










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.3 n.1 Lima mar. 1944
Investigación de portadores de Corynebacterium diphtheriae Oscar Rondón S.1 1 Departamento de Bacteriología e Inmunología del Instituto Nacional de Higiene y Salud Pública. Los conocimientos actuales dan especial interés al papel epidemiológico que juegan los sujetos sanos, que sin saberlo, llevan el germen consigo, distribuyéndolo entre las personas que conviven con él. Y si hoy en día está demostrado que el número de estos portadores no es tan grande como se creyó demostrar en los primeros tiempos -seguramente por que eran insuficientes los conocimientos de los caracteres biológicos del germen- (E. MOSHAGE y J. KOLMER, 1916) no por eso deja de ser, sino el único, el principal factor en la propagación de la difteria. Si bien el grado de inmunidad de una colectividad es el índice importante que señala la receptibilidad a la infección, es principalmente ésta, la infección, quien determina y gobierna la propagación de la enfermedad. En consecuencia, es de interés estudiar los factores que facilitan la difusión del germen y en especial las propiedades y condiciones biológicas que le permiten pasar de huésped a huésped. Es esta razón la que ha influído en nosotros para hacer la revisión de nuestros niños, deseosos de comprobar la existencia de estos portadores, que ya se presumían, y también reconocer las cualidades y tipos de gérmenes que ellos albergaban. Sin embargo, no creemos oportuno hacer la exposición detallada de todos los métodos y materiales que hemos usado con este fin, y nos vamos a circunscribir a informar sobre, los principales métodos y condiciones del trabajo; la presencia de portadores, y los tipos de C. diphtheriae encontrados, de acuerdo con las más recientes adquisiciones. METODOS Y MATERIALES Desde Agosto de 1940 hasta el mismo mes de 1941, se hizo la recolección de muestras de los niños aparentemente sanos que fueron durante este tiempo, probados por nosotros con la reacción de Schick y que ya han sido reseñados en un artículo publicado anteriormente (O, RONDÓN, 1943). Estas muestras fueron tomadas de la faringe y nariz de los niños por medio de hisopos estériles de algodón, los que fueron inmediatamente examinados por coloración de GRAM y LÖEFFLER, a la vez que sistemáticamente sembrados: 1º, en el medio tan comunmente usado de suero-coagulado de LÖEFFLER y después, en una placa Petri con agar-sangre-telurito, preparado de acuerdo a las indicaciones de ANDERSON y col. (1931) fórmula que indicaremos más adelante. Este medio fué escogido para el trabajo de rutina por ser reconocido en la literatura (COOPER y col., 1940; KERRIN y GAZE, 1937), como probablemente superior a los otros medios similares, debido a que da diferencias morfológicas más netas en las colonias, y así hace más fácil la distinción entre los tres tipos de C. diphtheriae, a la vez que permite reconocer a éste, dentro de un gran número de colonias de difteroides, tan frecuentes en los cultivos de muestras de portadores. Nosotros constatamos más tarde también, la veracidad de estas afirmaciones al probar otros medios tales como: el de HORGAN y MARSHALL (1932), el de NEILL (1937), el de WELSCH y THIBAULT (1939), y otros, que si bien algunos son más fáciles de preparar como el de HORGAN y MARSHALL (el cual sólo requiere un agar nutritivo ordinario al que se le agrega 10 cc. de una solución de telurito de potasio al 2% y 10 cc. de sangre de buey o de otro animal), no dan colonias tan características o suficiente desarrollo. No nos fué posible usar los tan recomendados medios de CLAUBERG (GROOTEN, 1940) debido a la dificultad de conseguir los ingredientes necesarios para su preparación. Por lo demás, ninguno de los medios probados dieron resultados mayormente satisfactorios, a tal punto, que nos obligara a substituir el medio de ANDERSON y col., con perjuicio en la uniformidad de la técnica para el aislamiento de los gérmenes. El uso del medio de LÖEFFLER, conjuntamente con el de ANDERSON no fué planeado con el fin de estudiar la superioridad de uno u otro, sino simplemente para aumentar el chance en el diagnóstico, tanto más que algunos bacteriólogos, y entre ellos los mismos ANDERSON y col., han reconocido la dificultad del desarrollo del tipo "mitis" en este medio de telurito. Sin embargo, nosotros no hemos encontrado resultados positivos de desarrollo de C. diphtheriae en el medio de LÖEFFLER que no hayan dado desarrollo en el medio de agar-sangre-telurito, de los investigadores de LEEDS. La rutina seguida por nosotros en el aislamiento de gérmenes de las muestras nasales o faríngeas de los niños fué el siguiente: Después de la incubación a 37ºC. por 24 a 48 horas, los cultivos fueron examinados macroscópicamente para hacer la diferenciación de colonias, y éstas fueron reconocidas microscópicamente por la coloración de GRAM. Los gérmenes que se mostraron Gram positivos y de morfología correspondiente a cualquiera de las distintas especies del género Corynebacterium, fueron resembradas en el medio de suero-coagulado de LÖEFFLER, para completar su aislamiento, y esto fué llevado hasta quedar seguros de su completa pureza, ya que según nuestra experiencia reclama particular atención, mayor que la que comunmente se presta para el aislamiento de otras especies bacterianas. El objeto seguido al aislar toda clase de gérmenes de aspecto morfológicamente parecido al C. diphtheriae, fue para ponernos a salvo del posible error de diagnóstico al eliminar como difteroide a algún C. diphtheriae de tipo "gravis" que pudiera no haber mostrado su característico aspecto de colonia en el medio de telurito, ya que este germen se muestra microscópicamente como un difteroide cuando es cultivado en este medio. Esto nos ha permitido a la vez estudiar el número de niños que albergan en su garganta el C., hofmanii. Por otro lado, en el caso de ausencia de desarrollo, tanto en el medio de agar-sangre-telurito como en el medio de suero-coagulado de LÖEFFLER, a la vez que, un resultado negativo en el examen microscópico de la muestra, no hemos hecho mayores exámenes y se ha catalogado al niño como no portador. El estudio y reconocimiento de las cepas así aisladas, fué llevado a término por lotes, de un número compatible con nuestros materiales, y probados frente a unas cuantas pruebas de identificación. Las semillas que no estuvieron en trabajo, fueron conservadas en tubos de medio de LÖEFFLER sellados, a 4ºC. y resembrados de tiempo en tiempo. Todo esto motivó, que el estudio de algunas cepas no se completara sino después de un año de aisladas y aún, en algunos