Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.1 n.1-2 Lima oct. 1942
TRABAJOS ORIGINALES
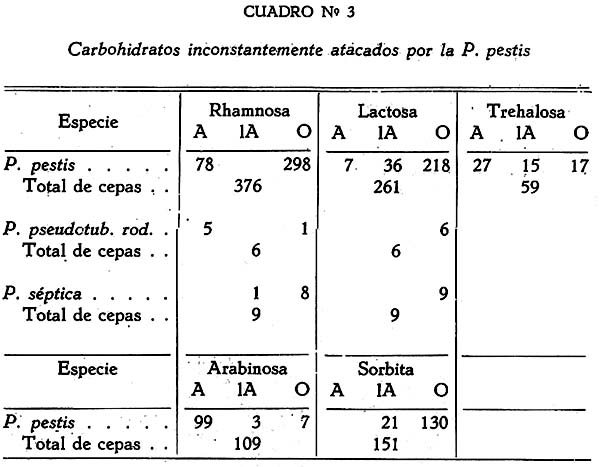
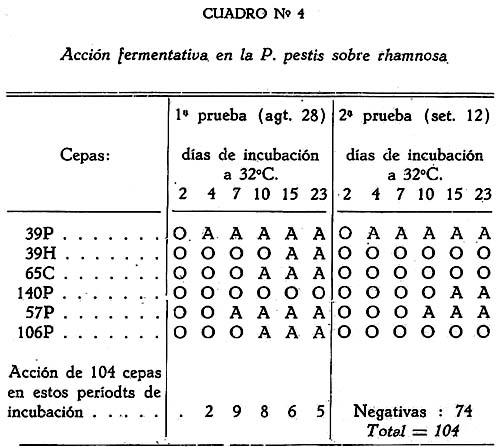
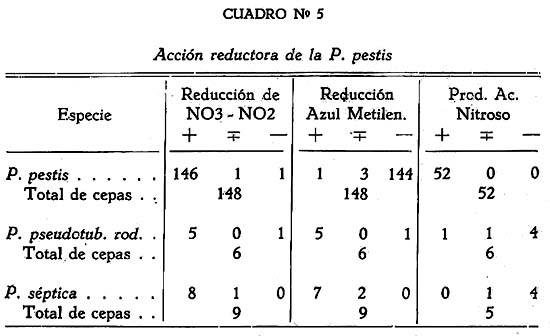
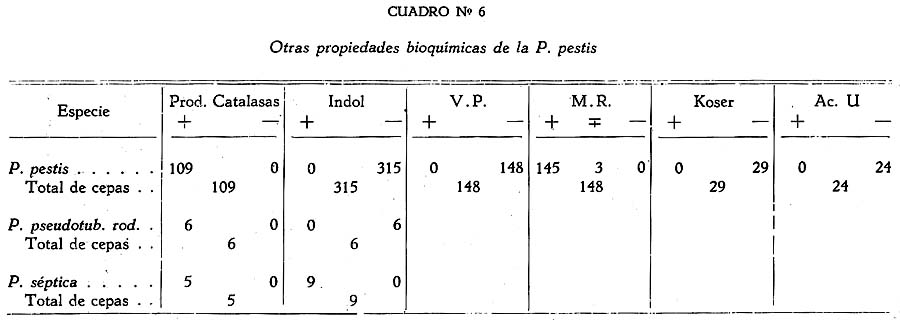
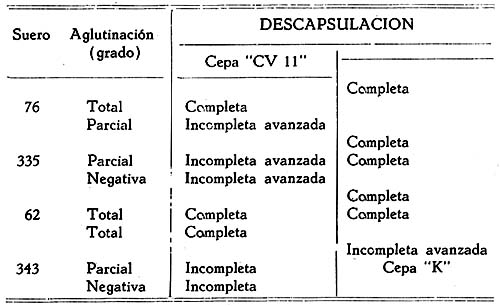
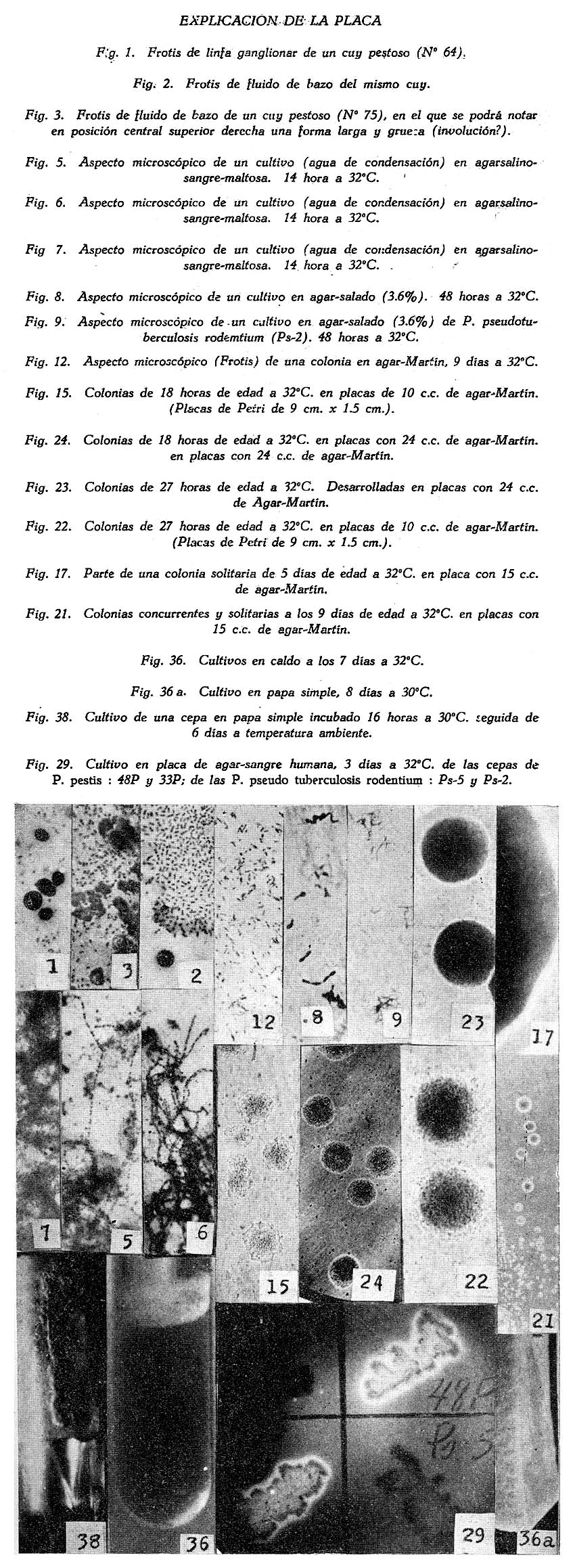
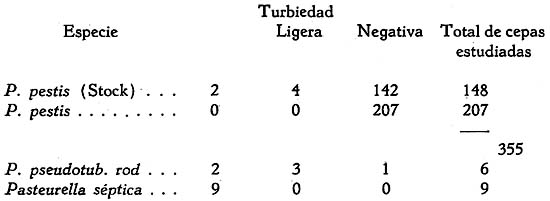
Estudio bacteriológico de la Pasteurella pestis Héctor Colichón 1 1 Departamento de Inmunología y Bacteriología, Sección Peste. SUMARIO Un considerable número de cepas de P. pestis aisladas en el Perú han sido estudiadas bacteriológicamente y los resultados obtenidos pueden ser resumidos en la forma siguiente: 2 cepas entre 148 estudiadas en caldo produjeron franco enturbiamiento del medio. La liquefación de la gelatina, la producción de indol y acetil metilcarbinol fueron negativos para todas las cepas estudiadas. En las condiciones ordinarias puede haber débil producción de SH2 por muy pocas cepas; en cambio, en el agar-Martín-infusión-extracto hepático la producción de SH2 ocurre en el mayor número de cepas, y en un considerable número de ellas en forma notablemente intensa. La temperatura de incubación es decisiva en la producción de hidrógeno sulfurado. Salvo una excepción la reducción del azul de metileno fue negativa para las cepas estudiadas, en esta misma forma la reducción de nitratos a nitritos, la producción de ácido nitroso y reacción del rojo de metilo fueron positivas para las cepas de P. pestis probadas. Todas las cepas estudiadas dieron reacción de catalasas positiva y no desarrollaron en ácido úrico, ni en medio de Koser. La acción de la P. pestis sobre los carbohidratos fue dividida en los siguientes grupos: Carbohidratos atacados constantemente con producción de ácido, no gas; ellos son: glucosa, maltosa, manita, xilosa, salicina y leche tornasolada. Carbohidratos constantemente no atacados, como son: sacarosa, rafinosa, dulcita, inosita, glicerina y dextrina. Carbohidratos, inconstantemente atacados y son: rhamnosa, trehalosa, lactosa, sorbita y arabinosa. En condiciones adecuadas de cultivo, muchas cepas pueden desarrollar en papa, entre las que desarrollaron se observaron algunas que producen pigmentación amarillo-cepia o amarillo dorado. Las pruebas, del ácido nitroso, de la reducción del azul de metileno y de la rahmnosa tienen valor relativo para el Diagnóstico de la Peste, en cambio la glicerina, el indol, la sacarosa, salicina, el agar-sangre-humana y la leche tornasolada son pruebas de valor en la diferenciación de la P. pestis, P. pseudotuberculosis y la P. séptica. I. INTRODUCCIÓN El agente etiológico de la Peste en su moderna posición sistemática figura como miembro del Género Pasteurella de la Familia Parvobacteriacea (1), constituyendo la especie Pasteure11a pestis. Como este organismo se encuentra estrechamente relacionado con los miembros del Grupo Septicemia Hemorrágica, ha permitido que algunas autoridades acepten su situación en dicho Género (2). Además, él está íntimamente vinculado y hasta participa de parentesco antigénico (3) (4) con el Baci1o pseudotub ercu1osis rodentium, organismo que según algunas clasificaciones (5) pertenece al Género Corynebacterium. Actualmente se le incluye también en el Género Pasteurella, al lado de la P. pestis y las otras especies de dicho Género. En el presente trabajo nos proponemos hacer un estudio bacteriológico de las cepas de P. pestis aisladas en el Perú. Para llevarlo a cabo hemos aprovechado de materiales y observaciones acumuladas entre los años de 1931 a 1940. Un conjunto de 265 cepas peruanas y dos ecuatorianas nos sirven para su realización. Las cepas fueron aisladas de enfermos y cadáveres a quienes se hizo la punción ganglionar o la autopsia, de cuyes autopsiados que murieron a consecuencias de la inoculación de productos pestosos de origen humano o murino, etc. En algunos casos de ratas espontáneamente infectados. El trasporte del material con el organismo viable para el cultivo o la inoculación, desde los distintos puntos del País, fue asegurado por el método de inclusión de fragmentos de órganos frescos (ganglio, bazo) en parafina sólida. (6). Estudiamos también 6 cepas de P. pseudotuberculosis rodentium y 4 Pasteurellas provenientes de The National Collection of Tipe Cultures (Medical Research Council-London), y 5 cepas de Pasteurellas aisladas de roedores en Lima. Mientras las cepas no fueron estudiadas permanecieron en stock a 10ºC en agar picadura, resembrándolas cada 4 meses. II. MATERIALES De las 265 cepas peruanas, 