Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.3 n.2 Lima jun. 1944
TRABAJOS ORIGINALES
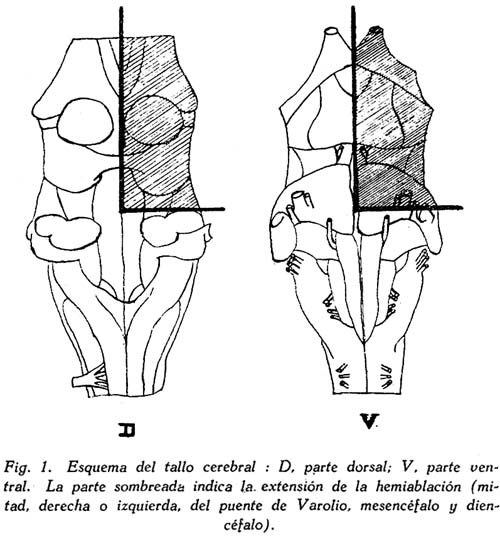
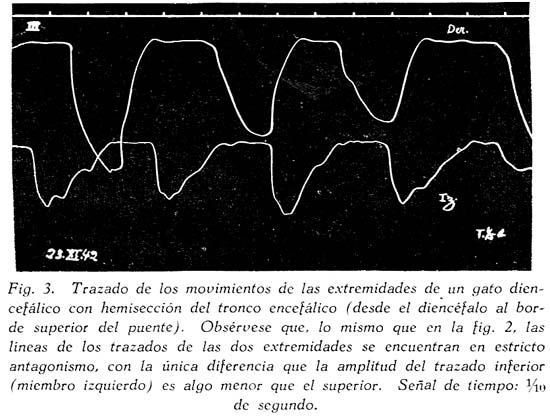
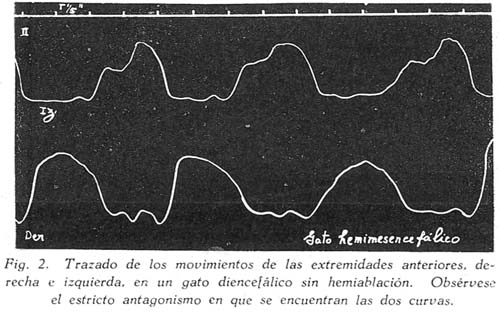
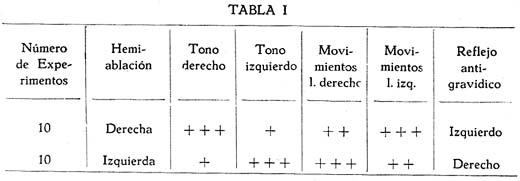
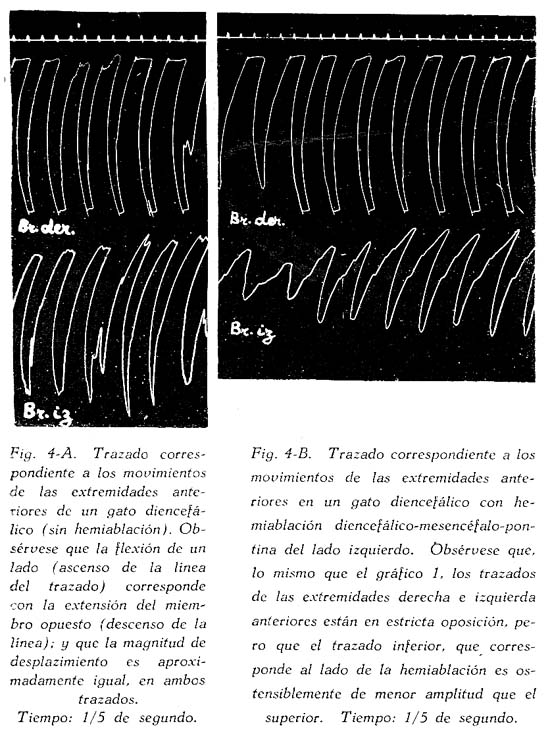
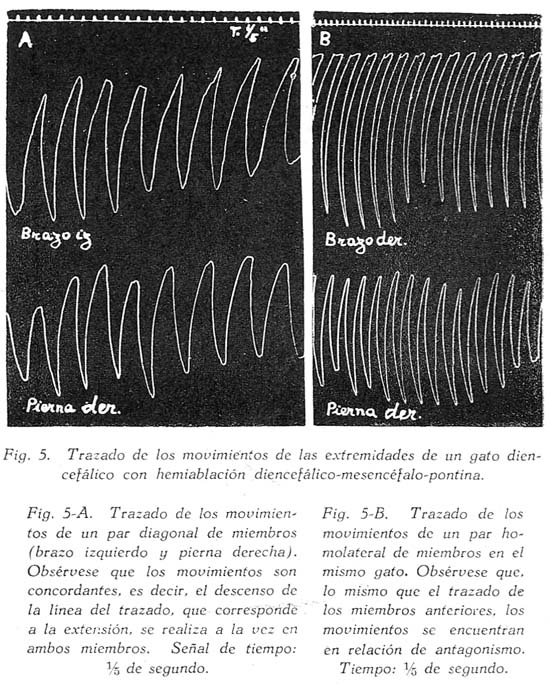
Sobre la ritmicidad de los movimientos de locomoción Carlos Gutiérrez-Noriega1 1 Departamento de Farmacología, Instituto Nacional de Higiene y Salud Pública. SUMARIO Se han estudiado los movimientos rítmicos de flexión y extensión de las extremidades (movimientos de la marcha) en gatos descerebrados con sección longitudinal mediana o con ablación unilateral o hemiablación del tallo cerebral. Los resultados fueron los siguientes : 1. La sección longitudinal mediana no altera la coordinación de los movimientos. 2. La hemiablación longitudinal o ablación unilateral del tallo cerebral debilita la intensidad de los movimientos en los miembros del lado de la ablación, pero no los suprime. Se debilitan, y en algunos casos hasta se extinguen, principalmente los movimientos de flexión de los segmentos distales de los miembros del lado de la hemiablación, sin modificarse la ritmicidad de los segmentos proximales. 3. La destrucción de los tubérculos cuadrigéminos no altera la ritmicidad. La destrucción de la substancia intercolicular la suprime definitivamente. 4. De acuerdo a los resultados experimentales se ofrece una explicación del mecanismo neurofisiológico de los movimientos rítmicos. SUMMARY Rhythmic movements of flexion and extension of the extremities (stepping movements) were studied in decerebrated cats by means of longitudinal median section and unilateral ablation or hemiablation of the brain stem and hypothalamus. The following results were observed : 1. Longitudinal median section does not alter the coordination of movements. 2. Unilateral ablation of the brain stem and hypothalamus weakens the energy of the movements of the extremities on the side of the ablation. The alteration occurs principally in the movements of flexion of the distal segments of the extremities of the same side, but the rhythmicity of the proximal segments remains. 3. Destruction of the superior and inferior colliculus does not alter the rhythmicity, but destruction of the region situated between right and left colliculus definitely does destroy rhythmicity. 4. In accordans with the results obtained with these experiments, an explanation of the neurophysiological mechanism of rhythmic movements is offered. Las investigaciones sobre la fisiología de los movimientos de locomoción del hombre y de los animales no han esclarecido en forma completamente satisfactoria los mecanismos de su ritmicidad. Se concede gran importancia a la medula espinal, en la que se originarían los reflejos fundamentales de los movimientos deambulatorios, pues los reflejos flexor directo y extensor cruzado, y la inducción sucesiva, son en última instancia fragmentos funcionales de naturaleza estrictamente espinal de los movimientos deambulatorios. Las observaciones sobre los movimientos rítmicos de las extremidades de gatos decapitados permiten, sin duda alguna, sostener este punto de vista. "Podemos interpretar la locomoción, dice HÖBER al sintetizar sus ideas sobre este problema, como resultado del encadenamiento de una serie de reflejos espinales. Las observaciones realizadas en el perro espinal pueden ser utilizadas también en la explicación de la locomoción humana" (6). Alude principalmente a los experimentos de BERITOFF (2), SHERRINGTON (19), PHILIPPSON (9), LAUGHTON (8), y FREUSBERG. Al lado de las numerosas observaciones sobre el origen espinal de la ritmicidad de la marcha conviene considerar las no menos importantes sobre la participación en el mismo proceso funcional del tronco encefálico, en especial