9.1) |
| T1-40 | 80 (100) | 37 (46.3) | 25 (31.3) | 13 (16.3) | 7 (8.8) |
| T2-40 | 63 (100) | 37 (58.7) | 23 (36.5) | 16 (25.4) | 9 (14.3) |
| T3-40 | 56 (100) | 33 (58.9) | 12 (21.4) | 3 (5.4) | 2 (3.6) |
| T1>50 | 63 (100) | 35 (55.6) | 22 (34.9) | 15 (23.8) | 4 (6.3) |
| T2>50 | 75 (100) | 43 (57.3) | 24 (32) | 14 (18.7) | 6 (8) |
| T3>50 | 61 (100) | 37 (60.7) | 21 (34.4) | 13 (21.3) | 3 (4.9) |
Tabla 13 Número (y porcentaje) de familias en función del diámetro mínimo de evaluación en los Transectos RAP evaluados (estrato premontano) de la Selva Central del Perú.
| Código de Transecto | Número total de familias APG (Porcentaje) | ||||
|---|---|---|---|---|---|
| > 2.5 cm DAP | > 10 cm DAP | > 20 cm DAP | > 30 cm DAP | > 40 cm DAP | |
| T1-30 | 21 (100) | 12 (57.1) | 9 (42.9) | 8 (38.1) | 6 (28.6) |
| T2-30 | 21 (100) | 13 (61.9) | 9 (42.9) | 4 (19) | 4 (19) |
| T1-40 | 23 (100) | 15 (65.2) | 12 (52.2) | 9 (39.1) | 4 (17.4) |
| T2-40 | 22 (100) | 13 (59.1) | 10 (45.5) | 8 (36.4) | 6 (27.3) |
| T3-40 | 26 (100) | 20 (76.9) | 11 (42.3) | 3 (11.5) | 2 (7.7) |
| T1>50 | 25 (100) | 14 (56) | 9 (36) | 9 (36) | 3 (12) |
| T2>50 | 31 (100) | 22 (71) | 13 (41.9) | 8 (25.8) | 5 (16.1) |
| T3>50 | 27 (100) | 19 (70.4) | 10 (37) | 7 (25.9) | 3 (11.1) |
Síntesis de Transectos RAP - Número de especies y familias de acuerdo al diámetro mínimo de evaluación en ambos estratos
Los datos hallados muestran que cuando las evaluaciones del componente arbóreo del bosque son realizadas con DAP mínimos de evaluación progresivamente mayores, la Diversidad Alfa registrada disminuye considerablemente. Esto se puede observar en las Tablas 12 y 13. En el estrato premontano, si se considera un DAP mínimo de 40 cm, la cantidad de especies registradas disminuye a 3.6-14.3% (promedio 8.4%) del total registrado para un DAP mínimo de 2.5 cm, mientras que, en el caso de las familias, según el sistema de clasificación APG, la cantidad disminuye a 7.7 - 28.6 (promedio 17.4%).
Discusión
Es de esperar que el registro de una mayor amplitud de diámetros se traduzca en la presencia de más especies cuando se evalúa la diversidad arbórea en bosques tropicales. Esto se ha evidenciado no sólo en diversos tipos de bosques tropicales. Un estudio realizado en un bosque húmedo semideciduo en Uganda ha mostrado también una cantidad significativamente mayor de especies en diámetros menores (Mwavu & Witkowski, 2015). En un bosque húmedo de tierra firme, en la Guayana Francesa, se halló bastante menor diversidad de especies en los diámetros por encima de 25 cm de DAP (19.5%) (Bordenave et al., 1998). Los datos analizados reafirman que evaluaciones forestales que se restringen solamente a diámetros mayores, por ejemplo (aquellos ≥ 40 cm de DAP) capturan un porcentaje muy limitado de la diversidad arbórea presente en el bosque (en promedio 23%, tanto para el estrato de bosque premontano como para el estrato montano). Incluso en un estudio comparativo de mayor escala, entre bosques tropicales de Asia, África y América, se encontró que el número de especies de árboles, con diámetros superiores a 30 cm de DAP, representaba entre el 24 y 32% de la riqueza (LaFrankie et al., 2006).
Los resultados muestran que la abundancia de individuos se reduce drásticamente en las clases de diámetro superiores. Del total de árboles registrados (individuos por hectárea), aquellos con diámetros iguales o mayores a 40 cm de DAP representaron entre el 4.9 y 9.6% para los bosques premontanos y entre el 5.7 y 10% para bosques montanos. La abundancia en árboles de menor diámetro ha sido reportada en otros estudios del Neotrópico. La notable diferencia entre el número de individuos de las primeras clases diamétricas y las últimas también ha sido destacada en un estudio comparativo entre dos bosques tropicales de Ecuador y Perú. Allí se registró poco más del 40% de individuos con diámetros de entre 10 y 15 cm, mientras que menos del 3% alcanzaron diámetros iguales o mayores a 40 cm de DAP (Pitman et al., 2002). A una mayor escala, cuando se comparó el número de individuos por hectárea en seis bosques tropicales de América, Asia y África, se evidenció que en todos los casos hubo una disminución importante conforme se restringía la evaluación a clases de diámetro mayores; de este modo, considerando árboles con DAP igual o mayor a 30 cm, sólo se registraron entre el 12 y 19% respecto del total registrado a partir de 10 cm (LaFrankie et al., 2006).