148 fueron aisladas y estudiadas hasta 1938 (7) y las 117 restantes en los años 1939 y 1940. Todas ellas son registradas también en la Dirección General de Salubridad, Servicio Nacional Antipestoso; proceden de Lima y alrededores, del Callao y Terminales Marítimos y de las distintas Provincias del Perú donde existen focos pestosos en actividad. Las 5 Pasteurellas fueron aisladas y estudiadas antes de 1938. Las dos cepas ecuatorianas llegaron a Lima en julio y octubre de 1936 respectivamente en fragmentos de bazo de cuy incluidos en parafina sólida. En resumen el material puede ser clasificado en la siguiente forma: CEPAS PERUANAS Pasteurella pestis: De origen humano 215 Pasteurellas aisladas de cuyes: 5 CEPAS ECUATORIANAS Pasteurella pestis: De origen humano 1 Cepas de The National Collection of Type Cultures-London III. MÉTODOS Los cultivos corrientes fueron realizados en medios obtenidos con peptona-Martín (8), ajustando su preparación a 250 gms. de estómagos de cerdo molidos para 1000 c.c. de agua acidulada al 1%; siendo debidamente controlada con la reacción del biurét al terminar la digestión. Algunos investigadores rusos han usado también medios corrientes con esta peptona para el estudio de la P. pestis. Caldo Martín: Composición: Peptona-Martín 500 c.c. Procedimiento: Calentar la peptona en baño-maría a ebullición 10 minutos, agregar el extracto de carne y, luego el cloruro de sodio, neutralizando enseguida con soda fuertemente concentrada mediante el empleo de papel-tornasol. Después de neutralizar en caliente, vuelve al b. m. 20 minutos más. Luego, dejar enfriar, ajustar el Ph. a 6.8-7.0 y filtrar por papel; terminando entonces la precipitación a 1 atm. 15 minutos. Enfriar, filtrar por papel y repartir. El Ph. del medio debe ser entonces de 6.7 a 6.9, después de la esterilización a 1 atm. 15 minutos. Debe evitarse todo calentamiento adicional a los estrictamente indicados. Agar y Gelatina (Martín). En el tiempo 2.) de la preparación del caldo, después de la primera filtración, agregar el agar o la gelatina, el primero en la proporción de 2 grms. p. 100 c.c. de caldo y el segundo en la de 14-16 gms. P. 100. 1 Después de agregado el agar se le deja remojar 1 hora, enseguida se disuelve a 1 atm. 15 minutos, filtrándosele en caliente por algodón, se reparte y esteriliza a 1 atm. 15 minutos. El Ph. al término de la preparación debe ser de 6.7 a 6.9. La gelatina se disuelve calentándola a 80:C. después de añadida al caldo, la temperatura del b.m. desciende entonces a 50-45ºC. para neutralizarla con soda y papel-tornasol. Se añade 1 clara de huevo batida para cada 1000 c.c. de medio, se precipita entonces a ½ atm. 15 minutos, filtrándosele en caliente. Después de ajustar a Ph. 6.8-7.0 se reparte y esteriliza por vapor al calor discontinuo. Agar-Martín-Infusión-Extracto hepático: Composición: Agar Martín-Infusión 1000 c.c. Agregar estérilmente el extracto hepático al agar fundido y a 50ºC. Repartir. Agar Martín-Infusión: En la composición del caldo-Martín descrito mas arriba, reemplazar el agua y el extracto de carne por 500 c.c. de infusión doble de carne; que se prepara infundiendo 1 noche en la nevera 1000 grms. de pulpa de carne fresca y 1000 c.c. de agua destilada. Calentar progresivamente en b. m. hasta ebullición y mantener a esta temperatura 30 minutos. En todo lo demás se continúa como para la preparación del caldo y agar. El Ph. no debe pasar de 6.7 a 6.9 al final del proceso. Extracto hepático: 500 a 600 grms. de hígado fresco (de buey) y molido son mezclados con 1000 c.c. de solución de cloruro de sodio (q. p.) al 0.9 p. 100. Después de permanecer una noche en la nevera se ajusta el Ph. a 7.6 mediante la soda concentrada, luego se calienta agitando suavemente a 55-56ºC. en b.m. durante 2 horas. Enseguida se filtra por algodón-gaza o lienzo, luego por papel y por último por una apretada capa de raspaduras de papel dispuesta en embudo y aparato para filtración al vacío. El líquido es entonces esterilizado por filtración a través de bujía V de Berkefeld. Después de controlar su esterilidad se reparte y guarda en la nevera hasta el uso. Agar-Sangre humana. Merece especial interés porque permite la demostración de un fenómeno característico de la P. pestis. Composición: Extracto hepático 5 c.c. En el momento de agregar la sangre el agar fundido es mantenido a 56ºC. Mezclar y continuar el calentamiento a esta temperatura 10 minutos en b.m., agitando suavemente. Repartir estérilmente placas-Petri (10 c. c. por placa de 9 x 1.5), dejar endurecer y secar. Pueden quedar en la