casos, después de mayor tiempo; sin embargo, al repetir algunas pruebas no se encontraron cambios en su comportamiento, mayores que los ya señalados en la literatura (BARRATT, 1924; FROBISHER, 1938 a), en especial referencia a la facultad fermentativa de los hidratos de carbono, que por lo que parece, no se presentan por cambios en las propiedades biológicas del germen; sino como resultado de las alteraciones no controlables en la composición de los medios de cultivo. En cuanto a la virulencia del germen, ésta se ha mostrado positiva hasta pasados dos años de la fecha del aislamiento. El diagnóstico de reconocimiento del C. diphtheriae fué hecho siguiendo las líneas generales de clasificación indicadas por BERGEY (1939), y las de otros autores dadas en informes que anotaremos oportunamente. Las condiciones excepcionales en que se coloca el bacteriólogo, cuando trata de hacer el diagnóstico de la presencia de gérmenes patógenos en personas que no dan signos clínicos de enfermedad, lo obliga a realizar su trabajo con técnicas más completas y cuidadosas que las usadas para el diagnóstico en sujetos clínicamente reconocidos. Desde luego que este mayor cuidado y más completa técnica, no puede llevarse hasta el extremo de estudiar todas y cada una de las propiedades biológicas y características culturales del germen. A este respecto hemos escogido las propiedades más importantes, y a nuestro criterio suficientes, para estar seguros de que el germen en cuestión, pertenece a la especie en que se le incluye. Estas propiedades pueden ser compendiadas dentro de cuatro modalidades especiales, que son: Primero el aspecto cultural en la placa de agar-sangre-telurito de ANDERSON y col.; Segundo las características morfológicas y tintoriales del germen; Tercero las propiedades biológicas más demostrativas y generalmente aceptadas, en especial la acción fermentativa sobre algunos hidratos de carbono, tales como, la glucosa, maltosa y sacarosa; y Cuarto la evidencia de su poder patógeno por demostración de su virulencia. 1º COMPORTAMIENTO DEL C. DIPHTHERIAE EN EL MEDIO DE ANDERSON Y COL. Medio de Cultivo. El medio de ANDERSON y Col. fue preparado siguiendo lo más estrictamente posible las indicaciones de sus autores, con algunas variantes, las que nos obligan a detallar su preparación. Preparación del Medio. a) A 1.000 cc. de agua de pozo, calentada a 48ºC., se le agregó 1 kilo de carne de res, previamente pesada, y se mantuvo esta temperatura por una hora. El jugo fué extraído por compresión a través de un lino, después fué dejado toda la noche en frío, filtrando a través de papel de filtro. b) A 1.000 cc. del filtrado, se agregó 20 grms. de peptona (P. D. y Cía.) y 5 grms. de cloruro de sodio, calentándose a no más de 45|C. para su completa disolución. c) La reacción de alcalinidad se ajustó a un pH de 7.6, tomando una pequeña parte de la mezcla anterior y calentándola a 90°C. por 15 minutos, con el objeto de conseguir la clarificación del medio y así hacer posible su titulación por el rojo de fenol. La cantidad de seda N/1 necesaria para dar el pH de 7.6 a 10 cc. de esta parte clarificada, indica la cantidad necesaria de soda que debe añadirse a la totalidad del medio. d) La esterilización se hizo por filtración a través de una bujía de Berkefeld, previo pasaje por papilla de papel, en reemplazo del filtro Seitz K que es indicado en la técnica original. El caldo así preparado fué repartido en frascos de 100 cc. cada uno, y guardados en frío, después de constatar su esterilidad. e) La preparación final se hizo en la siguiente forma: Se mezcló en partes iguales el caldo con un agar al 5% en agua, previamente esterilizado a 120°C. por 30 minutos, a esta mezcla mantenida a 50°C., se le agregó de 7 a 10 % de sangre desfibrinada de conejo, y 0.04% de telurito de potasio. Antes de ser repartido en placas Petri, fué calentado a 75°C. por 15 minutos en B. M. Aspecto de las Colonias. El reconocimiento de la presencia de desarrollo de C. diphtheriae en este medio, sin entrar en la diferenciación de tipos descritos para esta misma especie, se hizo por el desarrollo precoz y abundante en las primeras 24 horas, así como por la coloración negra o gris-negruzca y mayor tamaño de las colonias, en comparación con la de los difteroides. La observación más tardía rectificó nuestro primer criterio. Es importante hacer notar que este reconocimiento de colonias se ve perturbado cuando la siembra es demasiado abundante y las colonias están muy juntas. 2° CARACTERES MORFOLOGICOS Y TINTORIALES De cultivos jóvenes en medio de suero-coagulado de LÖEFFLER, se prepararon los frotis para estas observaciones. El germen fué reconocido por su morfología y agrupación característica, sirviéndonos en especial su aspecto bacilar, curvo, y abultado en uno o en ambos extremos; así como, por su coloración irregular con el azul alcalino de LÖEFFLER, y su comportamiento al Gram. La demostración de la presencia de corpúsculos metacromáticos, fué hecha por uno de los métodos de coloración de NEISSER, modificado en la siguiente forma (CARVALHO, 1934): Tres soluciones son necesarias:
| A | Azul de metileno............. | 1 grm. |
| Alcohol absoluto............. | 20 c.c. | |
| Accido acétido q.p.......... | 50 c.c. | |
| Agua destilada............... | 1000 c.c. | |
| B | Cristal violeta................. | 1 grm. |
| Alcohol absoluto............. | 10 c.c. | |
| Agua destilada............... | 300 c.c. | |
| C | Vesuvina...................... | 1 grm. |
| Agua destilada............... | 300 c.c. |
El método de coloración es el siguiente:
Se colorea por 5 a 10 minutos con la mezcla recientemente preparada de 2 partes de la solución A y una parte de la solución B. Después de un lavado ligero con agua, se hace actuar la solución C por uno a 2 minutos. Los tiempos señalados aquí, son los que la experiencia nos ha mostrado dar mejores resultados.
Todos los gérmenes reconocidos como C. diphtheriae mostraron granulaciones polares con esta técnica.
3° LAS CARACTERISTICAS CULTURALES Y BIOLOGICAS DEL GERMEN
No intentamos describir todos los caracteres culturales por los que fueron probados los gérmenes, basta sólo decir que se mostraron de acuerdo con lo conocido en lo referente al comportamiento del desarrollo en agar, caldo, gelatina, leche tornasolada, suero-coagulado, etc.
De las propiedades biológicas estudiadas son fundamentalmente importantes, las correspondientes a la acción fermentativa de la glucosa, maltosa y sacarosa (FROBISHER, 1938 a). El medio usado con este fin, fué el de Hiss (BARRATT, 1924) suero-agua conteniendo 1% de carbohidrato con tornasol como indicador.