del hipotálamo. Por cierto, no es posible aceptar en la actualidad la hipótesis de un centro de la marcha, hace algunos años sostenida por BECHTEREW (1), según el que "en los núcleos del puente, y en el núcleo reticulado, poseemos un extenso ganglio motor, por el intermedio del cual se transmiten los impulsos de la estación y de la locomoción que vienen de diferentes regiones-del cerebro". De todas maneras, sus observaciones, y las de otros fisiólogos modernos, llamaron la atención sobre la importancia de la región suprabulbar en la locomoción de los vertebrados. El animal espinal e incluso el bulbar carecen por completo de movimientos espontáneos. Los movimientos rítmicos de las extremidades que en ellos se han observado, son exclusivamente de origen reflejo (por estímulos eléctricos o mecánicos) y además carecen de continuidad. La presencia del mesencéfalo y más aún del hipotálamo permite reacciones locomotrices, o simplemente movimientos rítmicos de las extremidades, mucho más perfectos y prolongados, y sobre todo no sólo de origen reflejo, sino aparentemente espontáneos. Además, el estímulo por medio de drogas (coramina, cardiazol, insulina) que nunca determina estos movimientos en los perros o gatos espinales, fácilmente los origina en los animales mesencefálicos o hipotalámicos (5). Sobre todo la participación del tono, que tan importante es para el acto de progresión, es de origen bulboprotuberancial. Pero es además muy verosimil, como lo demuestran algunas observaciones experimentales (5), que los núcleos del tallo cerebral y sobre todo los subcorticales originen en parte el proceso rítmico de extensiones y flexiones alternativas. En cuanto al mecanismo de dicho proceso, existen dos teorías. La primera de GRAHAM BROWN (3) quien piensa que la ritmicidad de la marcha se debe a un proceso de inducción recíproca entre centros flexores y extensores situados simétricamente a derecha e izquierda del neuroeje. La sección longitudinal y mediana del tronco encefálico suprimirla la ritmicidad. Un punto de vista diferente mantiene SHERRINGTON (10) para quien "en el paso natural, las influencias excitantes que descienden del cerebro a los centros espinales de los miembros no son intermitentes sino continuas, aunque dan por resultado un movimiento rítmico alternativo de los miembros". La alternancia y ritmicidad de los movimientos se debe, en última instancia, a reflejos antagónicos que toman posesión de la vía final común en sucesión rítmica (succesive induction). De esta manera, considera que la ritmicidad de los movimientos de la marcha es un reflejo espinal (spinal stepping), que se puede suscitar por estímulos externos o por excitaciones directas de los centros superiores. Con el propósito de comprobar la exactitud de estas hipótesis, hemos verificado en gatos descerebrados ablaciones unilaterales hipotalámico-mesencéfalo-protuberanciales; y simples incisiones medianas en toda la extensión de la misma región. MÉTODO Los gatos fueron descerebrados en narcosis etérea, por sección transversa de la parte superior de los tubérculos cuadrigéminos anteriores al quiasma óptico. Se extirpó todo el tejido nervioso por encima de al sección. Luego se puso a descubierto la formación cuadrigémina por exéresis del tentorium, y separando cuidadosamente los tubérculos cuadrigéminos inferiores del cerebelo. Con un fino bisturí se verificó una sección longitudinal, exactamente en la parte media de la zona intercolicular por atrás, y entre los pedúnculos cerebrales y 109 cuerpos mamilares por delante. En los casos en que se practicó la ablación unilateral se hizo una sección transversa en la parte inferior desde la línea media, partiendo de la incisión longitudinal, hacia afuera, extirpándose a continuación todo el tejido nervioso comprendido entre las dos incisiones, incluyendo los tubérculos de un lado, el pedúnculo