










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
Print version ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.2 no.1 Lima Mar. 1943
TRABAJOS ORIGINALES
Observaciones sobre la Bartonelosis en el Cuy (Cavia Porcellus)
N. Van Der Walle1
1 Instituto Nacional de Higiene y Salud Pública, Lima.
RESUMEN
1. La Haemobartonella tyzzeri ha sido observada microscópicamente en 3 cuyes machos de 10 cuyes esplenectomizados procedentes de la zona verrucógena del Rímac.,, (5 machos, 5 hembras), mientras que 3 machos y 1 hembra de Lima resultaron negativos.
2. Una vez se ha cultivado la H. tyzzeri en el medio semi-sólido de NOGUCHI a 28ºC.
3. Sangre de un cuy naturalmente infectado fué inyectado intraperitonealmente a un cuy esplenectomizado de Lima; a las 2 semanas las bartonellas eran visibles en la sangre. De esta manera fueron inoculados 6 cuyes en pasajes sucesivos, siempre con resultados positivos. El porcentaje de glóbulos infectados (± 0.05 % en el cuy original), subió durante estos pasajes, hasta un máximo de 6.25%. El período de incubación bajó en los dos últimos pasajes hasta 1 semana. Tres de los 6 animales mostraron una ligera anemia (con 3.48, 3.74, y 3,19 millones de hematíes por mm.3, respectivamente) y otro (el del segundo pasaje) una anemia fuerte (1.25 millón), con macrocitos, normoblastos y un alto porcentaje de reticulocitos. Está comprobado pues, que la virulencia de la H. tyzzeri puede ser aumentada.
4. Se ha estudiado el cuadro hemático con los resultados siguientes. El diámetro medio del glóbulo parasitado resulta por lo. general un poco más pequeño que el del no infectado, diferencia que por lo menos en un caso no se podría atribuir al azar.
En tres cuyes se observó un aumento de los hematíes, precediendo al descenso de éstos.
En algunos de los cuyes parasitados se notó una disminución del número de los leucocitos, seguido por una leucocitosis, la cual era unas veces acompañada, de un aumento del porcentaje de los granulocitos neutrófilos.
Eosinofílía durante o después de la infección fué observada en tres cuyes, monocitosis en dos.
Algunas veces se pudo poner de manifiesto una hipersegmentación del núcleo de los granulocitos.
5. Se realizó un estudio anátomo-patológico del hígado de algunos cuyes parasitados y del bazo de un cuy naturalmente infectado; también dé un pedazo del bazo, dejado con una esplenectomía incompleta en otro cuy experimentalmente infectado.
SUMMARY
1. Haemobartonella tyzzeri was observed in Giemsa-stained films of the blood of 3 male guinea-pigs out of 10 splenectomized animals coming from the verruga zone of the Rimac Valley. As 5 of these guinea-pigs happened to be males and 5 females, it is possible that in nature males are more frequently infected than females. Four splenectomized guinea-pigs from Lima (3 males, 1 female) have been repeatedly examine d with negative results.
2. After several failures H. tyzzeri was cultivated once from the blood of an experimentally infected guinea-pig in the semi-solid medium of NOGUCHI at 28ºC.
3. Blood of one naturally infected guinea-pig was injected intraperitoneally into a splenectomized guinea-pig from Lima; two weeks later the bartonellae were visible in its blood. This strain was carried through six, successive passages in splenectomized guinea-pigs. Three of the latter (males) came from Lima, the other three (females) from the Rimac Valley, but had been examined first during at least one month with negative results. Thus it seems that females from the verruga zone, are not immune against experimental infection with H. tyzzeri.
During these animal passages there was a decreased in the mean number of bartonellae per infected red cell (from 17 in the original guinea-pig to 11, 12, 7, 7, 2 and 6), as well as in the maximum observed for any red cell. (from 28 in the original guinea-pig to 6, 25, 16, 16, 6 and 8). However, the percentage of infected red blood corpuscles increased from 0.05% to 6.25% during these passages.
The period of incubation decreased from 17 days in the original guinea-pig to 1 week during the last two passages.
Three of the six experimentally infected animals showed a slight anemia, with respectively 3.48, 3.74 and 3.19 million red cells per cu.mm., while one (second passage) showed a severe anemia (1.25 million) with macrocytes, numerous normoblasts and 47% of reticulocytes.
This indicates that the virulence of H. tyzzeri can be increased.
4. The diameter of the infected erithrocytes was slightly smaller than that of the non - infected ones; it has been calculated that in at least one case this difference could not be explained by chance. (Cuadro 1).
In 3 infected guinea-pigs the red count increased in the first period of infection.
Some of the infected guinea-pigs showed a temporary leucopenia before bartonellae appeared in the blood; the infection period, however, was caracterized by leucocytosis, (Cuadro 2) which was in some cases accompanied by a relative increase of the neutrophiles.
Eosinophilia during or after the period of infection has been observed in 5 guinea-pigs but 2 of these cases could be explained by the presence of worms. (Cuadro 3).
In the blood of some infected guinea-pigs an increased nuclear segmentation of the neutrophiles was detected. (Cuadro 4).
5. A pathological study was made of the liver of the infected animals. The principal changes found were degeneration of the liver-cells, focal areas of necrosis and proliferation of connective tissue, (especially around the bile-ducts) and extensive phagocytosis of blood. corpuscles and blood pigment. No signs of inflamation were present.
At the autopsy of the sixth passage guinea-pig a small piece of the spleen was found, obviously the consequence of an incomplete splenectomy. This showed marked reduction of lymphoid tissue and hyperplasia of the pulp; hyaline degeneration surrounded the small arteries in the Malpighian bodies. Numerous macrophages contained brown pigment.