nevera, previo control de esterilidad, hasta por 10 días. Agar-Salino-Sangre y Agar-Salino-Sangre-Maltosa. Tienen importancia en la conservación de la virulencia y para la demostración de ciertas peculiaridades morfológicas del bacilo. Composición: Agua destilada 100 c.c. Disolver el cloruro de sodio en el agua, añadir el agar y dejar remojar 1 hora; disolverlo enseguida a 1 atm. 15 minutos. Filtrar por algodón, repartir y esterilizar a 1 atm. 15 minutos. Para obtener el agar-salino-sangre añádase 10 c.c. de sangre desfibrinada de conejo a 100 c.c. de agar fundido y mantenido a 50ºC. y para el agar-salino-sangre-maltosa se añade además 5 c.c. de maltosa al 20 p. 100. Ambos son repartidos en tubos y endurecidos en posición inclinada. Papa simple. Se usaron papas blancas escogidas. Después de lavarlas al chorro, fueron convenientemente mondadas manteniéndolas bajo agua; luego fueron cortadas en cubos de grosor y tamaño adecuados, los cuales fueron lavados en agua por varias horas, repartidos en tubos y esterilizados a 1 atm. 20 minutos. Caldo Carbohidratos y Leche tornasolada. Para el estudio de la acción fermentativa, se usó al comienzo caldo libre de azúcares (9), posteriormente usamos agua de peptona tal como la usada por Kauffmann y otros autores (10) para las bacterias del Grupo Intestinal. El carbohidrato fue disuelto en la proporción de 1 grm. p. 100, usándose como indicador tintura de tornasol (3.5 a 5 c.c. p. 100) y haciendo la esterilización al vapor por acción discontinua. Los carbohidratos empleados fueron químicamente puros (certificados por marcas como Grübler, Pfanstiehl Chem. Co.). La leche fue descremada, neutralizada y tornasolada en forma semejante. Después de repartirla en tubos se le esterilizó en la misma forma. Producción de Indol. Se emplearon cultives de 3 a 4 días de edad en agua de peptona de Durham (11) usando el indicador de Ehrlich (12) previa extracción con éter. Reacción de Voges Proskahuer y Methil Red. (V. P. y M. R.).Se emplearon cultivos de 4 días de edad en medio de Clark (13). Para demostrar la producción de acetilmetilcarbinol (V. P.) se añadió al cultivo un volumen igual de Potasa (KOH) al 10 p. 100, ordinariamente 5 c.c. de ambos; después de permanecer 12 a 18 horas en la estufa se hizo la lectura, la aparición de un color rojo naranja fue anotado como positivo. La reacción del rojo de metilo (M. R.) fue verificada añadiendo 5 gotas de solución hidroalcohólica al 0,04 p. 100 de dicho indicador al tubo de cultivo. Reducción de Nitratos a Nitritos. Se emplearon cultivos de 48 horas en caldo nitrato (14). La presencia de nitritos fue demostrada por la mezcla recientemente preparada de ácido sulfanílico y alfa naphtilamina, ambos en solución, acética (N/5). La aparición de un color rojo fue anotado como positivo. Formación de ácido nitroso. Esta prueba señalada por Fusco (15), fue verificada empleando cultivos en caldo (Standard) y añadiendo dos gotas de solución hidroacética de ácido sufanílico (2 grms. p. 300 c.c. de ácido acético al 4%) seguida de dos gotas de solución de alfa-naphtilamina (0.2 gms. p. 300 c.c. de ácido acético al 4%). La aparición de un color rojo indica reacción positiva. Producción de Hidrógeno sulfurado. Fue demostrado mediante el papel-acetato de plomo (método de Huddlenson) en cultivos sobre agar inclinado cambiándose el papel cada 18-24 horas durante algunos días. Reducción del azul de metileno. A un cultivo de 48 horas en caldo-Martín se añadió 1 gota capilar de azul de metileno al 1 p. 100, el volumen del cultivo no fue menor que 8 c.c. Los resultados fueron leídos algunas horas después, siendo hecha la última lectura a las 18 horas. Producción de Catalasas. El bacilo fue cultivado en agar inclinado 48 horas, se añadió entonces 0.5 c.c. de agua oxigenada. Cuando la reacción fue positiva se constató un burbujeo con formación de espuma. IV. CARACTERES MORFOLÓGICOS a) En los tejidos. En el hombre o los animales autopsiados la P. pestis puede ser encontrado en el bazo, hígado y nódulos pulmonares con sus típicas características de bacilo bipolar más o menos uniforme (Figs. 2 y 3); no sucede lo mismo en el bubón (Fig. 1), donde la morfología; salvo excepciones, es mas o menos variable, encontrándose frecuentemente numerosas formas redondas con o sin centro claro. Entre los innumerables bacilos que se observan en las preparaciones de bazo o ganglio, no es raro encontrar formas robustas y largas, teñidas intensa, débil o parcialmente con los colorantes usuales, que miden dos o tres veces el largo de un bacilo ordinario. En la Fig. 