Después de la filtración preliminar del suero de ternera, la mezcla de una parte de suero y dos de agua destilada, fue calentada durante 30 minutos a 100°C., antes de añadirse el carbohidrato, con el fin de destruir la acción hidrolizante de la sangre. Cuando el carbohidrato y el indicador fueron añadidos, los tubos fueron calentados durante 15 minutos a 100°C., por tres días sucesivos.
Para probar la esterilidad, el lote completo fué incubado por 3 días a 37°C., ésto último demostró ser indispensable para obtener resultados uniformes. En casos dudosos fué demostrada la pureza de la semilla y nuevamente probadas bien en el mismo medio de HISS, o en otro de los usados por ANDERSON y col. (1931), FROBISHER JR. (1938), o por SCHROEDER (1930), quedando en esta forma definitivamente establecida la acción sobre estos tres azúcares.
El tiempo de observación de estos medios hidrocarbonados, para sorprender algún cambio tardío, fué hecho durante 7 días, y aún en algunos casos, hasta durante 2 semanas. Aunque se observaron ligeras variaciones en el grado de fermentación, en especial con la maltosa, éstas no eran tales como para llevarnos a falsas conclusiones.
Por los resultados alentadores encontrados por PISO ITALO (1939) en la acción del C. diphtheriae sobre medios líquidos conteniendo urea, para diferenciarlo de los difteroides, nosotros hemos probado todas las cepas en este medio.
La prueba ha sido realizada en un caldo ordinario preparado con peptona 10 grms. (DIFCO). Cloruro de Sodio 5 grms., infusión de carne 1.000 cc. esterilizado por autoclave a 120°C. por 30 minutos. Se completa la preparación agregándole 2 % de Urea, previamente disuelta en agua destilada y esterilizada a 100°C. Después de la incubación por 24 a 48 horas, se le agrega 1 ó 2 gotas de fenolftaleína, en solución alcohólica al 1 %.
Es interesante tener presente algunas precauciones recomendadas por los autores:
A) El caldo debe tener un pH. de 7.5 o sea por debajo del límite de viraje de la fenolftaleína (pH. 8.5), que sirve para dar un margen de seguridad a la posible formación de trazas de amoniaco por los C. diphtheriae, y en esta forma hacer mucho más seguros los resultados.
B) El medio debe de prepararse agregando a 9 cc. de caldo, 1 cc. de solución de Urea al 20 %, en condiciones de esterilidad, y con el control respectivo.
C) Por último, la solución de Urea debe ser esterilizada a no mayor temperatura de 100°C., porque se corre el riesgo de descomponer la Urea con producción de amoniaco, si se le esteriliza a temperaturas más altas. Esta solución se mantiene indefinidamente sin alterarse, siempre que no pierda su esterilidad.
Siguiendo este método no hemos encontrado ningún C. diphtheriae que de alcalinidad dentro de las 24 ó 48 horas de incubación, de acuerdo con lo señalado por el autor italiano. No podemos decir lo mismo para los resultados obtenidos con los difteroides, ya que en contra de lo afirmado por Piso I., hemos encontrado difteroides que tampoco alteran la Urea; sin embargo, creemos a este método útil para el reconocimiento del C. diphtheriae, ya que para éste, es una reacción sino propia, bien manifiesta y constante.
4° DETERMINACION DE LA VIRULENCIA
Las condiciones morfológicas, culturales y de coloración de los C. diphtheriae, deben de ser consideradas de valor relativo como medios de identificación del germen, por no estar definitivamente sancionados los métodos de prueba. En consecuencia, es peligroso fiarse únicamente de estos caracteres en la diferenciación de los C. diphtheriae de los bacilos C. difteroides y siempre es necesario, estudiar el poder patógeno experimental cuando el origen de los gérmenes es de sujetos aparentemente sanos.
Este concepto es generalmente aceptado por la mayoría de los bacteriólogos (ANDREWES y col. 1923; TOPLEY y WILSON, 1942; CARRIEU y col. 1937; WELSCH y THIBAULT, 1939 a). Nosotros hemos probado el poder patógeno experimental, ajustándonos al método intracutáneo que es el usado por ANDERSON y col. (1931 ). Este método ha probado dar resultados correctos tan iguales que el subcutáneo (ANDREWES y col. 1923).
Revisando la literatura más reciente acerca de este tema, se encuentra que la técnica intradérmica en el cuy, es la más comunmente usada y con resultados muy satisfactorios, sin embargo, hay quienes como SCHROEDER (1930 a), encuentran que el método subcutáneo da resultados positivos en proporción algo mayor que el intradérmico. La economía en animales, y por esto, la posibilidad de hacer varias pruebas del mismo germen en distintos animales, en otra ventaja del método intradérmico.
La técnica adoptada por nosotros, difiere de la usada por ANDERSON y col., lo mismo que de la más común de EAGLETON y BAXTER, descrita detalladamente por OKELL y PARISH (1926), y otras actualmente en desuso.
En general nosotros hemos seguido la técnica de W. MAIR (1930) usada por este autor en 1612 cepas de C. diphtheriae, que resumiremos en la siguiente forma.
Prueba Intradérmica. Se preparan suspensiones de cultivos jóvenes y puros, en medio de LÖEFFLER, aproximadamente standardizados por opacidad, que contengan 100 millones de bacilos por cc., y de éstas, 0.2 cc. (20 millones) son inyectados intradérmicamente en dos cuyes que llamaremos desde ahora, de "Prueba" y de "Control".
La dosis de suero antitóxico, suficiente para evitar la muerte del cuy de "Prueba" sin impedir la lesión cutánea, es de 1/50 de unidad de antitoxina por gramos de peso del animal, la que es administrada subcutáneamente, e inmediatamente después de la inyección de los gérmenes.
El cuy usado como "Control" recibe con 24 horas de anticipación al día de la prueba, 700 unidades antitóxicas, por vía subcutánea o intraperitoneal, y después es inoculado al mismo tiempo y con igual dosis que el animal de "Prueba".
Si ambos cuyes no muestran signos de lesión cutánea se repite la inoculación, pero sin administrar antitoxina al animal de "Prueba", y subiendo la dosis para ambos cuyes, a 100 millones de gérmenes contenidos en los 0.2 cc. de inyección. Si aún no muestran lesiones cutáneas, se considera al germen como "no virulento".
El autor señala la posibilidad de fallar el suero en su acción preventiva, para algunos animales; pero este suceso extraordinario, no se repite si se prueba nuevamente en otro animal.