Los valores hallados sugieren que una evaluación de solo árboles de diámetros mayores a 40 cm, tendría una resolución muy baja del componente de Diversidad Alfa arbórea en el espacio estudiado. La Diversidad Alfa, medida con el índice Alfa de Fisher (por ha) evidencia una importante disminución en las clases diamétricas mayores (en promedio se reduce de 46.9 a 28.4 para los bosques premontanos y de 34.4 a 19.6 para los bosques montanos). Esta tendencia a la diminución de la captura de la diversidad también se ha registrado en los bosques premontanos evaluados a partir de unidades de muestreo que incluyen diámetros a partir de 2.5 cm (Transectos RAP), en donde hay una clara reducción tanto de la abundancia como de la riqueza de especies. Un estudio, que incluyó todos los árboles con diámetros ≥ 1 cm de DAP, mostró que el valor de índice fue casi el mismo (39.4/ha) que cuando solo se consideraron los árboles con un diámetro < 10 cm de DAP (40.1); mientras que, para árboles de diámetro ≥ 10 cm de DAP el valor del índice fue mucho menor (30.8/ha) (Memiaghe et al., 2016). Si bien evaluaciones que incluyan diámetros por debajo de los 10 cm y más de 2.5 cm de DAP requieren una mayor inversión de recursos, es importante valorar la pérdida de información que podría estar ocurriendo.
A nivel regional, las familias Melastomataceae, Piperaceae y Malvaceae fueron registradas principalmente en las clases de diamétricas inferiores en bosques tropicales americanos (LaFrankie et al., 2006). Adicionalmente, recientes estudios sobre la flora leñosa (a partir de 2.5 cm de DAP), en nuestro mismo ámbito de estudio, muestran que Piperaceae es un componente principal de bosques jóvenes (en las clases diamétricas menores) y que Moraceae es un componente principal de bosques maduros (en las clases diamétricas mayores) (Echía & Reynel, 2019; Quintero et al., 2020). Puede haber importantes diferencias a nivel local, más aun considerando el ensamblaje de especies en ecosistemas montañosos como el estudiado. Los cambios en la composición florística, conforme nos acercamos a las clases diamétricas superiores, podrían ser sugerentes si se analizan desde una perspectiva funcional. Una estructura de alto número de individuos en las clases diamétricas menores ayuda a garantizar el establecimiento futuro de los árboles que ocuparán el dosel del bosque. Además, se ha sugerido que los bosques tropicales, a pesar de presentar similar número de individuos, áreas basal y taxonomía de árboles, ocupan la estructura del dosel de forma diferente (LaFrankie et al., 2006). Por otro lado, la reducción del número de individuos registrados en las evaluaciones reduce las posibilidades de incorporar especies raras o representadas por un solo individuo.
Dado que, los altos niveles de diversidad dentro de las clases de menor diámetro pueden dar a estos bosques altos niveles de resistencia estructural a las perturbaciones antropogénicas / naturales y a un clima cambiante (Memiaghe et al., 2016), es importante considerar el análisis de la diversidad de especies en rangos de diámetros más amplios, como los mostrados en este estudio. Considerando que los bajos niveles de riqueza de especies de árboles de diámetros mayores (> 60 cm de DAP) ha sido asociada a especies comúnmente conformadas por pocos individuos (Lutz et al., 2018), la estructura de las clases diamétricas menores requiere ser incluido en el análisis de la regeneración del dosel superior.
Finalmente, los árboles de diámetros mayores (> 60 cm de DAP) contribuyen con aproximadamente el 41% de la biomasa forestal, por lo que es importante considerar su dinámica y sensibilidad al cambio ambiental (Lutz et al., 2018), y para ello es necesario incluir en las evaluaciones las clases de diámetro menores, donde se registraría la regeneración natural de esas especies importantes. No obstante, son muy escasas las publicaciones que documentan este importante aspecto. Más aún, los esfuerzos por monitorear los bosques considerando individuos con diámetros menores a 10 cm son considerablemente menores, no sólo en el Neotrópico (Memiaghe et al., 2016), lo que limita la comprensión de la estructura, dinámica y diversidad de árboles de los bosques tropicales, considerando la regeneración de los árboles que ocupan el dosel superior y los árboles que nunca alcanzan los 10 cm de DAP.
Conclusiones
Las evaluaciones de la diversidad arbórea considerando solamente diámetros a la altura del pecho (DAP) ≥ 40 cm están bastante lejos de capturar el contenido de dicha diversidad en las áreas de bosque húmedo premontano y montano en la Selva Central del Perú. Considerando solamente diámetros a la altura del pecho ≥ 40 cm, la diversidad arbórea promedio hallada equivale al 29.2% del a diversidad total en el caso de bosques húmedos premontanos (940 - 1 150 msnm), y de 22.9% para el caso de bosques húmedos montanos (2 078 - 2 770 msnm).