3 se podrá observar en una posición central superior derecha, una de ellas. En la Peste subaguda el bacilo se encuentra a menudo formando agrupaciones entre los elementos del pús. En los fragmentos de bazo o ganglio incluidos en parafina sólida se encuentran con frecuencia formas redondas con la parte central clara en los primeros días que siguen a la inclusión, posteriormente aparecen típicas formas de involución. b) En los cultivos. En los medios líquidos el bacilo desarrolla formando cadenas de variable longitud. Al envejecer estos cultivos (12 días) pueden originarse formas de involución. Los cultivos en medios sólidos corrientes no ofrecen particularidades morfológicas especiales. Tiende a formar agrupaciones, ordinariamente se deforma con el envejecimiento, la Fig. 12 muestra estas modificaciones. En la Fig. 8 se podrán observar típicas formas de involución provenientes de un cultivo en agar salado al 3.6 p. 100 que se compararán con los de la Fig. 9 que corresponden a la P. pseudotubercuosis rodentium desarrollada en este mismo medio. En agar-salino-sangre-maltosa el bacilo crece originando formaciones de aspecto característico: Enormes cadenas de elementos que pueden atravesar lado a lado el campo microscópico, generalmente entrelazadas y unidas por una sustancia envolvente que fija débil y metacromáticamente el azul de Loefflër. Figs. 5 y 6. Masas de innumerables organismos, incluidos en la misma sustancia. Fig. 7. En este medio es difícil sino imposible encontrar formas de involución. c) La cápsula de la P. pestis. Descubierta por Rowland (16), descrita como envoltura por Schutze (17) y como cápsula por Sokhey (18), este elemento estructural de la P. pestis, lo hemos encontrado en los tejidos de los animales, en los cultivos a 32 y 37ºC., en las cepas virulentas y avirulentas, en las variantes S o R, en las formas típicas así como en las de involución. Las óptimas condiciones nutritivas favorecen su desarrollo siendo las cepas virulentas las mejor encapsuladas; es frecuente encontrar entre las cepas stock sea cual fuera su virulencia algunas pobremente encapsuladas. Estamos de acuerdo con los autores que asignan a la cápsula un alto valor para el Diagnóstico. En los tejidos de los animales pestosos, según ya se ha dicho, el bacilo desarrolla su envoltura, pero esto puede tener variaciones o alternativas derivadas del desarrollo de anticuerpos. Para sustentar este punto hemos hecho experimentos con cepas encapsuladas, una virulenta CV11 y otra avirulenta K frente a sueros inmunes. Relatamos estos resultados: Emulsiones control de las cepas CV11 y K no tratadas por el suero: Encapsuladas. Los sueros probados fueron obtenidos de conejos inmunizados con productos de la desintegración de bacilos virulentos desarrollados a 33ºC (sueros 76 y 62); los mismos productos pero de bacilos avirulentos (suero 335) y con fluido envoltura (Schutze) de una cepa avirulenta (suero 343). Se determinó el título de aglutinación frente a la emulsión viva y standardizada a 600-1000 millones de organismos por 1 c.c. La incubación fue de 2 horas a 44ºC. seguida de una noche en la nevera. La encapsulación fue apreciada por el método de la tinta china. En los tubos en que la aglutinación fue total o parcial, se examinó el sedimento, directamente en los sueros muy diluidos y con previo lavado, en los no diluidos o poco diluidos. También se hicieron pruebas añadiendo al medio suero de caballos inmunizados con emulsiones vivas virulentas y avirulentas, y se observó que en el caldo suero antipestoso la P. pestis crece a 37ºC. desprovista completamente de cápsula; los resultados en agár fueron poco prácticos, pues fue requerida gran cantidad de suero para obtener cultivos descapsulados. V. CARACTERES CULTURALES Caldo-Martín. El desarrollo aparece en la masa del líquido como partículas tenues y blanquecinas. Desde las 12 horas a 30-32ºC. los grumos de mayores dimensiones van hacia el fondo, los más finos quedan en suspensión. Hay cepas que desde el comienzo desarrollan con manifiesta turbiedad; este punto fue estudiado por Bessonova y Lenkaia (19), quienes creen que es un fenómeno relacionado con la disociación del organismo. A los 4-5 días en cultivos que han permanecido inmóviles pueden encontrarse las formaciones clásicas descritas para este bacilo (Fig. 36), gran parte del desarrollo se encuentra en el fondo del tubo constituyendo un sedimento. Un estudio especial sobre la turbiedad fue llevado a cabo obteniéndose los siguientes resultados: De las 355 cepas probadas, 148 habían permanecido en stock 2 a 3 