y el cuerpo mamilar correspondientes, y también en ciertos casos una mitad de la protuberancia. El preparado en estas condiciones conservaba el cuerpo mamilar, los tubérculos cuadrigéminos y la mitad del puente del lado opuesto. La fig. 1, representa este tipo de ablación. La pupila, que en estas condiciones debe presentar completa midriasis en el lado de la ablación unilateral y miosis en el lado indemne, sirve de referencia respecto a la estricta unilateralidad de la ablación. La observación de los movimientos rítmicos, espontáneos o provocados, se inició una o dos horas después de la descerebración. En los casos en que no se presentaron espontáneamente se les suscitó por estímulo farádico o por inyección endovenosa de coramina. Aunque a simple vista se puede apreciar la intensidad y ritmicidad de los movimientos, se obtuvo registros de ellos, conectando cada extremidad con un sistema de palancas y con una aguja inscriptora, A fin de estudiar la ritmicidad de las flexiones y extensiones de los músculos de un solo miembro, se hizo la disección de un flexor y un extensor, y se les conectó por medio de un sistema de palancas y poleas con agujas inscriptoras. Los músculos principalmente estudiados fueron el biceps brachii (flexor del brazo) y el caput longum del triceps brachii (extensor del brazo). Tanto la observación ad oculos como los trazados, permitieron apreciar la simetría o asimetría de los movimientos. Después del experimento el tallo cerebral fué endurecido con formol y estudiado, para la exacta determinación de las secciones longitudinales y áreas de ablación. RESULTADOS En primer lugar se refieren las observaciones en animales decorticados con sección longitudinal mediana del tallo cerebral; en segundo, las observaciones en animales con hemiablación o ablación unilateral del hipotálamo, mesencéfalo y protuberancial. La figura 1, representa las áreas comprendidas en estas dos operaciones. En tercer término se considera experimentos adicionales. Incisión longitudinal mediana del tallo cerebral. Cuando en un animal decorticado se practica una incisión vertical mediana, desde el hipotálamo al puente, dividiendo completamente en dos mitades al tallo cerebral en sentido anteroposterior, se produce en seguida midriasis y rigidez descerebrada. Los impulsos nerviosos hipotalámicos-mesencefálicos inhibidores de los centros bulbo-protuberanciales del tono pasan, probablemente, por la línea media. Su destrucción por la incisión longitudinal mediana del tallo cerebral es causa de la rigidez, aunque se conserven las dos mitades izquierda y derecha del mesencéfalo y del hipotálamo. En estas condiciones no se presentan movimientos rítmicos espontáneos de las extremidades y el animal se parece en todo a un preparado bulbo-protuberancial. Pero si se practica una inyección endovenosa de coramina (a dosis de 25 a 70 mgr. por kilo de peso) se produce típicos movimientos rítmicos, como en los animales decorticados o mesencefálicos sin incisión longitudinal. En la fig. 3, presentamos el trazado de los movimientos deambulatorios de los miembros anteriores en un gato decorticado con incisión longitudinal mediana del tallo cerebral. Se verificaron cinco experimentos de este tipo, uno de los cuales referimos a continuación: Experimento N° 2. Gato de 1.3 Kgr. A las 10.50 a. m., descerebración por el método ya descrito (sección transversal del borde superior de los tubérculos cuadrigéminos al borde superior del quiasma óptico). 11 a. m. Sección longitudinal mediana, del hipotálamo al puente, pasando entre los cuerpos mamilares y los pedúnculos cerebrales por delante, y entre los colículos derechos e izquierdos por atrás. Inmediatamente después de esta operación se observó midriasis y rigidez de las extremidades anteriores. 12.15 p. m. Persiste la rigidez y la midriasis. Los reflejos corneales están abolidos, la respiración es regular y lenta. 