I. Introducción
En 1938 WEINMAN y PINKERTON describieron con el nombre de Bartonella tyzzeri una nueva especie de bartonella, encontrada en cuyes procedentes del Perú. (12, 18). La hallaron en la sangre de 4 de 23 cuyes esplenectomizados. Este organismo se presentaba, ya sea en forma bacilar, con una longitud de 0.8 - 4 micras o cocóide con un diámetro de 0.2 - 0.3 micras, coloreable con Giemsa o May-Grunwald Giemsa. El número de hematíes infectados era siempre pequeño, más o menos de 0.1 %, los que aparentemente no sufrían alteraciones en cuanto a su forma, dimensiones y afinidad cromática. Los animales infectados soportaban la infección sin dar muestras de anemia o presentar síntomas especiales. Dichos autores nunca lograron poner de manifiesto la infección con bartonellas en cuyes procedentes de Boston.
Ellos consiguieron cultivar la Bartonella tyzzeri en el medio semi-sólido de Noguchi a 28ºC., del mismo modo que en el medio de Zinsser, Wei y Fitzpatrick, usado corrientemente en el cultivo de rickettsias; ellos emplearon en este caso túnica vaginal de cuyes normales. Con sus cultivos consiguieron infectar cuyes esplenectomizados, no así a monos (Macacus rhesus) que, como es sabido, son sensibles a la inoculación con Bartonella bacillitormis, descartando así la posibilidad que los cuyes pudieron ser, portadores de la Enfermedad de Carrión.
En 1939 TYZZER y WEINMAN proponen el nombre de Haemobartonella para las bartonelas que se multiplican sólo en la, sangre y no producen lesiones cutáneas, género éste que incluiría también a la B. tyzzeri. ( 16).
Anteriormente al trabajo concluyente de WEINMAN y PINKERTON, varios investigadores observaron microorganismos o elementos parecidos a bartonelas en los glóbulos rojos de cuyes esplenectomizados o sometidos a diversos tratamientos. Así entre otros, MARTIN MAYER en 1921 encuentra elementos parecidos a la B. bacilliformis en cuyes de Hamburgo con grave infección de Trypanosoma rhodesiense, después de ser sometidos al tratamiento con Bayer 205. (5, 7); posteriormente él y sus discípulos no lograron ver los mismos elementos en un gran número de cuyes esplenectomizados. (6). En 1930 NOÉ y PÁEZ observaron en Santiago de Chile en dos cuyes esplenectomizados, corpúsculos endoglobulares parecidos a bartonellas. (8, 14).
Al tratar de la H. tyzzeri es preciso tener en cuenta el hecho de que los hematíes de cuyes normales pueden también presentar de vez en cuando inclusiones semejantes a bartonelas, las que ya fueron observados por WENYON y LOW en 1914. (17) . SCHWETZ analizó en 1934 la literatura existente hasta entonces y concluyó de que mucho de lo descrito como bartonelas del cuy, debería ser considerado simplemente como inclusiones no patógenas. (18).
II. Observaciones en cuyes con infección natural
El trabajo de WEINMAN Y PINKERTON, hecho en Boston con cuyes llevados del Perú no ha sido continuado por otros según sabemos. Por eso juzgamos necesario repetir en el Perú tales investigaciones.
Empezamos con la esplenectomía de 10 cuyes procedentes de la zona verrucógena situada en el Valle del Río Rímac. Tres de estos cuyes eran de Tornamesa (altura de 1513 metros * ) y los siete restantes de Surco (2035 metros). Ambas localidades se hallan al lado de la línea del Ferrocarril Central que va de Lima hacia Oroya y distan respectivamente 76 y 91 kilómetros de la Capital.
En la sangre de 2 de los cuyes de Surco y de 1 de Tornamesa logramos encontrar microscópicamente la H. Tyzzeri; los tres animales infectados eran machos. Como de los 10 cuyes empleados la mitad eran machos, presumimos que la infección podría ser más frecuente en los animales de este sexo. Además teníamos presente el hecho de que MAR- MORSTON, GOTTESMAN y PERLA en sus estudios sobre la bartonela de la rata eligieron solamente animales machos, en vista de que en éstos la infección era más aguda (4). Tal vez pues, si hubiéramos usado en nuestro trabajo sólo cuyes machos, el porcentaje de infección obtenido (33 %) habría sido más elevado.
El 29 de Setiembre de 1941 tuvimos el agrado de ver por primera vez en un frotis de sangre del cuy Nº 1 los graciosos bastoncitos coloreados de rojizo- violeta con el Giemsa. Antes de ésto, hasta al día 5 del mismo mes, la sangre de este animal fué observada repetidas veces, con resultado negativo, por lo cual concluíamos que el período de incubación en este casó ha sido por lo menos de un mes. La infección fué posible constatarla microscópicamente durante 45 días. También en el cuy Nº 3, el período de incubación parece haber sido cuando menos de un mes, mientras que en, el cuy Nº 9 (de Tornamesa) sólo fué de 17 días. El grado de infección en este último animal fué más alto que en los dos anteriores. Aunque el número de los hematíes infectados era tan pequeño es decir, cuando más 0.05 % (**) - el promedio de bartonellas en los hematíes parasitados (***) alcanzó en el cuy 9 el valor de 17 y el número máximo encontrado en un glóbulo fué de 28. En el cuy Nº 1 estos valores fueron respectivamente 11 y 18; en el cuy Nº 3, 12 y 18. (Véase Tabla I).
Los cuyes parasitados no presentaron síntomas especiales; la temperatura rectal varió entre 37 y 40,5ºC., siendo la gráfica térmica idéntica a la de los cuyes que resultaron negativos.
El cuy 9 murió espontáneamente, el cuy 1 después de una punción del corazón y el Nº 3 después de una segunda inyección subcutánea con benzol-aceite de olivo, llevada a cabo con la intención de activar la infección latente y hacer visible de nuevo los parásitos ya desaparecidos de la sangre. [Este procedimiento que no tuvo éxito en nuestro caso, nos fué inspirado por los experimentos de ZINSSER y. CASTAÑEDA (21 ) en la rickettsiosis del cuy y de la rata], Ninguno de estos tres animales presentó anemia marcada. Lo mismo dicen WEINMAN y PINKERTON de sus cuyes parasitados, presumiendo dichos autores que esta ausencia de anemia fuera debido al escaso número de corpúsculos infectados. En todo caso este hecho debe extrañarnos, porque la bartonelosis en los animales (rata, perro) casi siempre está caracterizada por una anemia fuerte. (Pocas son las excepciones conocidas, por ej. la de Phodopus praedilectus, mencionada por WEINMAN en su tesis).