años, mientras que las restantes fueron estudiadas poco tiempo después de su aislamiento. Son comprendidas entre estas últimas (207) 90 cepas obtenidas en el diagnóstico de rutina y que no figuran en el total del presente trabajo. La turbiedad positiva en la P. pestis se mantiene por algunas generaciones, la ligera puede desaparecer de un subcultivo a otro. Medios sólidos. Los medios bacteriológicos corrientes distan mucho de ser óptimos para el desarrollo de la P. pestis aunque contengan sustancias iniciadoras de crecimiento como las sales de hierro y cobre, sulfito, etc., (20). Los factores nutritivos y otros agentes ejercen manifiesta acción modificadora sobre la colonia de la P. pestis; por esto las descripciones clásicas de la colonia necesitan ser revisadas a la luz de los conocimientos sobre el óptimo desarrollo de este organismo. Variaciones en la cantidad de medio nutritivo ejercen modificación en la morfología de la colonia. En las Figs. 15 y 24 se podrán observar colonias de 16 a 18 horas de 32ºC., las primeras desarrolladas en una placa con 10 c.c. de agar-Martin y la otra con 24 c.c. del mismo medio. Mientras que las primeras han desarrollado fleco las segundas no. Un resultado semejante se puede observar con Colonias de mayor edad, las Figs. 22 y 23 de 27 horas de edad a 32ºC. ilustran a este respecto. En los 4 casos se trata de una misma cepa, lo cual demuestra que la formación de fleco parece estar relacionado con las probabilidades nutritivas de la colonia; Burgess (21) indicó que la adición de sulfito de sodio estimula el desarrollo de fleco. Siguiendo las modificaciones experimentadas por el envejecimiento de las colonias se observa que aquellas que crecen concurrentes sufren pocas modificaciones, no sucede lo mismo con las que lo hacen aisladamente, las que presentan desde los 4 días surcos radiados (Fig. 17) zonas circulares concétricas alrededor de un cono central (Fig. 21), adoptando a los 7 días el aspecto de flor. Cuando un medio nutritivo promueve el crecimiento en forma rápida y copiosa, las colonias pueden presentar o no esboso de fleco, en cambio ellas son acuminadas, notablemente densas, algo consistentes y de considerables dimensiones a las 48 horas a 30 ó 32ºC. Hemos desarrollado el estudio de un medio descrito en la parte III con el nombre de agar-Martin-infusión-extracto hepático. En este medio la P. pestis crece copiosamente y en muchos casos da un desarrollo adherente a las 48 de incubación a 32-37ºC. Los bacilos desarrollan su envoltura en forma igual como en los tejidos animales. El rendimiento en desarrollo es tan alto que permite obtener fácilmente emulsiones fuertemente concentradas. Los siguientes experimentos así lo demuestran: También la temperatura ejerce acción sobre el desarrollo de la P. pestis en el agar-Martín-infusión-extracto hepático. Describimos los resultados obtenidos con 4 cepas: A este respecto Sokhey llama la atención sobre el pequeño tamaño de los bacilos desarrollados a 26ºC. La Cuenta de bacterias fue hecha por escala de opacidad de Mc. Farland; los cultivos se hicieron en placas de Roux conteniendo un volumen igual de medio y sembradas con un inóculo de 4 c.c. de cultivo en caldo-Martín desarrollado a 32ºC. durante 48 horas; la incubación se hizo invariablemente a 32ºC., durante 48-50 horas, y la cosecha fue estandardizada a un volumen fijo por placa. Las emulsiones de cultivo a 32-37ºC. son marcadamente mucosas y estables, las de cultivos a 26ºC no presentan estos caracteres. El desarrollo colonial puede ser obtenido en este medio (agar-Martín-infusión-extracto hepático) sin adición de substancias iniciadoras del crecimiento. Referimos el siguiente experimento como una demostración: Un cultivo en caldo de 24 horas a 37ºC. fue diluido a 1:10,000 y 5 gotas de esta dilución fueron sembradas en cada placa. Sólo han sido referidos algunos de los experimentos realizados que han servido para fundamentar el valor de este medio para el estudio de la P. pestis. Agar-sangre humana. En este medio se observa en los primeros días un desarrollo más o menos apreciable, a partir del 3er. día de incubación a 32-37ºC. aparece alrededor de las colonias un halo transparente similar a la hemolisis (Fig. 29). La P. pseudotuberculosis rodentium y la P. séptica pueden producir otro tipo de modificaciones transitorias en el medio pero nunca un halo permanente, amplio y definido. Este fenómeno tiene utilidad práctica en el Diagnóstico: Agar-salino-sangre-maltosa. El bacilo puede adaptarse a este medio después de trasplantes repetidos, entonces el