12.20 p. m. Inyección intrayugular de 25 mgr. x Kgr. de Coramina. Después de 18" aumento de la hipertonía y de la respiración. Dos minutos más tarde se inicia débiles movimientos rítmicos de las extremidades posteriores, y luego más intensos de las anteriores. 12.40 p. m. Inyección intrayugular de 50 mgr. x Kgr. de Coramina. Después de 15" débiles convulsiones tónicas en flexión, y luego convulsiones clónicas. Dos minutos más tarde intensos movimientos rítmicos de las extremidades, que se inician en las posteriores y se propagan después de las anteriores. Estos movimientos son perfectamente simétricos y no difieren en nada de los movimientos correspondientes de los descerebrados mesencefálicos sin sección longitudinal. Compárese el trazado de los movimientos de un animal hipotalámico y sin sección longitudinal (fig. 2) con el trazado de los movimientos del animal del presente experimento (fig. 3), y no se encontrará gran diferencia. La autopsia demostró que la sección longitudinal llegaba hasta la parte mediana de la protuberancia. La incisión longitudinal mediana del tallo cerebral, indudablemente, no altera la coordinación de los movimientos rítmicos de las extremidades. Si estos, y por supuesto también la deambulación normal, fueran el resultado de la interdependencia funcional de dos centros simétricos por inducción recíproca, la incisión suprimirla la coordinación de los movimientos de las cuatro extremidades; mas no ocurre así. Nuestros resultados concuerdan con, las antiguas observaciones experimentales de TARCHANOFF (1) que practicó una incisión longitudinal sobre el bulbo de la rana descerebrada sin que perdiera su capacidad de arrastrarse. Se deduce de ello "que el acto de la reptación se verifica gracias a la existencia de líneas, directas y entrecruzadas del centro (unilateral) de la locomoción con los centros subyacentes de la médula". La incisión longitudinal suprime o debilita los movimientos rítmicos, hace aparecer la rigidez descerebrada, y el animal en conjunto se comporta como un descerebrado protuberancial o bulbar, con la única diferencia que un enérgico estímulo, la. inyección de coramina por ejemplo, suscita los movimientos mencionados. La división de la parte hipotalámica-mesencefálica del tronco cerebral en dos mitades tiene más efecto sobre el tono que sobre la coordinación de los movimientos deambulatorios, porque posiblemente los impulsos inhibidores de los centros bulbo-protuberanciales tonígenos descienden por haces, estrictamente medianos, y serían destruídos por la incisión; o porque los mismos impulsos siguen en parte, por vías cruzadas (ej. el haz de Monakow) que son totalmente seccionados por el corte longitudinal. Si se tiene en cuenta que en el síndrome de Benedikt se presenta hipertonía contralateral al núcleo rojo lesionado, y que de las experiencias de MAGNUS & KLEIYN y de RADEMAKER también se infiere una relación entre este núcleo y el tono, la hipertonía que se obtuvo inmediatamente después de la incisión longitudinal sería originada, en parte al menos, por la destrucción de la decusación de Forel. Sin embargo, RANSON (7) no encontró ninguna modificación del tono al destruír el núcleo rojo. Finalmente, hay que tener en cuenta que la incisión mediana longitudinal, además de destruir otras decusaciones (de Maynert o decusación tecto-bulbar, las comisuras intercoliculares y la decusación del brazo conjuntivo), destruye también el importante fascículo longitudinal mediano, y en parte el ganglio interpenduncular. Así, la hipertonía que provoca la sección longitudinal debe ser la consecuencia de múltiples factores. Hemiablación longitudinal del hipotálamo, mesencéfalo y protuberancia. Llamamos hemiablación longitudinal a la exéresis de la mitad izquierda o derecha, sin sobrepasar la línea media, de cualquiera de las porciones del