A los resultados de esta prueba han sido a los que les hemos dado mayor importancia en el reconocimiento del C. diphtheriae, y nos ha servido también para distinguir los gérmenes virulentos de los no virulentos, dentro de les clasificados como C. diphtheriae por las otras características.
Los exámenes llevados con el fin de determinar los tipos de C. diphtheriae, según lo encontrado por ANDERSON y col. serán estudiados con especial amplitud más adelante.
Los gérmenes que no pudieron ser identificados como pertenecientes a la especie C. diphtheriae, fueron reconocidos e identificados, en su mayoría, como pertenecientes al C. hofmanii. Para esto seguiremos las mismas líneas generales que las recorridas para el C. diphtheriae y que resumiremos a continuación:
1° Aspecto de la Colonia y otras características del C. hofmanii en el Medio de Agar-sangre telurito de Anderson y col.
El reconocimiento del C. hofmanii es más posible en este medio que cuando se le siembra en otros similares. Sin embargo, casi todos los autores no le prestan mayor atención y algunos como PETERSON (1939), KERRIN y GAZE (1937) y NEILL (1937), ni siquiera mencionan esta posibilidad. Seguramente algunos de los muchos medios con telurito, no permiten esta diferenciación, como lo hacen notar TEMPÉ (1930) y la investigadora GROOTEN (1940), para uno de los medios con telurito de CLAUBERG.
Nosotros pensamos que el reconocimiento de la colonia del C. diphtheriae en estos medios, implica necesariamente la posibilidad del reconocimiento deja colonia del C. hofmanii, ya que ambos desarrollan sin impedimento en cualquiera de estos medios.
Por lo demás, de acuerdo con las afirmaciones de ANDERSON y col. y también con las de ALLISON y AYLING (1939), para el medio ideado por ellos, creemos posible distinguir estas colonias, si se tiene alguna experiencia en el uso del medio.
Los caracteres más importantes para esta diferenciación son principalmente, el desarrollo tardío, a veces después de pasadas las primeras 24 horas de incubación, esto parece ser debido, como lo ha demostrado CORRADO AJO (1935), no a la presencia del telurito de potasio, sino a una propiedad del plasma (suero citratado u oxalatado).
La morfología de la colonia, que es circular, lisa y convexa, la hace muy parecida al tipo "mitis" del C. diphtheriae, con la diferencia de que aquella es de color blanco perla ligeramente gris, en contraste con la colonia de tipo "mitis" que es negra. Más tarde la parte central de la colonia toma un color gris negro, quedando un margen opaco de color blanco grisáceo. En lo referente a este tono gris-negruzco, que adquiere la colonia en su parte central, hace inseguro su reconocimiento;
pero esto es posible salvar, si se tiene cuidado de que las colonias se encuentren bien separadas una de otra, con este fin la siembra no debe de ser abundante.
Esto y la presencia, en algunas muestras, de gérmenes que no corresponden a las dos especies que estamos estudiando, hacen a veces muy difícil el reconocimiento.
2° Caracteres Morfológicos y Tintoriales. Los gérmenes de esta especie fueron reconocidos por su morfología bacilar, regularmente uniforme, de tamaño corto y ligeramente afilado en sus extremos. De coloración uniforme con el azul de LÖEFFLER, interrumpido sólo por un septum transverso, mediano, sin colorear. Generalmente ausente de gránulos polares, por la coloración de NEISSER ya indicada. Fuertemente Gram positivo.
Todos estos caracteres fueron estudiados de un cultivo joven en medio de LÖEFFLER, ya que sabemos, como hemos dicho anteriormente, que el tipo "gravis" en el medio chocolate-telurito de ANDERSON y col., se muestra parecido a los gérmenes pertenecientes a esta otra especie.
3° Caracteres Culturales y Biológicos. Los caracteres culturales y biológicos de estos gérmenes fueron probados sustancialmente, en lo que se refiere a su propiedad negativa para atacar cualquiera de los tres azúcares principales, (ANDREWES y col., 1923; BARRATT, 1924) glucosa, maltosa y sacarosa, conjuntamente realizados con las pruebas del C. diphtheriae en el mismo medio de Hiss, suero-agua tornasolada.
Hemos tenido especial cuidado de comprobar las condiciones de esterilidad del cultivo y el desarrollo abundante en este medio. A las pruebas conseguidas por este método les hemos dado mayor valor para el reconocimiento de esta especie.
La prueba de la acción metabólica sobre la Urea, en el medio indicado por ITALO PISO (1939), fué inconstante para muchos de ellos; aún prolongando el tiempo de incubación hasta las 48 horas. Esto nos impidió incluir en esta especie un gran número de gérmenes, que al ser probados con posterioridad para otros caracteres generales, demostraron ser pertenecientes a esta misma especie. Lo que, como vemos, está en contra de lo afirmado por ITALO PISO y resta mucho valor a esta prueba.
4° Prueba de la Virulencia. La constatación del poder patógeno experimental por determinación de la virulencia en el cuy, no fué determinada por ninguno de estos gérmenes, que mostraron todos los caracteres típicos de la especie C. hofmanii, y que en consecuencia la prueba de la virulencia era superflua para su identificación, ya que era de todos conocido que no tienen rol patógeno.
Además de probar el comportamiento frente a estas cuatro características de identificación, todos los tipos fueron probados en sus propiedades generales sobre los medios comunes, tales como: agar, caldo, gelatina, leche tornasolada, suero coagulado de LÖEFFLER, etc. De acuerdo con esto, nosotros identificamos de primera intención, un gran número de cepas, que hubiera sido mayor, sin la interferencia de los resultados equívocos en el medio de urea.
Con todo, un número pequeño de cepas no fué posible incorporarlas, ni dentro de la especie C. diphtheriae, ni en la especie C. pseudodiphthericum o C. hofmanii, quedando agrupadas como especies "no identificadas". Es posible que muchas de ellas puedan ser incluídas dentro de uno de los once grupos reconocidos por la autora BARRATT (1924) pertenecientes al género Corynebacterium; pero sin mayor ventaja para su interpretación desde el punto de vista con que planeamos nuestro trabajo.
La mayoría de estos gérmenes fueron fermentadores de sacarosa y glucosa, fallando en esta acción sobre la maltosa, o bien en su comportamiento frente a otros medios o reacciones, por lo cual no fué posible considerarlas como C. xerosis.
RESULTADOS OBTENIDOS
En el cuadro I resumimos los resultados obtenidos en el examen bacteriológico de 791 muestras nasales y faríngeas de niños sanos, tomadas por medio de hisopos estériles de algodón, y examinadas por cultivo en el medio de ANDERSON y col. a la vez que en el medio de suero coagulado de LÖEFFLER.