desarrollo se hace marcadamente ostensible, en las cepas sembradas por primera vez éste puede ser insignificante apreciándosele mejor en el agua de condensación. Cepa C19 Trasplantes verificados entre set. 14 y dic. 9 (1937): 42 Cuyes inoculados por escarificación, en el mismo lapso de tiempo: 23 Resultado de la autopsia: Todos murieron con lesiones de Peste aguda entre 3 a 10 días. La cepa C19 aislada de sangre de cuy pudo mantener su virulencia alta después de 42 trasplantes diarios, interdiarios o cada tres días en el medio agar-salino-sangre-maltosa; después de 42 horas el medio toma un color bruno cepia. En el agar-salino-sangre sin maltosa no se producen las modificaciones señaladas y el bacilo puede también adaptarse a él y mantener alta su virulencia. Papa simple. Para obtener desarrollo visible en éste medio es necesario incubar los cultivos a bajas temperaturas y a largo plazo (12 días) (24), cerrando herméticamente los tubos para evitar la desecación, aún así no siempre se obtienen los mismos resultados con una cepa de una resiembra a otra. La siembra debe hacerse con una asa bien cargada con cultivo joven desarrollado en agar. Cuando el desarrollo ocurre puede notarse en algunas cepas una discreta pigmentación doradocepia o amarilla, este hecho está de acuerdo con las observaciones de Bessonova (25). Algunas cepas pueden llegar a adquirir el aspecto que se observa en las Fig 36a. y 38. Los resultados obtenidos con 320 cepas son los siguientes: Gelatina. Ninguna de las cepas de P. pestis, P. pseudotuberculosis rod y P. séptica licuaron la gelatina. A los 10 o más días de desarrollo la P. pestis puede emitir prolongaciones arborescentes a ambos lados del trayecto de la picadura; la P. pseudotuberculosis rod y las P. sépticas también pueden emitirlas pero no en la extensión y abundancia de la P. pestis, habiendo cepas incapaces para producirlas: Cepas estudiadas: P. pestis 128 VI. CARACTERES BIOQUÍMICOS Se ha buscado en la acción fermentativa y otras propiedades bioquímicas de la P. pestis, reacciones constantes y características para el Diagnóstico y diferenciación de la misma. Muchos autores en distintos países del mundo se han ocupado de este asunto debido a que la P. pestis y pseudotuberculosis rodentium son organismos tan afines que pueden originar verdaderos problemas de diferenciación con trascendencia epidemiológica cuando se trata de la Peste en los roedores. Este estudio lo hemos dividido en grupos para dar una idea aproximada y de conjunto sobre el comportamiento bioquímicos de las cepas aisladas en el Perú. La ferrnentación de carbohidratos se hizo con formación de ácido, nunca de gas; las incubaciones se hicieron a 32ºC. leyéndose resultados a las 24-48 horas y 4, 8 y 15 días, en algunos casos el número de observaciones en el mismo lapso de tiempo fue mayor. La glucosa y la levulosa fueron atacadas mas pronto que la maltosa y la manita por la P. pestis. La salicina fue fermentada solo desde los 2 a 3 días, no habiendo sido atacada por ninguna de las 9 P. séptica. La leche tornasolada fue directamente acidificada por las 291 cepas probadas mientras que todas las 6 P. pseudotuberculosis rod la alcalinizaron desde las 48 horas. Algunas cepas de P. pestis fermentaron ligeramente xilosa. Según se demuestra en el Cuadro Nº 2, 100 a 265 cepas de P. pestis probadas resultaron sin capacidad fermentativa para la sacarosa, rafinosa, dulcita, glicerina, inosita y dextrina; excepto 2 que atacaron tardíamente sacarosa. Las 6 P. pseudotuberculosis rodentium fermentaron glicerina en forma rápida o lenta y las 9 P. séptica, atacaron sacarosa invariablemente. Esto indica que la glicerina y la sacarosa son pruebas de valor en la diferenciación. La acción variable de la P. pestis para la rhamnosa fue demostrada estudiando 6 cepas que habían sido encontradas con acción positiva, probándolas en dos oportunidades mas con un período de 15 días entre una y otra prueba; por otro lado, otro lote de 104 cepas fue minuciosamente seguido a través de los mismos períodos de incubación; siendo todas ellas cepas stock conservadas a 10ºC. en agar sellado y con trasplantes cada 4 meses durante 2 a 3 años. Los resultados se relatan en el Cuadro Nº 4. El grupo de carbohidratos estudiados en los Cuadros 3 y 4 han sido atacados en forma variable por las cepas de P. pestis; así la rhamnosa fue fermentada entre 4 y 23 días por muchas de ellas, siendo las cepas que la atacan mas rápidamente las que mantienen este carácter más o menos fijo, no sucede lo mismo con las que la atacan tardíamente que pueden perder el carácter de una resiembra a otra. Es de notar que entre el stock hay muchas mas cepas con acción fermentativa para está carbohidrato que entre las cepas de reciente aislamiento. 