tallo cerebral. Es preciso distinguir la ablación longitudinal de la transversa, que consiste en la exéresis total de la mitad superior (del mesencéfalo o del puente). En los experimentos correspondientes a este grupo se extirpó una mitad del hipotálamo del mesencéfalo y de la protuberancia, procurando no exceder la línea media. El examen de las pupilas constituye un punto de referencia útil como medio de control. Si la ablación no sobrepasa la línea media, si no compromete la integridad del lado opuesto, se observa completa midriasis en el lado extirpado, y completa miosis en el intacto. Si la lesión. excede bilateralmente la línea media, la midriasis es bilateral. Por lo regular, después de la hemiablación longitudinal del tallo cerebral se observa rigidez de descerebración en el lado extirpado y tono normal en el opuesto, (v. la tabla I, en la que la intensidad del tono está indicada por el número de cruces). Pero esta asimetría en el tono desaparece paulatinamente, y al cabo de media o una hora se extingue la hipertonía del lado lesionado. ¿Son las fibras hipotalámico-mesencefálicas inhibidoras de la rigidez, de acción bilateral Si es así, el entrecruzamiento al lado opuesto se haría a nivel bulbar. En algunos experimentos se observó que las diferencias en la intensidad del tono no eran latero-laterales, sino anteroposteriores, con predominio del tono en los miembros anteriores o posteriores. No fué posible establecer relaciones con la lesión anatómica en este caso. Se observó también abolición de los reflejos antigravídico y corneal del lado de la lesión (v. la tabla I). Si bien en los animales con hemiablación se presentan asimetrías funcionales del tono general, de algunos reflejos y del tono pupilar, no ocurre lo mismo con los movimientos rítmicos de las extremidades. La asimetría funcional en este caso es menos evidente o aparentemente no existe. Si se observan los movimientos ad óculos sólo en algunos casos los del lado intacto son más perfectos que los del lado lesionado (v. tabla I). Con el registro de dichos movimientos se suelen descubrir asimetrías no aparentes a simple vista. En la fig. 4, presentamos los movimientos de los brazos derecho e izquierdo de un animal decorticado sin hemiablación (A) y de un animal decorticado con hemiablación hipotalámico-mesencefálica-protuberancial izquierdo (B). En ningún caso se observó movimientos exclusivamente, unilaterales. La ablación longitudinal afecta mucho más a los movimientos rítmicos de flexión y extensión de los segmentos distales del miembro que los segmentos proximales. Este resultado se advierte sobre todo en los casos en que los miembros del lado de la exéresis están rígidos y los del intacto no. En este caso, mientras que en estos últimos los movimientos consisten en una triple flexión y una triple extensión del miembro, los primeros presentan movimientos de flexión sólo en la parte proximal del miembro, (brazo muslo), permaneciendo rígidos los segmentos distales. Otras veces sólo se observan movimientos en torno al hombro o a la cadera de un miembro rígido (del lado de la lesión nerviosa), sin flexión propiamente dicha. Existen en consecuencia, dos factores relacionados con los movimientos rítmicos de la marcha : el primero con los músculos de la parte proximal del miembro, y sobre todo con los músculos del hombro y de la cadera; el segundo con los músculos de las partes distales (antebrazo, pierna, mano y pie). Gracias al primer factor se desarrolla el conjunto armónico de flexiones y extensiones de la marcha normal. El primer factor, el más primitivo y elemental mecanismo de la marcha, persiste aún después de la hemiablación, longitudinal del hipotálamo, del mesencéfalo y de la protuberancia. El segundo factor requiere la participación de aquellas estructuras. El estudio de la ritmicidad de la marcha por medio del registro