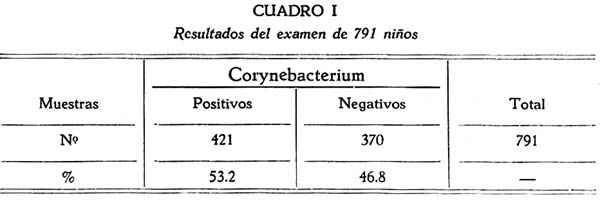
De estas 791 muestras, fué posible aislar 421 cepas de gérmenes que podían corresponder a cualquier especie del género Corynebacterium, o sea que el 53.2 % de estos niños, resultaron ser portadores de alguno de ellos, mientras que 370 (46.8 %) se mostraron libres de esta infección.
Después del estudio bacteriológico para el reconocimiento de las especies a que pertenecían las 421 cepas aisladas, encontramos que 332 pertenecían a la especie C. hofmanii, o sea que 41.9 % de los niños eran portadores de este germen.
En los estudios preliminares sobre las cuatro bases de identificación del C. hofmanii, fué posible reconocer solamente 225 gérmenes de esta especie, que reunían todas las condiciones para ser identificados como tales; quedando un grupo de 117 cepas, que siendo C. hofmanii, fallaron en presentar un o más de los caracteres principales estudiados especialmente en su comportamiento en el medio de urea de ITALO PISO, y en parte a otros caracteres, como morfología y presencia de corpúsculos polares; colonias no características en el medio de ANDERSON y col. y por último, alguna acción fermentativa anormal en el medio de HISS; pero con estudios más completos y repetidos se concluyó catalogándolos siempre, como pertenecientes a esta especie, despreciando el comportamiento en el medio de urea, que como ya hemos dicho, no nos dió los resultados esperados.
En el cuadro II, nosotros mostramos los resultados del estudio bacteriológico de 421 cepas, aisladas de 791 niños y catalogadas de acuerdo a la identificación que fué posible realizar.
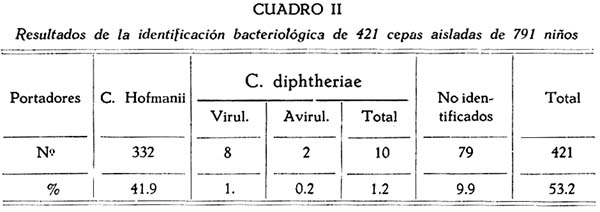
En él señalamos que solamente 10 gérmenes cayeron en el grupo de la especie Corynebacterium diphtheriae. De éstos, 8 se mostraron patógenos para el cuy por la prueba de virulencia y 2 fueron negativos para ésta; o sea, que el 1% de niños, eran portadores de gérmenes virulentos.
Los otros gérmenes que consignamos en el Cuadro II con el nombre de "no identificados", forman un grupo heterogéneo constituido en su mayoría, por cepas que no fueron completamente identificadas, debido a que las perdimos por muerte de la semilla, en tiempos variables. Es importante indicar, que ellas no mostraron características de C. diphtheriae, hasta donde llegó su reconocimiento antes de su muerte o contaminación.
Un total de 11 cepas, incluídas en este grupo de "no identificadas", a las cuales se les hizo un estudio completo, merecen especial mención; ya que algunos, en especial 2 de ellos, dieron caracteres muy similares a C. diphtheriae, fallando en atacar la maltosa, y aún solo débilmente, la glucosa. Los demás caracteres, como colonia en el medio de ANDERSON y col., forma y granulación polares, lo mismo que su comportamiento en el caldo urea, fueron similares a los C. diphtheriae, y nosotros los hubiéramos incluido entre ellos, si hubieran producido lesión en la piel del cuy con la prueba de virulencia.
Los otros 9 gérmenes de estos 11, fueron en su mayoría fermentadores de la sacarosa, pero, no identificados como C. xerosis por fallar, bien la acción sobre la maltosa, o bien en sus características morfológicas frente al colorante de azul de LÖEFFLER o al de NEISSER. Además los que no fermentaron la sacarosa, atacaron la manita.
Por todo esto no fué posible catalogarlos dentro de alguna de las especies de Corynebacterium bien definidas; pero fué suficiente para distinguirlas de las especies C. diphtheriae o C. hofmanii, que era nuestro principal objetivo.
TIPOS DE C. DIPHTHERIAE ENCONTRADOS
Creemos indispensable completar el estudio de los C. diphtheriae encontrados, de acuerdo con las nuevas adquisiciones de ANDERSON y colaboradores. (1931, 1933).
Si el reconocimiento de la virulencia en gérmenes aislados, de niños sanos, es fundamental en el estudio de la difusión de la difteria, no menos importante es hoy día el reconocimiento del tipo "gravis" en estos mismos portadores, ya que los investigadores que lo reconocieron, lo mismo que muchos otros que han confirmado estos trabajos, especialmente en Inglaterra, Estados Unidos y Alemania, lo asocian a procesos clínicos más graves, que no ceden a la terapéutica usual de la antitoxina y lo más importante aún, constatan que en las comunidades donde hay un aumento en el índice de morbilidad y mortalidad para la difteria, hay también el predominio de tipo "gravis" sobre el "mitis", y como dice DUDLEY y col. (1934), el "gravis" reemplaza al "mitis" en los portadores, cuando se presenta un aumento de la morbilidad. (FROBISHER JR. 1942).
Por tanto, es de suma importancia estudiar el predominio de alguno de ellos en nuestra colectividad.
El reconocimiento de estos tipos de C. diphtheriae está realizado a base de las indicaciones y características dadas por ANDERSON y col. (1931 y 1933), que resumiéndolas son:
1° El aspecto de la colonia en el medio de telurito-chocolate en estos mismos autores, cuya preparación ya hemos indicado anteriormente.
Ellos afirman que el tipo "gravis" desarrolla con colonias de color gris o gris-negruzco, grandes (3 mm.) y con una morfología característica por su estriación radiada, en correspondencia a sus bordes dentados, un centro irregular más elevado y obscuro, que según la comparación de los mismos investigadores, en conjunto se parece a una "margarita". Describen otras colonias de este mismo tipo que son conocidas con el nombre de "gravis no típico", en las que las estriaciones radiadas y lo dentado de los bordes es más suave.
El tipo "mitis" da una colonia de color negro, mediana, (1 a 2 mm.) y de forma convexa, superficie y bordes lisos con un angosto margen blanco-grisáceo, más manifiesto cuando se examina a trasluz.