5 de las P. pseudotuberculosis rod. atacaron este carbohidrato desde las 24 horas y una de ellas resultó negativa. La lactosa fue atacada tardía y débilmente por un pequeño número de cepas pestosas (stock), en algunos casos la acidificación fue transitoria, en cambio, la trehalosa lo fue por un porcentaje mucho mayor de cepas de P. pestis. La arabinosa que debía figurar entre los carbohidratos siempre atacados, no fue fermentada por un pequeño número de cepas pestosas. La sorbita que es considerada como negativa fue atacada en forma débil y transitoria por algunas cepas de P. pestis. Con excepción de una cepa entre las 148 estudiadas, la P. pestis reduce los nitratos a nitritos. Una de las 6 P. pseudotuberculosis rod. resultó negativa mientras que las 9 P. séptica resultaron invariablemente reductoras. La prueba de Fusco, o producción de ácido nitroso, propuesta para la diferenciación de la Peste, no siempre es negativa para todas las P. pseudotuberculosis rod. y P. séptica, en cambio las 52 cepas de P. pestis que fueron probadas dieron reacción positiva. El azul de metileno fue reducido débilmente por 3 cepas y acentuadamente por 1 entre las 148 cepas stock de P. pestis estudiadas. Entre las P. pseudotuberculosis rod. hubo una negativa mientras que todas las P. séptica redujeron con mayor o menor intensidad. La producción de SH2, depende de la base nutritiva en que se desarrolla la P. pestis; mientras que entre 105 cepas estudiadas en agar-Martín solo 3 produjeron ligeramente SH2; en el agar-Martín-infusión-extracto hepático, entre 381 cepas pasadas por este medio, 128 produjeron acentuadamente SH2, 132 en forma ligera y solo 21 resultaron negativas. La temperatura de incubación tiene influencia decisiva en la producción de SH2, a temperatura ambiente 9 cepas probadas fueron negativas, mientras que a 37ºC. 8 entre 11 cepas fueron positivas, dos ligeras y solo una resultó negativa. Por último, en el Cuadro Nº 6. se registran los resultados de las pruebas de acetil-metil-carbinol (V. P ), indol, reacción del rojo de metilo (R. M.), producción de catalasas, desarrollo en el medio de Kosercitrato y en el de ácido úrico (Ac. U). Estos resultados demuestran que la P. pestis no.produce indol ni acetil-metil-carbinol, que la reacción del rojo de metilo es positiva, produce constantemente catalasas y no crece en el medio de citrato de Koser o en el de ácido úrico; sin embargo se constató un ligerísimo desarrollo en el medio de Koser para algunas cepas, siendo comprobada la presencia del organismo, por observación microscópica. VII. DISCUSIÓN Desde los comienzos de la era bacteriológica de la Peste hasta nuestros días, las descripciones de los caracteres culturales y la morfología de la colonia se han basado en hechos y observaciones constatados en cultivos realizados en medios bacteriológicos corrientes. En tales medios no se ofrecen las óptimas condiciones exigidas por este bacilo para su desarrollo, él crece lenta y discretamente, en el borde de su colonia se forma un ancho ribete o fleco que ha sido mirado como característico en las descripciones clásicas. El medio que hemos descrito para el óptimo desarrollo del bacilo, no solo se caracteriza por su alto rendimiento proliferativo sino por que ofrece una nueva base para el estudio de su colonia que conducirá tarde o temprano a la aclaración definitiva del problema un tanto oscuro de la disociación bacteriana de la P. Pestis Por otro lado, medios como el agar-salino-sangre-maltosa que no teniendo valor proliferativo alguno y originando cultivos con curiosas agrupaciones de bacilos, conservan alta e intacta su virulencia, todo lo cual da una idea de la compleja biología de este organismo. Trabajos realizados por Rao demuestran que ciertos amino-ácidos como prolina, fenilalanina y cistina son necesarios para el crecimiento del bacilo (26); factores como la hematina, cccymaza, ácido nicotínico y thiamina (27) estimulan la respiración y aceleran el desarrollo; encuentra también este autor que la P. pestis tiene capacidad para oxidar ciertas sustancias como carbohidratos, amino-ácidos y ciertos ácidos orgánicos (28). Estudiando Sokhey (29) un medio de elección para el crecimiento del bacilo, encuentra que el agar ordinario no es el óptimo, y señala al agar sangre (37ºC.) como medio de elección. Cuando el bacilo encapsulado se pone frente a suero, antipestoso (antibacteriano