Además hemos examinado la sangre de 4 cuyes precedentes de Lima, con resultados negativos. (Tiempo de observación 4, 4, 5 y 6 semanas después de la esplenectomía, respectivamente). Tres de estos animales eran machos y uno hembra.
III. Observaciones experimentales
Desde el principio nos habíamos preguntado si sería posible, por medio de pasajes sucesivos, aumentar la virulencia de la H. tyzzeri, consiguiendo tal vez así anemia en los cuyes infectados. Teniendo presente esta idea, llevamos a cabo una serie de inoculaciones en 6 cuyes distintos, 3 de los cuáles eran de Lima y los otros 3 procedían de la zona verrucógena del Rímac. Estos últimos fueron observados de 3 a 6 meses después de la esplenectomía sin lograr poner de manifiesto en ellos la infección natural. (Véase Tablas II y III).
I. Cuy A (macho, procedente de Lima) fué observado durante 6 semanas después de la esplenectomía sin lograr ver bartonelas en la sangre. Después de 14 días de ser inoculado intraperitonealmente con 2 cc. de sangre infectada (0.05 % de hematíes parasitados) del cuy Nº 9. se logró ser bartonellas en la sangre. La infección fué más intensa (0.25 %) que en el cuy 9 y la cifra de eritrocitos descendió de 4.92 a 3.48 millones.
II. Cuy B (macho, procedente de Lima), con periodo de 13 días de observación después de la esplenectomía. A los 21 días de la inoculación intraperitoneal con 3 cc. de sangre del cuy A se logró poner de manifiesto las bartonellas. El porcentaje de glóbulos parasitados nunca fué mayor de 0.1 %. Los eritrocitos descendieron de 4.54 millones hasta 1.25 millón. En este momento nos fué imposible obtener sangre de la oreja, por lo cual hubo de punzarse el corazón, muriendo poco después el cuy.
El cuadro hemático demostró microcitos, macrocitos y numerosos normoblastos. (Fig. 9). El porcentaje de los reticulocitos (coloreados con azul - cresíl) fué de 47 % cuyes normales 3.7 %). Siendo el porcentaje más alto encontrado en nuestro después de un examen prolongado encontramos un hematíe con bartonellas. [Fenómeno que correspondería al observado en los perros (20) y en las ratas (17).]
En la autopsia se observó el hígado pálido y endurecido con focos hemorrágicos; la vesícula biliar contenía un gran coágulo y la cavidad abdominal una gran cantidad de líquido amarillo -oscuro, conteniendo bilirubina.
III. Cuy Nº 6. (Hembra, procedente de Surco), período de observación 5 meses después de la esplenectomia con resultados negativos. A las 2 semanas de la inoculación intraperitoneal con 3 cc. de sangre del cuy B se logró poner de manifiesto la infección con 0.5 % de glóbulos infectados. El número de eritrocitos descendió ligeramente, de 5.57 a 4.65 millones. Murió espontáneamente y en la autopsia mostró algunas alteraciones. (Edema gelatinoso subcutáneo en la región inguinal; glándulas inguinales aumentadas, hiperémicas. Gran cantidad de líquido amarillo - oscuro en la cavidad abdominal; en el hígado se encontraron manchas verdes).
IV. Cuy Nº 5. (Hembra, procedente de Surco), con 6 meses de observación después de la esplenectomia. También en este animal se vieron bartonellas después de 2 semanas de la inyección intraperitoneal con 3 cc. de sangre infectada del cuy Nº 6. La infección fué de 0.3 % de glóbulos parasitados; los eritrocitos descendieron de 6.01 hasta 3.74 millones., Aparentemente este animal murió a causa de una infección Secundaria.
V. Cuy Nº 10. (Hembra, procedente de Tornamesa) fué observado 3 meses después de la esplenectomía. A la semana de la inyección intraperitoneal con 1 cc. de sangre infectada del cuy Nº 5 se observaron las bartonellas. El porcentaje de hematíes parasitados ascendió en una semana de observación de 0.05 % a 6.25 %. Murió a consecuencia de una punción del corazón, evidentemente al principio de la enfermedad y sin dar todavía muestras de anemia. (En la autopsia se encontró además de una hematoma del pericardio, sólo una pequeña cantidad de líquido turbio en la cavidad abdominal).
VI. Cuy E. (Macho, procedente de Lima), observado 1mes después de la esplenectomía. A la semana de la inoculación intraperitoneal con 0.5 cc. de sangre del cuy Nº 10 se lograron ver bartonellas en la sangre con un porcentaje máximo de 0.05 %. Él número de hematíes después de la esplenectomía fué de 5.22 y descendió basta 3.19 millones. Al hacer la autopsia tuvimos la sorpresa de encontrar un pedacito del bazo (más o menos la 1/15 parte del órgano). Esperábamos en este sexto pasaje obtener una virulencia mayor del germen, lo que fatalmente no hemos podido conseguir, tal vez por la influencia del pedazo de bazo que se había dejado de extraer, [En este sentido son conocidos los experimentos, de Perla y Marmorston - Gottesman (10), que basta la presencia de la 1/4 parte del bazo para proteger a la rata contra la barlonellosis].
De los resultados de estas infecciones experimentales se puede deducir las observaciones siguientes:
1: Está probado que es posible transmitir la infección con H. tyzzeri de cuyes naturalmente infectados a otros no parasitados.
2: Se ha demostrado que cuyes hembras no parasitadas procedentes de la zona verrucógena, no son inmunes a la infección con H. tyzzeri.
