










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
Print version ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.4 no.1 Lima Mar. 1945
TRABAJOS ORIGINALES
Síntesis de argumentos experimentales en favor de la llamada Función X en el Suero Antitifoideo
Alois Bachmann1
1 Miembro de la Academia Nacional de Medicina de Buenos Aires
Hace cerca de diez años que estoy estudiando nuevos elementos en el suero antitifoideo, nacidos, a mi juicio, en el organismo a expensas de la excitación de antígenos o cuerpos nuevos formados por la Salmonella typhi en el medio interno infestado. Elementos que desempeñan un papel importante en la inmunidad y se manifiestan aparentemente de diferente manera que los anticuerpos comunes.
Para entrar, en materia haremos una breve reseña del mosaico de antígenos del, bacilo de Eberth. Según WHITTE las Salmonellas tienen un núcleo central más o menos común, el que constituye la forma R apatógena, tanto que el suero anti R de una de ellas aglutina más o menos bien a todas las otras formas similares, aún cuando tengan antígenos ciliares diferentes. Por encima de este núcleo se encuentra la caparazón agresiva de las formas patógenas S con características especiales para cada grupo, lo que permite distinguirles fácilmente uno de otros. El diagnóstico de la especie se efectúa por la combinación de estos antígenos con los ciliares.
En el bacilo de la fiebre tifoidea esa envoltura está constituida especialmente por dos capas superpuestas, la más profunda formada por un antígeno lípido glúsico denominado IX según la clasificación de KAUFFMANN WHITTE, parcela de constitución similar, probablemente no completamente igual se encuentra en todo el grupo D, de las salmonellas. Además tiene otro propio llamado Vi de igual estructura, química a la IX y un tercero de menor importancia el XII, que se encuentra generosamente repartido entre numerosas salmonellas y que no desempeña papel en la inmunidad en contra de este microbio. Por encima de todo sellos se encuentran las cilias comunes a las dos formas la R y la S, constituyendo el antígeno ciliar, en el caso de EBERTH, el llamado d, que es de naturaleza proteica, el que no interviene activamente ni en la virulencia ni en la inmunidad de estos gérmenes, por lo cual solo lo mencionaremos de paso en el curso de esta publicación.
Los estudios de KAUFFMANN que luego han sido conformados, demostraron que los gérmenes de las colonias S de la Salmonella typhi pueden presentarse según la existencia y la cantidad que contengan del antígeno Vi en tres formas bien diferentes. 1º La forma W que contiene el antígeno IX y está desprovista del Vi, es poco virulenta, se aglutina fácilmente con el suero anti 0 IX y como carece del Vi no es influenciado por el anticuerpo. 2º La forma W V con un estado de equilibrio, entre estos dos antígenos, de mediana virulencia, aglutinable tanto por el suero anti IX como por el anti Vi, y por fin, 3º La forma V, con predominio marcado del antígeno, Vi, muy virulento y de alto poder vacunante, solamente aglutinable por el anticuerpo Vi. Como es lógico todas esas formas cuando con ciliadas son influenciadas por el suero anticiliar d, anticuerpo fácilmente eliminable lo que permite estudiar aisladamente los otros.
Por el alto poder virulento de las cepas conteniendo la parcela Vi, FÉLIX y sus colaboradores descubridores de ella, sostuvieron que ese elemento era el antígeno de la virulencia, pero investigaciones ulteriores no les dieron razón, especialmente por encontrarse esa parcela en algunas colonias, R apatógenas demostrándonos que su sola presencia no concede patogenidad a los gérmenes.
Por otra parte se le ha encontrado en otras Salmonellas, en las cuales no desempeña papel en la agresividad. Eso sí, la conjunción de los dos antígenos IX y Vi dan mayor patogenidad a los microbios que los contienen, tanto que BOIVIN sostiene que esa combinación constituye toda la endotoxina del gérmen pues las investigaciones de este autor en colaboración con MESRABEANU, así como las de RAISTRICK y TOPLEY, como también las de HENDERSON y MORGAN han demostrado que las llamadas endotoxinas están constituidas por la totalidad de las parcelas cuya constitución química corresponde a la de un lipidoglúsico, ellas aisladas del cuerpo del microbio, poseen todas las propiedades de lo que conocíamos como endotoxinas.