directo de las contracciones musculares nos permite establecer lo siguiente : a) El primer factor consiste principalmente en un aumento del tono extensor. Se trata pues, de descargas rítmicas de impulsos hipertónicos, para los músculos extensores, alternativamente al lado derecho y al lado izquierdo. b) El segundo factor consiste en un aumento del tono flexor con inhibición simultánea del tono extensor. Como en el caso de la marcha no es necesario que los músculos flexores tengan una actividad tan considerable como los extensores, se infiere que en este caso la inhibición de estos últimos es uno de los procesos más importantes. El segundo factor se compone, indudablemente, de impulsos hipertónicos para los músculos flexores, e impulsos inhibidores para los extensores. Estos impulsos flexores son de naturaleza hipotalámico-mesencefálica;* y como está perfectamente demostrado que los impulsos inhibidores de la hipertonía de los extensores tienen el mismo origen, se explica porque, después de la hemiablación longitudinal hipotalámico mesencefálica, hay mengua de la actividad rítmica de los segmentos distales de las extremidades (es decir mengua de les movimientos rítmicos de flexión e inhibición de los extensores). Pero, si bien es cierto que muchas observaciones experimentales demuestran que la región hipotalámica-mesencefálica tiene el control de los flexores y la región bulbo-protuberancial de los extensores, de ninguna manera se podría inferir que la ritmicidad de uno y de otro grupo de músculos se encuentra regida por los núcleos de las regiones mencionadas. En un trabajo anterior (5) hemos demostrado que el bulbo casi carece de actividad rítmica y la protuberancia sólo la posee muy escasamente. En consecuencia, los impulsos rítmicos de la marcha, tanto para los músculos extensores como para los flexores, proviene de la región hipotalámico-mesencefálica, y es casi seguro que tiene acción bilateral, especialmente en el primer caso. Ya hemos dicho que la hemiablación longitudinal no suprime la ritmicidad; sólo debilita los movimientos rítmicos homolaterales. Nuestros experimentos demuestran que los centros de cada lado tienen una actividad rítmica bilateral. Experimento N° 15. Gato de 2 Kgr. Descerebración a las 10.40 a. m. 10.45 a. m. Ablación de la mitad derecha del hipotálamo, mesencéfalo y protuberancia. Inmediatamente después de esta operación se produjo hipertonía de los miembros anteriores, predominante al lado derecho. Mediana hipertonía en los miembros posteriores, predominante en el derecho. Midriasis sólo a la derecha. Abolición del reflejo corneal del mismo lado. Desviación de la cabeza hacia el lado izquierdo. Miosis del mismo lado. 12 m. En las mismas condiciones. 1 p. m. Al estimular la piel con una corriente farádica se producían movimientos rítmicos de flexión y extensión en las cuatro extremidades. Los movimientos de los miembros del lado izquierdo presentan la característica triple flexión de la deambulación. En el lado derecho el movimiento es menos perfecto, Pues las partes distales permanecen rígidas en vez de flexionarse, predominando la actividad rítmica en el segmento proximal. No obstante la ritmicidad con los movimientos del miembro del lado opuesto no está alterada. 1.05 p. m. Inyección endovenosa de 25 mgr. de Coramina por kilo de peso. Sólo se obtuvo aumento de la rigidez descerebrada en ambos lados. 