3: Se ha puesto de manifiesto la posibilidad de aumentar la virulencia de la H. tyzzeri, de modo que condicione una anemia.
4: De las Tablas II y III se desprende que el promedio de las bartonellas por hematíe, no aumentó como. habría podido esperarse, con, los pasajes sucesivos. Pues, en el cuy 9, naturalmente infectado y del cual se partió dicho promedio fué de 17 y en los 6 cuyes de esta serie experimental fué sucesivamente de 11, 12, 7, 7, 2 y 6. Cosa idéntica se observó con el número máximo de bártonelas por eritrocito ya que en el cuy 9 fué de 28 y en los otros 6 animales experimentales de 16, 25, 16, 16, 6 y 8. (Naturalmente tales cifras no tienen sino un significado relativo, porque la selección de los glóbulos examinados es determinado por el azar). 1
5: En cambio en el porcentaje de glóbulos infectados se notó un marcado aumente. Así, de 0,05 % que era en el cuy 9, aumentó hasta 6.25 % al quinto pasaje. En los 6 cuyes de esta serie dicho porcentaje
varió de la siguiente manera : 0.25 %, 0.1%, 0.05 %, 0.3 %, 6.25 % y 0.05 %. La cifra baja observada en el último pasaje podría explicarse recordando que este animal no fué esplenectomizado completamente.
En otro sentido es notable que el cuy con anemia grave (B., segundo pasaje) nunca mostró un porcentaje mayor de 0.1 Este hecho induciría a pensar que la anemia en este case sería tal vez debida más a la virulencia del germen que a su número en los eritrocitos.
VI. Observaciones sobre la Haemobartonella tyzzeri
En los frotis, fijados con alcohol metílico o con May-Grünwald y coloreados, durante una hora con una solución de Giemsa al 1/20, se destacan claramente las bartonelas con un color rojizo-violeta sobre el fondo gris-rosado de los glóbulos parasitados. En el comienzo de la infección predomina la forma bacilar, es decir bastoncitos rectos o un poquito curvos, que tienen dimensiones casi iguales (1.2 a 2 micras); algunos tienen aspecto bipolar. Se ven muy pocas formas cocóidas (0.3 a 0.5 micras) en este período. El número de los organismos en cada eritrocito es todavía pequeño, por ej. 1 a 7. (Fig. 1 ). En la mayor parte de los casos las bartonelas son distribuidas bastante uniformemente en el hematíe, aunque de vez en cuando ellas se localizan más en una mitad del glóbulo o en un círculo cerca de la periferie. (Fig. 3). En el caso que un glóbulo esté parasitado por sólo una bartonella, se ve con frecuencia que ésta se halla exactamente al borde. A medida que la infección progresa, aumenta el número de las bartonelas en los eritrocitos (Fig. 4); el número máximo que observamos fué de 28. También en este período pueden presentarse la mayoría de las bartonellas como bastoncitos de longitud más o menos igual, pero muchas veces aumenta relativamente el número de las formas cocóidas. (Fig. 5). La impresión general de las láminas es la de una gran regularidad, la cual en algunos casos puede ser tan pronunciada que sería fácil confundir estos microorganismos con grahamellas. En un período más avanzado, se observan formas más largas (hasta 4 micras), rectas, curvas o serpenteadas (Fig. 6); algunos organismos están pegados en la totalidad o parcialmente a sus vecinos; los contornos resultan menos nítidos de modo que es algo difícil distinguir los diferentes elementos. Entonces ha desaparecido el aspecto de regularidad de los primeros períodos.
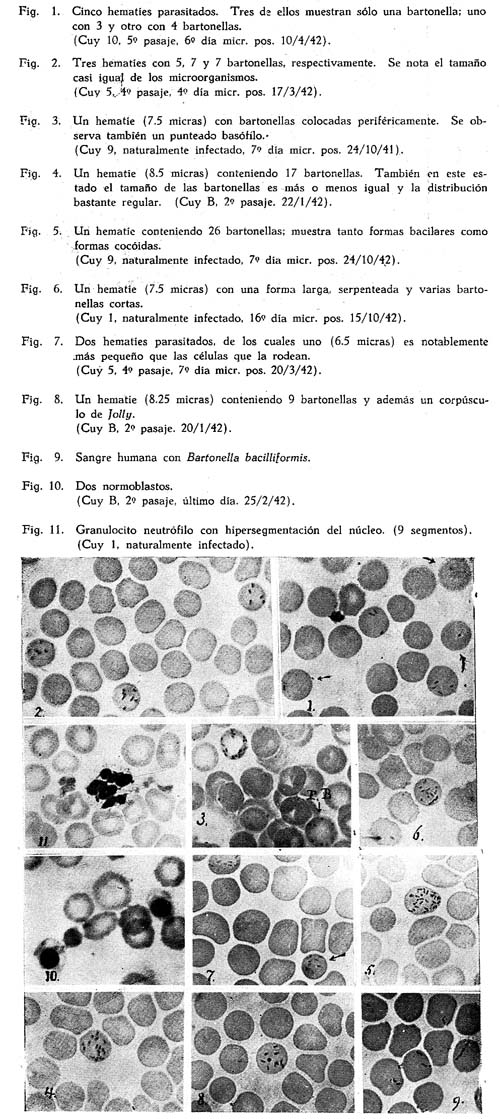
Al comparar los frotis de sangre del cuy infectado con H. tyzzeri con otros de sangre humana parasitada con la B. bacillifornis, nos parecía que las formas del último organismo varían más en cuanto a su longitud y aspecto. Muchas veces se ven formas en V o Y, lo que se encuentra sólo rara vez en las láminas con H. tyzzeri. Cadenas de 3 hasta 5 elementos se observan con frecuencia en la infección con B. bacilliformis y casi nunca en la con H. tyzzeri. (Fig. 9).
También la distribución en los glóbulos parece menos regular en la bartonella del hombre que en la del cuy. (9, 15).