o antienvoltura) se ocasiona la pérdida de la cápsula, fenómeno que lo hemos designado con el nombre de descapsulación. Si el bacilo se desarrolla a 37ºC. en caldo-suero antipestoso el desarrollo de la cápsula se inhibe y el bacilo crece con los caracteres de los cultivos a 26ºC. Es necesario pues precisar el fenómeno de la aglutinación del cuerpo bacilar, si éste existe, y diferenciarlo del fenómeno de la descapsulación. Las emulsiones de bacilos desarrollados a 26ºC son inestables; las de los desarrollados a 37ºC. son estables, se deduce de todo esto que el fenómeno de la descapsulación rompe la estabilidad de las emulsiones encapsuladas de la P. pestis. El estudio bioquímico que hemos realizado revela que la actividad metabólica de la P. pestis está sujeta a variaciones, según la cepa y según las condiciones básicas del desarrollo. Esta tendencia de la P. pestis a la variación en el orden metabólico puede guardar concordancia con las variaciones observadas en otros aspectos biológicos del organismo. CONCLUSIONES El medio agar-Martín-infusión-extracto hepático promueve, exalta y acelera el desarrollo de la Pasteurella pestis. En este mismo medio un fuerte porcentaje de cepas producen hidrógeno sulfurado en forma notable o intensa. El agar-salino-sangre-maltosa puede mantener alta e intacta la virulencia. El bacilo crece en este medio formando extensas cadenas y densas agrupaciones. El suero antipestoso produce la descapsulación de la Pasteurella pestis o inhibe su encapsulación cuando el suero se añade al medio de cultivo. La formación de un halo tipo hemolisis en las placas de agar-sangre humana es un fenómeno constante de la Pasteurella pestis. BIBLIOGRAFÍA 1. D. H. BERGUEY, R. S. BREED & E. G. D. MURRAY: Manual Det. Bact., 5ª ed., p. 289, 1939. [ Links ] 2. W. W. TOPLEY & G. S. WILSON: The Principles Bact. and Inmunol. 2ª ed., p. 605, 1938. 3. H. SCHUTZE: Brit. Jour. Exp. Path., t. 13, p. 289, 1932. 4. S. S. BHATNAGAR: Ind. Med. Res., Nº 1, p. 1-17, 1940. 5. P. HADUROY: Diccionaire des Bacteries Pathogenes, Paris, 1ª ed., p. 162, 1937. 6. J. D. LONG: Bol. of. San. Panam., Nº 1, p. 11, 1934. 7. H. COLICHÓN: Acerca de la Biología y Diferenciación de la P. pestis, Tesis de bachiller, Facultad Medicina, Lima, 1938. 8. L. MARTIN: Bibl. Therap. Batherotera. Vaccin. Serotherap., 1ª ed., p. 1936, Paris, 1909. 9. H. ZINSSER & S. BAYNE-JONES: A Textbook of Bacteriolg. 7ª ed., p. 1057, New York-London, 1935. 10. CH. BERGE & J. FAUCONNIERE: Arch. L Institut. Pasteur Tunis, t. 30, p. 103, 1941. 11. J. A. KOLMER & F. BOERNER: Approv. Lab. Tech., 2ª ed., p. 356, 1938, New York-London. 12. Idem, Idem. p. 406. 13. Idem. Idem. p. 357. 14. Idem. Idem. p. 356-407. 15. G. Fusco: Bull. Off. Int. Hyg. Pub., t. 14, Nº 4, p. 612, 1932. 16. S. ROWLAND: Jour. of Hyg., t. 13, Plague supl. III, p. 418, 1914. 17. H. SCHUTZE: Brit. Jour. Exp. Path., t. 13, p. 284, 1932. 18. S. S. SOKHEY: Journal Path., and Bact., t. 51, Nº 1, p. 97-103, 1940. 19. A. BESSONOVA & G. LENKAIA: Rev Micr. Epidem. Saraoff, Nº 3, p. 270, 1929. 20. H. SCHUTZE & M. A. HASSANEIM: Brit. Jour. Exp. Path., t. 10, p. 204, 1929. 21. A. S. BURGESS: Jour. of Hyg. Cab., t. 30, p. 165, 1930. 22. F. M. HUNTOON: Jour. of Inf. Dis., t. 33, p. 169, 1918. 24. J. M. LIGNIERES: Annales de LInst. Pasteur, t. l5, p. 808, 1901. 25. A. BESSONOVA: Giorn. di Batteriol. e. Inmunol., t. 16, Nº 5, p. 754-760, 1936. 26. M. RAO (Sadashiva): Ind. Jour. Med. Res., (27 july), p. 75, 1939. 27. M. RAO (Sadashiva): Ind. Jour. Med. Res., t. 27, Nº 4, p. 883-846, 1940. 28. M. RAO (Sadashiva): Ind. Jour. Med. Res., t. 27, Nº 3, p. 617-626, 1940. 29. S. S. SOKHEY: Ind. Jour. Med. Res., t. 27, Nº 2, p. 321-329, 1939.
De origen murino 49
De piojos 1
De origen rnurino 1
B. pseudotuberculosis rodentium (N2200) Group II 1093/25.
B. pseudotuberculosis rodentium (N1780) Group II tipe B cll.
B. pseudotuberculosis rodentium (N1894) of Monkey.
B. pseudotuberculosis rodentium (N2496) Group II tipe A turkey 313
B. pseudotuberculosis rodentium (N3245) Group III.
B. pseudotuberculosis rodentium (N941) Group I tipe B. ginea-pig3A.
Pasteurella of mouse 49 (N948)
Pasteurella bovisépticum (N929)
Pasteurella lepisépticum (N1876)
Pasteurella suiséptica (N1737)
Agua corriente 500 c.c.
Extracto de carne (bacteriolog.) 3 c.c.
Cloruro de sodio q. p. 5 c.c.
Extracto hepático 80
Agar-Martín-infusión 100 c.c.
Sangre humana desfibrinada 5 c.c.
Cloruro de sodio (q. p.) 0.7 a 0.8 grms.
Agar lavado 14 grms.





P. pseudotuberculosis rod 6
P. séptica 5 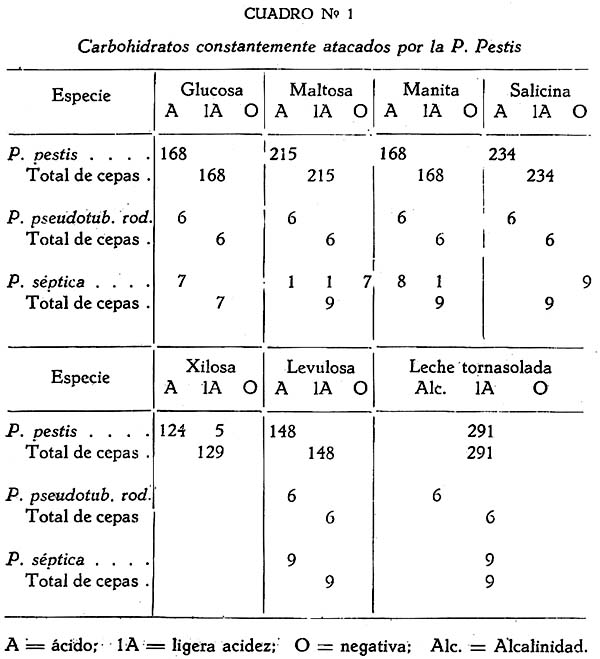
![]()