1.10 p. m. Inyección de 75 mgr. de coramina por kilo. Se obtuvo primero una crisis de movimientos irregulares, con sacudidas clónicas intermitentes. Luego, intensos movimientos rítmicos de las cuatro extremidades durante dos minutos. La rapidez de los mismos no permitió establecer diferencias entre los del lado derecho y del izquierdo. Se observó además que los movimientos rítmicos aparecieron primero en los miembros anteriores y algunos segundos más tarde se propagaron a los posteriores. Protocolo : A la autopsia se encontró que las mitades derechas del hipotálamo y del mesencéfalo habían sido completamente extirpadas. También se demostró la ablación de los dos tercios superiores de la mitad derecha del puente de Varolio. Las mitades izquierdas del hipotálamo, mesencéfalo y puente se encontraban intactas. La sección mediana pasaba exactamente entre los cuerpos mamilares, entre los tubérculos cuadrigéminos izquierdos y derechos y a lo largo del acueducto de Sylvio. Para determinar la coordinación de los movimientos de los miembros anteriores con los posteriores en los animales descerebrados con hemiablación longitudinal hipotalámica-mesencefálica-pontina, se obtuvieron trazados de los movimientos de los dos miembros diagonales (fig. 5, A) y de los miembros homolaterales, anteriores y posteriores (fig. 5, B). En el primer caso se observó que los movimientos de flexión de extensión eran simultáneos. Por, ejemplo, la flexión del miembro anterior izquierdo coincidía con la flexión del miembro posterior izquierdo, (v. fig. 5, A).- Al contrario, en el trazado de los miembros homolaterales, la extensión del anterior coincidía con la relajación del posterior, y a la inversa, como puede observarse en la figura (v. fig. 5, B). Todo lo cual indica que la destrucción de los centros motores del lado izquierdo o derecho del hipotálamo o del mesencéfalo no altera la coordinación, de los movimientos en sentido anteroposterior. Para interpretar correctamente los resultados es necesario tener en consideración los diferentes tipos de coordinación de las extremidades del gato. Durante la marcha cada par de miembros, anterior o posterior, funciona alternativamente, simulando movimientos de pedal; mientras que la coordinación anteroposterior se hace de tal suerte que cada miembro anterior realiza el movimiento opuesto al posterior del mismo lado, y el mismo movimiento que el posterior del lado opuesto. Es decir, en la marcha normal la coordinación latero-lateral consiste en movimientos asimétricos, de extensiones y flexiones alternativas; mientras que los movimientos del par diagonal de miembros son simétricos, coincidiendo la extensión de un par diagonal con la flexión del par diagonal opuesto. Durante la carrera la coordinación funcional se modifica. Si los movimientos no son muy rápidos, el par anterior de miembros conserva su coordinación ordinaria, mientras que los dos miembros posteriores realizan movimientos simétricos y rítmicos de extensión y relajación. En el movimiento de galope tanto el par anterior como el posterior realizan movimientos simétricos. En este último caso cuando los dos, miembros anteriores se extienden los posteriores se relajan. Destrucción de los tubérculos cuadrigéminos o de la región intercolicular. Puesto que la calota mesencefálica tiene tan definida participación en el origen de los movimientos de las extremidades, realizamos la destrucción de la lámina cuadrigémina para averiguar su importancia en el proceso. La destrucción fué realizada por medio de termocauterio. Los animales descerebrados hipotalámicos o mesencefálicos pero con tubérculos cuadrigéminos destruídos reaccionaron en forma idéntica a los animales con tubérculos intactos. Tampoco la aplicación de papel secante imbibido en solución de coramina sobre los tubérculos del lado izquierdo o derecho, o, sobre ambos, reveló alguna reacción. Al contrario, las lesiones de la substancia intercolicular, incluso la simple incisión en la línea media con un bisturí sin filo, basta para suprimir definitivamente los movimientos rítmicos de las extremidades, DISCUSIÓN La sección longitudinal mediana y la ablación unilateral o hemiablación del tallo cerebral no alteran la coordinación de los movimientos rítmicos de flexión y extensión de las cuatro extremidades. La destrucción de los tubérculos cuadrigéminos