Hemos medido 200 ejemplares de la H. tyzzeri, encontrados al azar en distintas láminas y también igual número de B. bacilliformis en otra lámina de sangre humana. Resulta de estos cálculos que en nuestro caso se ha mostrado que la longitud media de la H. tyzzeri es más pequeña que la de la B. bacilliformis.

Sucede a veces que un glóbulo parasitado con pocas bartonellas está parcialmente cubierto por otro que no tiene parásitos; entonces se puede observar uno o dos organismos a través del segmento. También ocurre de vez en cuando que uno de los bastocitos pasa por el borde del hematíe o se halla libre junto a la periferie. En otros casos tuvimos la impresión que una o dos bartonelas, se habían deslizado encima de otro glóbulo contiguo. Tales láminas nos sugerían la idea que las bartonelas están colocadas encima y no dentro de los glóbulos rojos. [Semejante conclusión hizo también ALDANA (1) ].
Por lo general el número de los hematíes infectados era muy pequeño, exactamente como WEINMAN y PINKERTON describen. Hay muchos Campos microscópicos en los cuales no se ve ningún corpúsculo parasitado; pocas veces se encuentran dos en un campo, rara vez tres. (Fig. 2). La única excepción era la sangre del cuy 10 (quinto pasaje) con una infección de 6.25 % de los hematíes, cada uno con muy pocas bartonellas. (Fig. 1).
Después de haber alcanzado un cierto máximo, disminuye el número de los hematíes parasitados, los cuales son finalmente tan escasos que se hace necesario buscar diez o más minutos antes de encontrar los parásitos. Es muy posible que también al principio de la infección exista una tal escasez de glóbulos infectados, de modo que cabría la posibilidad que el período de incubación fuera más corto que él que hemos notado, especialmente en los cuyes naturalmente infectados. (En este sentido recordamos la observación de MAYER (7) que también en la rata puede existir en el comienzo de la infección una gran escasez de parásitos y que es preciso buscar mucho tiempo antes de encontrarlos). No existe un paralelismo entre el número de las bartonellas en los hematíes y el número de los glóbulos parasitados. Al contrario, a menudo se observa que las primeras aumentan, mientras que, el porcentaje de los glóbulos infectados queda más o menos el mismo o disminuye. (Lo que se puede desprender claramente dé las Tablas adjuntas). De esta discordancia resulta a nuestra parecer que las bartonellas se multiplican en o más probablemente encima de -los hematíes.
En la sangre de algunos cuyes no esplenectomizados encontramos unas veces bastoncitos y gránulos, que tenían más o menos el aspecto de bartonellas pero de un color violeta pálido; el número de hematíes que contenían tales elementos era siempre muy pequeño, cada uno teniendo cuando más tres de éstos. Presumimos que tales elementos corresponden a aquellos descritos por WENYON y LOW. En este sentido era muy interesante la sangre del cuy Nº 7, procedente de Surco. Una semana después de la esplenectomia vimos en casi la mitad de los eritrocitos bastoncitos bipolares y diplococos, muy finos y de color violeta pálido, algunos de forma muy semejante a la de bartonelas. Probablemente también estos elementos tenían un carácter no específico y no patógeno, pues sin éxito ninguno inyectamos la sangre de este cuy en otro animal esplenectomizado y negativo. En este momento no habíamos visto todavía H. tyzzeri y teníamos duda sobre la naturaleza de estos elementos. Pero esta duda desapareció inmediatamente, cuando, observamos por primera vez las verdaderas bartonellas, las cuales tienen un aspecto tan característico que difícilmente uno podría engañarse, después de haberlas visto una vez.
Después de varios ensayos con resultados negativos, el Dr. A. HERRER, con su gran experiencia con cultivos de B. bacilliformis, logró cultivar la H. tyzzeri de la sangre del cuy B. en el medio semi-sólido de NOGUCHI y a 28ºC. Las colonias fueron visibles del 5º al7º das con la lente (20X ). Tenían la forma de puntitos gris-blancos rodeados por un halo semitransparente típico, exactamente como las de la B. bacilliformis. Se hallaban esparcidas en una zona de más o menos 1 cm. a partir de la superficie del medio. Posteriormente las colonias originales aumentaron de tamaño, aunque sin exceder de 1 mm. de diámetro, y se multiplicaron. Las resiembras de este cultivo fueron positivos hasta él tercer pasaje.
En el campo obscuro las bartonellas eran visibles como bastoncitos brillantes con, un centro negro característico, parecido a la bartonella del hombre, pero faltó la movilidad que posee este último organismo.
V. Observaciones sobre el cuadro hemático
A. Eritrocitos.
Los eritrocitos parasitados no se distinguen ni en forma ni en color de los no infectados. De vez en cuando se encuentra un microcito parasitado (Fig. 7). Rara vez se ven bartonellas en un hematíe que contiene un corpúsculo de JOLLY (Fig. 8); aun más raro es la infección de un glóbulo policromático.
HURTADO et al. (2) han constado que en la bartonellosis humana el diámetro de los glóbulos parasitados es por lo general un poco menor que él de los no infectados; en algunos casos esta diferencia está claramente pronunciada. Por eso hemos medido el diámetro de 100 glóbulos parasitados y también de 500 no infectados en frotises de 6 cuyes. (En la sangre del cuy Nº 10, que era fuertemente infectada, 300 de cada especie). Resulta que en todos estos casos el diámetro medio del glóbulo parasitado es un poco más pequeño que él del río parasitado y que por lo menos en el cuy Nº 1, esta diferencia no se podría atribuir al azar. (Véase cuadro I).