El tipo "intermedius" da en este medio colonias que ellos describen como, de color negro, pequeñas (1 mm.), aplanadas, elevadas en el centro, de color más obscuro y brillante y ligeramente dentada en la periferia.
En nuestra experiencia, en algunas ocasiones se presentan colonias que son difíciles de catalogar, especialmente en lo que se refiere al tipo "mitis".
2° La acción fermentativa sobre el almidón, dextrina y glicógeno. Los autores recomiendan un medio especial formado por suero-agua con 1% de peptona y rojo de fenol como indicador. Nosotros hemos usado este medio y también el de HISS suero-agua, o más simplemente el recomendado por FROBISHER JR. (1938) constituído por caldo ordinario, libre de carbohidratos y con púrpura de bromo-cresol como indicador.
Los hidratos de carbono fueron añadidos estérilmente a los tubos, en soluciones acuosas y esterilizadas al calor. La concentración de fécula es de 0.5 % y de dextrina 1 %. Sin embargo, nosotros hemos encontrado resultados más uniformes para la fécula, usando agar-suero con 0.5% de fécula, e incubadas cuatro días a 37°C., en el que se encuentra un halo definido alrededor de las colonias "gravis", cuando las placas son tratadas con el reactivo de LUGOL.
El tipo "gravis" es reconocido por su acción hidrolizante definida sobre la fécula, glicógeno y dextrina.
El tipo "mitis" y el "intermedius" no atacan la fécula ni el glicógeno, y son variables con la dextrina.
3° El aspecto de desarrollo en caldo. Para este estudio, nosotros hemos usado el caldo indicado por FROBISHER JR. (1938) libre de carbohidratos, ya que la presencia de éstos acidifica el caldo por la acción del germen, y esto dificulta la formación de película.
El tipo "gravis" es señalado como formador constante de película en la superficie, y desarrolla en grumos gruesos que se adhieren a la pared del tubo o caen al fondo.
El tipo "mitis" por el contrario, no forma película y desarrolla en grumos finos enturbiando el caldo.
Por último, el "intermedius", sin formar películas, se parece mucho al desarrollo del "gravis" en este medio.
4° Cambio rápido del pH. en el caldo. Usando el mismo medio libre de carbohidratos, se constata que: el tipo "gravis" da una acidificación fugaz del medio, el que se hace alcalino después del segundo día; hecho que no es constatado en los tubos en que desarrollan los tipos "mitis" o "intermedius".
5° El poder hemolítico señalado ya desde antiguo (ANDREWES y col., 1923), para algunas especies del C. diphtheriae, ha sido reconocido por ANDERSON y col., como un buen método de identificación del tipo "mitis", si se siguen las indicaciones de HAMMERSCHMIDT, las cuales se deben tener en cuenta si se quiere obtener resultados satisfactorios, debido a la gran variabilidad de esta prueba, sobre todo en relación al tipo de sangre usada, como ha sido señalado por GOLDIE (1933).
Nosotros nos hemos valido, para el estudio del germen, del mismo caldo corriente libre de carbohidratos, ya indicado. Los glóbulos rojos han sido humanos y emulsionados al 2 % después de ser lavados.
La prueba fué llevada a cabo, de acuerdo a la manera descrita por ANDERSON y col. (método de HAMMERSCHMIDT); así, a 1 cc. de cultivo del germen en caldo, de 48 horas, se mezcló 1 cc. de la emulsión de glóbulos rojos humanos, incubando la mezcla por dos horas a 37°C. y haciendo la lectura después del reposo de toda la noche en la nevera.
Según lo indicado, el tipo "mitis" es el único que tiene acción hemolítica, fallando los tipos "gravis" e "intermedius" para producirla.
6° Las diferencias morfológicas y de coloración también han sido indicadas como útiles para reconocer estos tipos: el "gravis" se muestra en el medio de telurito, con una morfología "parecida a la de un difteroide muy corto" (según la expresión de ANDERSON y col.), sus granulaciones polares son pequeñas y escasas. Todo esto en contraposición con el tipo "mitis", que en este medio tiene la morfología clásicamente descrita, con gran número de granulaciones polares y éstas de gran tamaño. El tipo "intermedius", se muestra con la morfología propia de la especie con granulaciones también relativamente abundantes y de buen tamaño; sin embargo, se afirma que su coloración es la más desuniforme de todas ellas (TOPLEY y WILSON, 1942).
Creemos necesario hacer presente que a nuestro criterio, las pruebas de hemolisis, reversión del pH, y formación de película, son las pruebas de lectura más nítida.
El resultado obtenido al probar para estas seis características, todas nuestras cepas "de C. diphtheriae aisladas de sujetos sanos, es el siguiente:
Solamente una cepa se mostró conforme con todos los caracteres propios del tipo "gravis" y aún ésta, con una morfología en el medio de telurito-chocolate no del todo característica, pues fué relativamente grande, aunque sin la morfología propia y descrita para el tipo "mitis" o "intermedius". Otras dos cepas tipo "gravis" fallaron en su comportamiento frente a la dextrina, la que no fué atacada, así como también, una de éstas dió colonias en el medio de telurito-chocolate de tipo intermedio, y no formó película, resistente en caldo. Esta última cepa, de las tres enunciadas, fué la única virulenta para el cuy, mientras que las otras dos se mostraron indiferentes.
El grupo de cepas con todos los caracteres del tipo "mitis" fué más numeroso, ya que cinco de ellas fueron reconocidas con todos los caracteres propios del tipo y fueron virulentas.
Dos cepas se mostraron francamente intermedias entre "gravis" y "mitis", por el comportamiento frente a estas características, sin embargo una de ellas se mostró más próxima al tipo "gravis" por atacar la fécula, no tener poder hemolítico y dar ligera película en caldo. Por lo demás, no hemos encontrado un germen con las características señaladas por ANDERSON para el tipo "intermedius"; pero si, la mayoría de ellas exceptuando la típicamente "gravis" y las cinco típicamente "mitis", han mostrado poseer signos intermedios entre "gravis" y "mitis".
En resumen, sólo una cepa se mostró casi típicamente "gravis", más otra que sólo falló en el carácter de fermentación de la dextrina. Ambas fueron no virulentas. Cinco cepas se mostraron típicamente "mítis" en todos sus carácteres y fueron todas virulentas. El resto mostró características intermedias entre "gravis" y "mitis", acercándose más a uno u otro tipo. Todas estas también fueron virulentas.