De modo pues, que según la existencia e importancia de la parcela Vi los gérmenes se manifiestan más o menos virulentos y de mayor o menor acción vacunantes, antígeno que durante el paso de los microbios por los cultivos tiende a desaparecer, quedando el microbio en estado W y con el andar del tiempo este evoluciona hacia la forma R completamente apatógena.
Según mi modo de ver la evolución de este germen es más compleja. BAIL fué el primero en lanzar la idea de que los microbios patógenos, en el organismo infestado adoptan estados especiales, que él denominó animalizados, estado durante el cual ellos eliminarían substancias especiales y específicas no tóxicas que denominó agresinas, cuya función sería la de neutralizar las defensas orgánicas, permitiendo la supervivencia del germen y la infección. Agresinas que serían por lo tanto las causantes de la posibilidad de la marcha de la infección; su neutralización suprimiría la defensa del
germen y por ende facilitaría su eliminación y la sanación del organismo. Según su modo de ver un suero antiagresínico impediría la infección.
En el carbunclo BAIL y sus colaboradores han demostrado que con las agresinas de dicho bacilo, conseguidas al filtrar el edema carbuncloso, se puede vacunar a los animales. Este concepto fué rudamente. combatido y dominado el ambiente bacteriológico por la importancia exclusiva de los anticuerpos, como portadores únicos de la inmunidad, fué bien pronto abandonado y quedó reducido al estado de una curiosidad. Sin embargo, con el tiempo se han ido acumulando una serie de experiencias y datos que no encuadran dentro de la doctrina que pretende ver en los anticuerpos toda la explicación de los fenómenos de inmunidad, muy cierta sin duda para las antitoxinas, pero no así para el proceso inmunizatorio antimicrobiano, en el cual intervienen la vida y los poderes de acomodación de los microbios patógenos, los que, al introducirse en un organismo, se encuentran en un medio hostil ante el cual reaccionan para amoldarse a él y poder seguir viviendo, lo que solo es factible mediante modificaciones de orden biológico que traen a veces modificaciones morfológicas, como sucede en el carbunclo que, en él organismo se rodea de una cápsula, la que no es sino testigo de una profunda alteración biológica, pues el microbio en este estado deja de ser fagocitado por los leucocitos del invadido como lo demostraron las experiencias de GRABER y FUTAKI, a pesar de la integridad de los fagocitos. En el curso de mis investigaciones me he convencido que con el bacilo de EBERTH pasó algo semejante, El microbio en el organismo adquiere propiedades nuevas que se manifiestan y comprueban fácilmente in vitro por su poder intenso de neutralización de los sueros específicos, muy superior a la de los microbios comunes de cultivo, aún de los que se encuentran en estado V.
El punto de partida de estas investigaciones fué la repetición de una experiencia muy sugestiva de FÉLIX y de sus colaboradores. Ellos demostraron que un suero antitífico con anticuerpos, anti IX y anti V, además del anti H, si se le agotaba por un microbio, sin poseer la parcela Vi, que por lo tanto dejaba en el suero solamente al anticuerpo Vi, seguía siendo preventivo, con tal que se le inyectase horas antes de la infección, aún utilizando vías diferentes para la introducción de estos elementos; con lo cual creían demostrar la importancia del antígeno Vi y de su anticuerpo. La experiencia ideada es perfectamente exacta y venía a corroborar otras antiguas y completamente olvidadas, como las de WEIL y BRAUN, así como a las de SPAET, de BAIL Y PETERSEN, como las de MATSUMETE, experiencias todas que como no encuadraban en la doctrina reinante, del anticuerpo como elemento esencial y preponderante, sino exclusivo para la explicación de la inmunidad, quedaron olvidadas. Las de FÉLIX y sus colaboradores llamaron la atención porque se basaban en la existencia y demostración de un nuevo antígeno, cuya existencia fué prontamente confirmado.