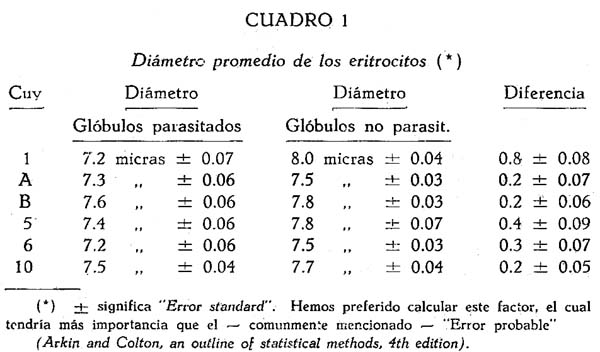
El número de los glóbulos rojos, contado antes de la esplenectomia varió entre 3.94 y 6.61 millones.
Estas cifras fueron determinadas en 15 cuyes de Lima, siendo los valores parciales de: 3.94 - 3.98 - 4.58 - 4.58 - 4.58 - 4.67 - 4.78 - 4.79 - 4.94 - 4.99 - 5.06 - 5.11 - 5.16 - 5.50 -5.51.
Promedio: 4.81 ± 0.14.
Las mismas cifras determinadas en 15 cuyes de Surco y de Tornamesa fueron de: 4.67 - 4.87 - 5.08 - 5.09 - 5.36 - 5.47 - 5.48 - 5.67 - 5.67 - 5.79 - 5.86 - 5.97 - 6.26 - 6.45 - 6.61.
Promedio: 5.62 ± 0.13.
Diferencia del promedio en ambos grupos 0.81 ± 0.19.
Aparentemente cuyes de las regiones elevadas presenten una ligera poliglobulia, comparable a lo que es conocido en el hombre.
Inmediatamente después de la operación hecho en ayunas - baja el número de los hematíes ligeramente, evidentemente causado por pérdida de sangre. En cuatro de nuestros cuyes hemos podido observar, que después este número aumenta por encima de la cifra original, antes de volver a descender, tal vez causado por algún estímulo de las bartonelas en los órganos hemopoyéticos. Este fenómeno fué especialmente pronunciado en los tres cuyes naturalmente infectados Nº 1, 3, y, 9, en los cuales el número de los hematíes que era de 5.97, 5.36 y 4.78 millones después de, la esplenectomía, subió hasta 6.46, 6.36 y 5.74 millones respectivamente, (Véase Tabla I).
[NEUMANN en la rata encontró un ligero aumento al comienzo de la infección (17). Parece también que en la bartonellosis humana un tal aumento precede en raros casos a la anemia, (3)].
En la sangre de los cuyes encontramos antes de la esplenectomía algunas veces hematíes policromáticos, excepcionalmente corpúsculos de JOLLY y rara vez punteado basófilo. En cuanto a estos últimos elementos, sólo la sangre del cuy 9 - infectado naturalmente - hacía una excepción, conteniendo numeroso punteado basófilo (Fig. 3). Después de la operación aparecen en todos los animales los corpúsculos de JOLLY, muchas veces ya a la semana; en la mayor parte de ellos se presentan - o aumentan - glóbulos policromáticos y punteado basófilo, pero por lo general más tarde. No hemos observado a este respecto una diferencia entre cuyes infectados y no infectados.
Ya hemos mencionado que la sangre del cuy B. mostró en su último período el porcentaje enorme de 47 % de reticulocitos. Este hecho concuerda con los datos obtenidos en la bartonellosis de otros animales durante la anemia o en las recidivas.
El valor más alto que encontramos en los cuyes no esplenectomizados, pero con infección natural, fué de 3.7 % (Cuy 9). Sin embargo parece que esta cifra bastante alta no es una indicación de la infección latente, ya que la sangre de los cuyes Nº 1 y 3 (ambos naturalmente infectados) tenía solamente 0.1 % y 1.5 %, respectivamente.
B. Leucocitos.
Su número tenía en los cuyes no operados un mínimo de 3,280 y un máximo de 11.360 por mm. 3.
Estas cifras fueron en 15 cuyes procedentes de Lima (expresado en miles) de 3,16 - 3,20 - 3,80 - 4,00 - 4,04 - 5,08 - 5,24 - 5,32 - 5,40 - 6,08 - 6,40 - 7,00 - 7,80 - 9,16 - 14,64.
Promedio: 6,02 ± 0.74.
En 15 cuyes procedentes de Surco y de Tornamesa estas cifras fueron de: 3,28 - 4,68 - 4,72 - 5,12 - 5,20 - 6,08 - 6,48 - 6,52 - 6,56 - 6.68 - 8,16 - 8,24 - 8,96 - 10,44 - 11,36.
Promedio: 6,83 ± 0,56.
Diferencia del promedio en ambos grupos: 0.81 ± 0.93.
No se puede pues, dar significado a tal diferencia.
Después de la esplenectomía hay un ligero aumento del número de los leucocitos, causado probablemente por una ligera infección secundaria.
En cinco de nuestros animales parasitados este aumento fué seguido por una disminución, precediendo a la presencia de las bartonellas en la sangre. [Semejante fenómeno fué descrito por WEISS Y PONS en la bartonellosis del perro (20) ].
El período en que las bartonellas se encuentran en la sangre está, al contrario, caracterizado por leucocitosis. (Véase cuadro 2). [También este fenómeno se observa tanto en la rata (17) como en el perro (20)]

Los leucocitos del cuy están constituidos en su mayor parte por linfocitos y una pequeña cantidad de granulocitos. En los cuyes procedentes de Lima el porcentaje de los linfocitos varió entre 63 % y 94% en los de la Sierra, entre 59 % y 87 %. Para los granulocitos estas cifras eran respectivamente de 5 a 30 % y de 7 % a 30 %. No pudo comprobarse una diferencia marcada en ambos grupos.
Durante la infección observamos en cuatro cuyes un aumento del porcentaje de los granulocitos neutrófilos. Es decir en los cuyes Nº 1, 5, 10 y E, este porcentaje fué después de la esplenectomía de 8 %, 39%, 9 % y 11 %; durante la infección estas cifras subieron hasta 51%, 71%, 35 % y 42 5%, respectivamente.
[Tal fenómeno es mucho más marcado en la rata (17) y al contrario muy inconstante en el perro (20).]