La totalidad de estos gérmenes pertenecientes al C. diphtheriae, fueron aislados de niños sanos, comprendidos en las edades límites de 4 años como mínimo y 15 años como máximo. Todos estos niños dieron reacción de Schick negativa, incluyendo hasta los que fueron portadores de gérmenes avirulentos, desde luego que las muestras fueron tomadas indistintamente de niños Schick positivos o negativos, así como también, sin distinción entre edades, sexo o raza. Cabe informar que gran número de muestras fueron tomadas de niños menores de la edad de 4 años, límite bajo el cual no hemos encontrado portadores.
DISCUSION
El estudio de "portadores" de bacilos diftéricos, está lleno de serias dificultades, que nacen tanto de la falta de técnicas precisas y fáciles de aplicar en el reconocimiento del germen, como seguramente de la naturaleza misma de estos organismos.
Así, es frecuente encontrar cambios en sus respuestas culturales, aún teniendo precaución de mantener condiciones uniformes en el trabajo; y uno, rara vez está seguro de que la respuesta de un solo cultivo individual, pueda revelar exactamente las características del germen, obligándonos a repetir las pruebas y a veces, a multiplicar las técnicas. Sin embargo, hay algunas pruebas que son más claras y constantes en sus resultados, pero durante el reconocimiento de las especies o tipos, hemos podido constatar que ningún carácter solo o grupo menor de los que hemos usado, es digno de nuestra confianza como criterio único para la clasificación de las especies, y esto es mucho más manifiesto al hacer el reconocimiento de los tipos de C. diphtheriae con las seis características de ANDERSON y colaboradores.
La necesidad de establecer métodos simples y mejores para la identificación del bacilo diftérico, se hace sentir más cuando tratamos de hacer un estudio comparativo de los resultados obtenidos en la búsqueda de portadores de un lugar con otro o de un tiempo a esta parte. Además, la falta también de un método y criterio único en el reconocimiento de portadores, hace estériles la mayoría de las comunicaciones sobre este tema para ser utilizadas comparativamente. Es frecuente encontrar en la literatura estudios sobre portadores en comunidades numerosas, reconociendo el germen solamente por el examen microscópico después del cultivo en suero-coagulado de LÖEFFLER, seguramente porque estos estudios son seguidos con el deseo del reconocimiento rápido de portadores para ser retirados de la colectividad infectada; y es natural, que los porcentajes altos encontrados en estas condiciones, disminuyan fuertemente si son sometidos a un examen más completo.
Así DUDLEY (1923) nos da un ejemplo claro: de 14 gérmenes aislados de portadores en la población en general, y reconocidos morfológicamente como bacilos diftéricos, 4 de éstos fallaron en fermentar la glucosa, y 6 fueron avirulentos al completar su estudio, con lo que el porcentaje inicial de 3.5 % disminuyó a 1 % de portadores verdaderos.
El mismo término de "portador" debe ser aceptado, como reclama ANDREWES y col. (1923), en el sentido de "sujetos que albergan el germen diftérico virulento, sin dar signos de enfermedad", ya que éstos son los verdaderamente importantes en la epidemiología de la difteria. Y aún si estos sujetos probaran ser Schick-negativos, como en nuestro caso, habríamos despejado la posibilidad de confundirlos con sujetos que estén en un período de incubación; tal como lo ha hecho DOPTER (1935), para distinguir las anginas banales en portadores de C. diphtheriae.
En conclusión, el estudio comparativo entre los resultados de un lugar con otro, se hace particularmente difícil, tanto porque los niños probados no se encuentran en condiciones similares en relación al momento epidemiológico en que son examinados; como por no haber uniformidad en el criterio de lo que se entiende por "portador", ni tampoco en las bases del reconocimiento del bacilo diftérico.
Es frecuente encontrar en la literatura porcentajes bien altos de portadores; pero en estos casos, o no se ha hecha el estudio completo de identificación del germen, o se trata de colectividades en plena evolución de una epidemia.
Por lo demás, el 1 % de portadores reconocidos por nosotros, concuerda con algunos porcentajes encontrados en otros lugares, como lo indicado por DUDLEY en Inglaterra, ya señalado anteriormente y en el cual se ha trabajado en condiciones similares a las nuestras, esto es: investigando en niños de la población en general, fuera de la influencia de una epidemia y en las mismas bases de identificación del germen.
En fin, el 1% de portadores virulentos, encontrado por nosotros, tiene valor en sí y para el momento en que fué realizado el examen, ya que está muy lejos de poder indicar el número de personas que pueden haber estado en esta condición de tiempo en tiempo en nuestra colectividad.
Sin embargo, si el estudio comparativo en relación al tiempo y al lugar no es posible, la constatación en sí de los portadores en niños previamente probados con la reacción de Schick, nos permite deducir, de acuerdo con los conocimientos actuales, que:
Al comprobar la existencia de portadores de gérmenes virulentos en niños Schick inmunes, estamos confirmando el criterio de que el germen diftérico puede ser "virus para unos y antígenos para otros", a la vez que la inmunidad diftérica entre nosotros también se establece o se mantiene por la influencia de estas infecciones inaparentes, que son verdaderos estímulos antigénicos naturales, ya que en les niños examinados no había antecedentes de inmunización artificial hasta el momento en que realizamos estos exámenes.
Además que, entre nosotros, este número de niños portadores en proporción de 1 por cada 100 niños sanos, son los causantes de los casos clínicos de difteria que observamos en nuestros hospitales (Memorias, 1933-1938), ya que se está de acuerdo en considerar esta fuente de infección como fundamental en la difusión de la difteria.
Por otro lado, en relación con los tipos de C. diphtheriae reconocidos por las seis características indicadas por ANDERSON, HAPPOLD, MC. LEOD y THOMPSON (1931) y encontrados por nosotros en nuestra población infantil con un predominio del tipo "mitis" sobre el "gravis", y éste, en la mayoría de los casos sin virulencia, queremos ver un factor mas que explicaría la no existencia entre nosotros de esas temidas invasiones epidérmicas que existen o existieron en otros países; ya que después de los estudios de ANDERSON y col. parece confirmado por muchos otros investigadores (DUDLEY y col., 1934; ANDREWES y col., 1923; TOPLEY y WILSON, 1942), que el predominio del C. diphtheriae "gravis", ,coincide con el aumento del índice de morbilidad y mortalidad, sin que esto signifique, por otra parte, que el predominio del tipo "mitis" entre nosotros nos ponga en condiciones deficientes de inmunización, porque parece no haber diferencia entre los tipos "gravis" y "mitis", como agentes inmunizantes naturales (DUDLEY y col., 1934). Por lo demás, todo esto no tiene el carácter de una afirmación, hasta que no se hagan estudios más amplios y comparativos.