tampoco afecta los movimientos mencionados. Al contrario, la destrucción de la substancia intercolicular los suprime. Según estos resultados, la hipótesis emitida por GRAHAM BROWN para explicar la coordinación de los movimientos de la marcha no debe aplicarse a los centros motores del tallo cerebral. Entre los movimientos rítmicos y el tono existen relaciones. La flexión de un par de extremidades, anterior y posterior, implica la inhibición de los centros tónicos bulbo-protuberanciales. Al contrario, el movimiento de extensión -que permite a los miembros convertirse en dos columnas rígidas para mantener la actitud erguida de la marcha- implica el prevalecimiento de los centros tónicos bulbo-protuberanciales del lado correspondiente y la inhibición de los centros mesencefálicos de los impulsos flexores. Puesto que la sección mediana longitudinal y la hemiablación del tallo cerebral sólo debilitan la ritmicidad pero no la suprimen, es preciso admitir que de los centros de cada lado parten impulsos rítmicos bilaterales. Por ejemplo, de los centros hipotalámico-mesencefálicos derechos parten alternativamente impulsos flexores para los músculos del mismo lado y para los músculos del lado opuesto. Como se ha observado que los movimientos de flexión del lado de la ablación no son tan perfectos como los movimientos flexores del lado intacto -por lo menos en los segmentos distales del miembro- se debe aceptar que los impulsos de flexión cruzados tiene valor secundario. En otras palabras, los movimientos de flexión de la marcha del gato se originan principalmente de impulsos nerviosos de los centros del mismo lado, reforzados por impulsos de los centros del lado opuesto. Por eso cuando se destruyen los centros de un lado la acción flexora bilateral de un solo centro mantiene la ritmicidad. No sabemos si los impulsos flexores directos y los cruzados proceden de las mismas o de diferentes neuronas, siendo mucho más probable que se trate de diferentes sistemas de neuronas. Existe otra posible explicación concordante a nuestros resultados experimentales : los impulsos procedentes de los centros de un lado originarían los movimientos de flexión homolaterales y una acción tónica estimulante heterolateral sobre los músculos extensores. Variaciones rítmicas del tono extensor en el lado de la ablación pueden producir los movimientos rítmicos imperfectos que fueron observados en los animales con hemiablación longitudinal. Si los centros hipotalámico-mesencefálicos de un lado ejercen una acción inhibidora sobre los centros del lado opuesto no sería gracias a simples conexiones transversales, sino en forma mucho más compleja, por intermedio de centros inferiores y por vías descendentes, pues la sección longitudinal mediana no destruye la coordinación. NOTAS * En el gato intacto el Cardiazol produce convulsiones tónicas primero en flexión y luego, en extensión. En los gatos hipotalámicos y mesencefálicos se obtiene el mismo resultado. En los gatos bulbares sólo se presentan convulsiones tónicas de los extensores. BIBLIOGRAFÍA 1. W. BECHTEREW : Les fonctions Nerveuses. Les fonctions bulbo-médullaires, París, 1909. [ Links ] 2. J. S. BERITOFF : Aechiv für die gesamte Physiologie, v. 199, P. 248, 1922. 3. T. GRAHAM BROWN : Procedings of the Royal Society of London, v. 85, p. 278, 1912. 4. T. GRAHAM BROWN : Procedings of the Royal Society of London, v. 84, p. 1912. 5. C. GUTIÉRREZ-NORIEGA : Revista de Neuro Psiquiatría, v. 4, p. 333, 1941. 6. R. HÖBER : Fisiología Humana (trad. Fernández Galindo), Barcelona, 1928. 7. W. R. INGRAM & S. W. RANSON : Archives of Neurology and Psychiatry, v. 28, p. 483, 1932. 8. N. B. LAUGHTON : American Journal of Physiology, v. 70, p. 358, 1924. 9. M. PHILIPPSON : Travaux du Lab. de Physiol. Institute Solvay, Bruxelles, 1905. 10. CH. SHERRINGTON : Journal of Physiology, v. 40, p. 28, 1910.