Eosinofília antes, durante o después de la infección fué descrito por varios autores. SCHWARTZ la encontró en la rata hasta 6% a 8%, pero no en todos los casos. Este fenómeno hemos podido comprobar en algunos de nuestros cuyes. (Véase cuadro 3). Sin embargo se necesita juzgar este fenómeno con cautela, porque, como se sabe, la presencia de vermes puede ser también la causa de eosinofília. Sin duda éste fué el caso en el cuy 3, en las heces del cual encontramos huevos de Tricocéfalos y en las del Nº 6 que contenían huevos de Toxascaris.

Solamente en dos de nuestros cuyes hemos observado un aumento notable de los monocitos. La sangre del cuy Nº 1 contenía 3% de monocitos antes de la esplenectomía; una semana después de la operación estas células no eran visibles pero más tarde reaparecieron y durante la infección espontánea el porcentaje subió hasta 25%,. La sangre del cuy A no presentaba monocitos antes de la esplenectomía y tampoco una semana después; aparecieron estas células 2 semanas después de la inyección con sangre parasitada y aumentaron hasta 11% en el período en que los hematíes infectados ya. eran muy escasos.
[En la rata monocitosis fué observador por BAYON. (17). También en la bartonelosis del hombre este fenómeno es conocido y fué descrito Por MONGE como uno de los primeros síntomas, de regresión, en la fase de defensa, que puede ser precedida algunas veces por eosinofília (13). WEISS lo consideró como fenómeno de remisión sanguínea. (19).]
Los núcleos de los granulocitos en la sangre infectada daban la impresión de tener por lo general más segmentos que los de la sangre no parasitada. (Fig. 11 De vez en cuando encontramos núcleos cuyos segmentos eran colocados en forma de rosetas con 6 hasta 9 pétalos. Semejante fenómeno ha sido descrito por SORINA en la bartonellosis de la rata ( 17). Con el objeto de precisar este fenómeno, contamos los segmentos de 100 granulocitos en la sangre de 8 cuyes normales y en la de 5 cuyes parasitados. Del resultado de estos cálculos, se deriva que el promedio de los segmentos en los cuyes normales fué de 3.46 ± 0.13 y que este valor fué por lo general un poco más grande en la sangre de los cuyes infectados, pero no en todas las muestras. Unas veces se observó un aumento progresivo del número de los segmentos durante la infección. En el cuadro 4 se indican, como ejemplos, los datos sobre la sangre del cuy Nº 1 (naturalmente infectado) y del Nº 5 (experimentalmente infectado).

VI. Anatomía patológica (****)
1. Hígado.
Hemos podido examinar el hígado de los cuyes 3, 6, B, y B; el órgano fué fijado en el líquido de ZENKER e incluido en parafina Los cortes,, fueron coloreados con hematoxilina-eosina. Las, alteraciones encontradas son las siguientes:
A. Alteraciones degenerativas.
a : Focos de necrosis de las células hepáticas que se presentan en el tercio medio o extremo de los lobulillos. (Fig. 12).
b : Degeneración de las células hepáticas, sobre todo granulosa y algunas veces también grasosa.
c : Delgadez de las trabéculas de REMAK; en algunos lugares se han desprendido las células hepáticas, que tienden a aislarse.
d : Se nota también dilatación de las venas, sobre todo de las suprahepáticas; algunas muestran una dilatación máxima y contienen trombos.
B. Alteraciones proliferativas.
Estas tienen lugar en los espacios de KIERNAN; ahí se ven además de leucocitos y linfocitos, una proliferación de tejido conjuntivo. (Fig. 13). Algunas de estas células son macrofagocitarias, habiendo englobado hematíes, ya intactos, ya alterados y conteniendo también pigmento de un color anaranjado, probablemente hemosiderina. En algunos de los animales se ven células de KUPPFER con tal pigmento.
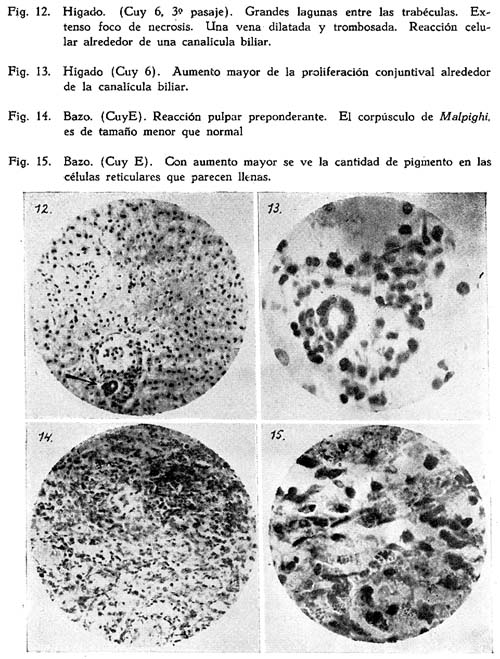
Estas alteraciones son más o menos marcadas según el caso; por ej. en el hígado del cuy B (con anemia grave) domina una proliferación periportal inmensa, mientras los focos de necrosis son modestos. Al contrario, el hígado del cuy 3 (naturalmente infectado) está caracterizado por focos de necrosis extensos. Falta en el órgano del cuy anémico B. las dilataciones de venas tan sobresalientes de los cuyes 3, 6, y E.
Recordamos que focos de necrosis en el hígado han sido descrito en la bartonelosis del hombre (9), del mono (15), del perro (20) y de la rata (11). Sin embargo quizás, más características en el hígado del cuy son las proliferaciones de las células conjuntivas alrededor de los conductos biliares y también los signos de fagocitosis, presentando estas alteraciones un paralelismo con las lesiones del hígado humano. Mencionamos por fin que en ninguno de estos hígados se han observado degeneraciones extensas o gran proliferación de las células de KUPPFER, conocidas en los otros animales. Tampoco se han notado signos de verdadera inflamación.