En algunos países como el nuestro, donde no es frecuente la difteria, se han encontrado porcentajes pequeños de portadores de C. hofmanii; así en la India, informa ANDREWES y col. (1923), no pasan de 5 % los niños portadores de este germen, pero ellos aumentan hasta 50 % cuando se presentan mayor número de casos de difteria clínica.
Por otro lado, el hecho de haber encontrado entre nosotros un 4% de portadores de C. hofmanii, proporción relativamente alta y además, si estamos de acuerdo con lo comunmente aceptado de que esta especie de Corynebacterium no patógeno, se disemina de garganta en garganta por el mismo mecanismo que sirve para la diseminación del tipo patógeno, podemos concluir afirmando que en nuestra colectividad, debe haber excelentes condiciones para la difusión de ambos gérmenes, y en consecuencia, sino tenemos grandes epidemias es seguramente debido a otros factores muy distintos a la falta de condiciones que permitan el pasaje del germen de huesped a huesped.
Por último, el hecho de que hayamos encontrado portadores de C. hofmanii en niños indistintamente Schick negativos o positivos, nos dice que no existe relación entre este presumible saprofito y el bacilo diftérico, al menos en lo que se refiere a su valor antigénico.
CONCLUSIONES
1° Es de urgente necesidad establecer técnicas y métodos simples para el reconocimiento del C. diphtheriae y uniformar el criterio en el concepto de "portadores".
2° La tasa de "portadores" de C. diphtheriae virulento entre nosotros, fué de 1% y todos los niños reconocidos como tales se mostraron Schick-negativos.
3° En el momento en que hemos realizado esta investigación, hemos encontrado un predominio de portadores de tipo "mitis" sobre el "gravis".
4° El porcentaje de niños portadores de C. hofmanii fué de 42% comprendiendo a niños indistintamente Schick-positivos o negativos.
5° Por todo lo anteriormente expuesto se puede afirmar que, la propagación del C. diphtheriae lo mismo que la inmunidad natural antidiftérica, se establece y mantiene en nuestra población estudiada, por influencia de los mismos factores demostrados para otros lugares.
BIBLIOGRAFIA
1. AJO, CORRADO, Comptes Rendues Societé de Biologie, v. 70, p. 842. 1935. [ Links ]
2. ALLISON, V. D. y ALLING, T. H., J. Pathology & Bacteriology, v. 32, p. 299, 1939. [ Links ]
3. ANDERSON, J. S., HAPPOLD, F. C., Mc. LEOD, J. W. y THOMPSON, Pathology & Bacteriology, v. 36, p. 169. 1931. [ Links ]
4. ANDERSON, J. S., COOPER, K. E., HAPPOLD, F. C. y Mc. LEOD, J. W. J. Pathology & Bacteriology, v. 36, p. 169, 1933. [ Links ]
5. ANDREWES, F. W., BULLOCH, W., DOUGLAS, S. R., DREYER, G., FILDES, P., LEDINGHAM, J., y WOLF, C. G., Diphtheria, Medical Research Council, London. 1923. [ Links ]
6. BARRATT, M. N., Journal of Hygiene, v. 23, p. 241. 1924. [ Links ]
7. BERGEY, D. H., Manual of Determinative Bacteriology. 1939. [ Links ]
8. CARRIEU, JANBON, RAMBAULT y ALQUIE, Comptes Rendues Societé de Biologie, v. 76, p. 285. 1937. [ Links ]
9. CARVALHO LIMA, Bacteriologia. 1934. [ Links ]
10. COOPER, K. E., HAPPOLD, F. C., JOHNSTONE, K. I., Mc. LEOD, J. W., WOODCOCK, H. E., y ZINNEMANN, K. S., Lanoet, v. 238, p. 865. 1940. [ Links ]
11. DOPTER, M., Revue d'Immunologie, v. I, p. 305. 1935. [ Links ]
12. DUDLEY, S. F., Medical Research Council, Special Report, N° 75. 1923. [ Links ]
13. DUDLEY, S. F., MAY, P. M. y O'FLYNN, J. A., Medical Research Council, Special Report, N° 195. 1934. [ Links ]
14. FROBISHER, M. JR., American Journal of Hygiene, v. 28, p. 13. 1938. [ Links ]
15. FROBISHER, M. JR., American Journal of Hygiene, v. 28, p. 1. 1938 a. [ Links ]
16. FROBISHER, M. JR., American Journal of Public Health, v. 32, p. 709. 1942. [ Links ]
17. GOLDIE, H., Comptes Rendues Societé de Biologie, v. 62, p. 1210. 1933. [ Links ]
18. GROOTTEN, O., Revue d'Immunologie, v. 6, p. 44. 1940. [ Links ]
19. HORGAN, E. S., MARSHALL, A., Journal of Hygiene, v. 32, p. 544. 1932. [ Links ]
20. KERRIN, J. C. y GAZE, H. W., Journal of Hygiene, v. 37, p. 280. 1937. [ Links ]
21. MAIR, W., J. Pathology & Bacteriology, v. 33, p. 230. 1930. [ Links ]
22. MEMORIAS DEL HOSPITAL DEL NIÑO, 1933 a 1938. [ Links ]
23. MOSHAGE, E. y KOLMER, J., Journal of Infectious Diseases, v. 19, p. 19. 1916. [ Links ]
24. NEILL, G. A. W., Journal of Hygiene, p. 552, v. 37. 1937. [ Links ]
25. OKELL, C. C., y PARISH, H. J., Journal of Hygiene, v. 25, p. 363. 1926. [ Links ]
26. PETERSON H. N., American Journal of Public Health, v. 29, p. 664. 1939. [ Links ]
27. PISO, ITALO, Diagnostica e, Tecnica di Laboratoria, v. 10, p. 643. 1939. [ Links ]
28. RONDON, O., Revista de Medicina Experimental, v. II, p. 449. 1943. [ Links ]
29. SCHOROEDER, R., Comptes Rendues Societé de Biologie, v. 55, p. 1347. 1930. [ Links ]
30. SCHOROEDER, R., Comptes Rendues Societé de Biologie, v. 55, p.91. 1930. [ Links ]
31. TEMPE, G., Comptes Rendues Societé de Biologie, v. 54, p. 1339. 1930. [ Links ]
32. TOPLEY, W. W., y WILSON, Tratado de Bacteriología e Inmunología, Londres. 1942. [ Links ]
33. WELSCH, M. y THIBAULT, J., Comptes Rendues Societé de Biologie, v. 132, p. 319. 1939. [ Links ]
34. WELSCH, M. y THIBAULT, J., Comptes Rendues Societé de Biologie, v. 132, p. 322. 1939 a.
[ Links ]