2. Bazo.
Bazo Cuy 9. (Sacado con la esplenectomía de este animal naturalmente parasitado).
Se ha conservado la parte linfóide, dispuesta como de ordinario en les corpúsculos de MALPIGHI. La parte pulpar muestra figuras de fagocitosis, también hay kariokinesis en algunas de estas células. Las lagunas sanguíneas son muy amplias.
Bazo Cuy E. (Pedacito del órgano, encontrado con la autopsia).
Hiperplasía de la porción pulpar y, en cambio, notable disminución de la parte linfóide. Gran número de los elementos macrófagos está cargado de hemosiderina; se disponen en anchas bandas que se reconocen fácilmente por su pigmento. En torno de las paredes de las arteríolas en los corpúsculos de MALPIGHI hay degeneración hialina. No se nota el manto de linfocitos que envuelve a estos vasos. Sólo unos cuantos elementos aislados se pueden reconocer como tales. (Figs. 14 y 15).
La anatomía del bazo en la bartonelosis de la rata ha sido estudiado por PERLA Y MARMORSTON - GOTTESMAN. En el bazo de ratas jóvenes, infectadas con sangre de animales adultos esplenectomizados, encontraron grandes focos de necrosis y fagocitosis de hematíes por las células de la pulpa ( 11 ). Realizaron también auto-transplantaciones de una parte del bazo en ratas adultas, quitando 7 semanas después el resto de éste órgano; 4 del los 7 animales, con que trabajaron, no mostraron anemia y no fueron visibles bartonellas en la sangre (10). En el autotransplante de las ratas protegidas hallaron una regeneración completa de la porción pulpar, asimismo de los corpúsculos de MALPIGHI; las células de la pulpa contenían gran cantidad de hemosiderina. Al contrarío, en el transplante de la rata no protegida encontraron una destrucción casi total de la parte pulpar, con regeneración de les corpúsculos de MALPIGHI
La estructura del pedacito de bazo del cuy E. - dejado con la esplenectomía incompleta - corresponde, en cuanto a la hiperplasía de la porción pulpar, a la del auto-transplante de las ratas protegidas en los, experimentos de los investigadores citados; este hallazgo parece sostener la hipótesis de dichos autores, que la porción pulpar fuera responsable de la acción protectiva del bazo.
Al concluir esta comunicación, me es grato, manifestar mi agradecimiento al Prof. Dr. TELÉMACO BATTISTINI por haberme permitido realizar este trabajo en el Instituto Nacional de Higiene y Salud Pública de Lima.
Agradezco también a sus Asistentes por la colaboración prestada, ,especialmente a los Dres. V. M. AYULO, A.. HERRER Y J. PONS. M.
Aprecio igualmente la valiosa contribución del Prof. Dr. D. MACKEHENIE y del Dr. J. JIMÉNEZ FRANCO en el estudio anátomo-patológico.
Por fin debo expresar mi gratitud al Dr. E. ENCINAS, quien me permitió obtener algunas microfotografías; presentadas en este trabajo.
* Estación de San Bartolomé del F.F.C.C., lugar de donde Weinman y Pinkerton obtuvieron uno de los cuyes con que trabajaron.
**Determinado con el contómetro.
*** Promedio obtenido a base de 100 glóbulos rojos.
****En colaboración con los Dres. D. Mackehenie y J. Jiménez Franco.
BIBLIOGRAFIA
1. L. ALDANA: Bacteriología de la Enfermedad de Carrión, Lima, 1929. [ Links ]
2. A. HURTADO, J. PONS M. & M. C. MERINO: La anemia de la Enfermedad de Carrión. Lima, 1938. [ Links ]
3. R. JARAMILLO: Rev. de Higiene, t. 20, 1939. [ Links ]
4. J. MARMORSTON-GOTTESMAN D. PERLA: Journal of Experimental Medicine, t. 52, p. 121, 1930. [ Links ]
5. M. MAYER: Arch. f. Schiffs u. Tropenkrankh., t. 25, p. 150, 1921. [ Links ]
6. M. MAYER Comunicación personal. [ Links ]
7. M. MAYER H. ZEISS: Arch. f. Schiffs u. Tropenkrankh.,t. 24, p. 263, 1930. [ Links ]
8. J. NOÉ: Comunicación personal. [ Links ]
9. H. NOGUCHI : Journ. Exp. Med., t. 47, p. 235, 1928. [ Links ]
10. D. PERLA & J. MARMORSTON-GOTTESMAN: Journ. Exp. Med .,t. 52, p. 131, 1930. [ Links ]
11. D. PERLA & J. MARMORSTON-GOTTESMAN: Journ. Exp. Med., t, 53, p. 869, 1931. [ Links ]
12. H. PINKERTON & D. WEINMAN : Proc. Soc. Exp. Biol. and Med., t. 37, p. 596, 1937. [ Links ]
13. R. REBAGLIATI : Verruga Peruana, Lima, 1940. [ Links ]
14. A. RUIZ DEL RÍO: Bol. Soc. Biol, de Concepción, t. 13, p. 47, 1939. [ Links ]
15. P. STRONG ET AL. : Report of first expedition to South America. 1913. [ Links ]
16. E. E. TYZZER & D. WEINMAN : The Amer. Journ. of Hygiene, 30, p. 141, 1939. [ Links ]
17. D. WEINMAN: Les parasites érythrocytaires révélés par la splénectomie, Paris, 1935. [ Links ]
18. D. WEINMAN & H. PINKERTON,: Ann. of Trop. Med. and Paras., t. 32, p, 215, 1938 [ Links ]
19. P. WEISS: Hacia una concepción de la Verruga Peruana, Lima, 1927. [ Links ]
20. P. WEISS & J. PONS M. : Actualidad Médica Peruana, Lima Nº 6, 1938. [ Links ]
21. H. ZINSSER & M. RUIZ CASTAÑEDA Journ. Exp. Med., t. 52, p. 649, 1930. [ Links ]