La repetición de la experiencia me demostró su exactitud, pero el análisis de ella me convenció que no era exacta la interpretación y me llevó a la convicción dela presencia de elementos o antígenos para llamarlos con la terminología corriente que el microbio produce al adaptarse a vivir en el medio interno hostil del huésped. Elementos que se ponen de manifiesto por el análisis del agotamiento de los sueros por los microbios en diferente estado de constitución antigénica y por lo tanto química. De esta manera he llegado a demostrar que el bacilo recientemente aislado del organismo en su primer pasaje por nuestros medios de cultivo que denomina de primer cultivo posee propiedades culturales y biológicas diferentes de los comunes aún de los que se encuentran en estado V, propiedades que se encuentran en su máximo de integridad en los microbios que se encuentran en el organismo infestado, y que por eso llamo de infección, propiedades que en el curso de los ,cultivos ulteriores en nuestros medios, se van perdiendo paulatinamente.
Corroboran este modo de ver las características diferentes de los estractos de estos diversos bacilos obtenidos por el método de LANCEFIELD. Estos productos, cuyas reacciones demuestran que están constituidos especialmente por polisacaridos, precipitan además fácilmente con débiles soluciones de sulfato de cobre. Estos precipitados son insolubles en agua destilada, pero los conseguidos de los extractos de primer cultivo se disuelven fácilmente en débil solución alcalina, mientras que los de 901 H o de Ty2, encontrándose éste en estado V, solo lo hacen en medio ácido. Por otra parte estos extractos, así como sus precipitados redisueltos, reaccionan diferentemente con los sueros específicos. Así, tratados los extractos; con suero antitifoideo Nº 262, conseguido mediante inyecciones endovenosas a conejos de emulsiones de cultivo en agar de primer pasaje, después de extraído del animal, los de primer cultivo, precipitan ya en la cantidad de 0,000012 grm., con el producto purificado aún en proporciones menores, pues es sensible la cantidad de 0,000003 grm. En cambio el extracto crudo de T y 2 solo precipita en la cantidad de 0,00005 y el producto purificado en la de 0.000.025 grm. Vale decir que los productos de primer cultivo son respectivamente cuatro y ocho veces más activos que los de Ty2. Datos todos que demuestran la diferente constitución de los microbios de primer cultivo, recientemente sólidos de organismo, del de los otros, aún de los en estado Vi, quienes en el curso de los pasajes in vitro han ido perdiendo propiedades que solo se adquieren y conservan al reaccionar en el conjunto hostil del medio interno.
Por otra parte el aspecto de las colonias, en agar, del primer cultivo difieren de las comunes porque son más opacas, más espesas, blanquesinas, brillantes, tan diferentes, que a primera vista, sin estar prevenido, parecen que no fuesen colonias del bacilo de EBERTH. Los microbios de estos cultivos se aglutinan difícilmente por los sueros obtenidos con los gérmenes comunes y cuya acción se debe casi especialmente al anticuerpo H, porque ellos no se aglutinan por el anti 0 IX, lo hacen débilmente por el anti Vi y a veces solamente por los sueros conseguidos por inyección de ellos mismos, endovenosamente al conejo. Como lo he manifestado las diferencias fundamentales se manifiestan especialmente en el diferente poder de neutralización para con el suero específico, de estos diversos gérmenes, corno lo veremos de inmediato en el estudio de la acción preventiva.
Acción preventiva. Como ejemplo de la diferente acción neutralizadora de los bacilos de la tifoidea, según el estado constitucional en que se encuentren, sobre los sueros específicos ofrecemos el estudio de: los sueros 538 y el 539, obtenido el primero por inyecciones endovenosas de bacilos de primer cultivo en conejo y el segundo por introducción por la misma vía en animal de la misma especie, de bacilos de infección, conseguidos mediante emulsiones de cerebro de conejillo de Indias, infestado cerebralmente con bacilos de EBERTH, éstos pululan ampliamente en el sitio y en los alrededores del foco de la infección. (Véase Cuadro Nº 1)

El agotamiento del suero, se efectúa diluyendo a este al décimo y emulsionando en él a los bacilos cultivados en agar. La mezcla se mantiene a 37º dos horas y luego en la heladera hasta 24 horas. Se sedimenta y en el líquido claro se mide el poder aglutinante para el microbio agotador. Si existe aglutinina aún, se agota de nuevo y tantas veces como sea necesario para que la aglutinación del suero sea negativa. Para con los bacilos de infección se procede en manera parecida, emulsionando cerebros infestados de conejillo de Indias, hasta reacción negativa para cultivo, en estado V.
Los valores preventivos son los términos medios de 30 experiencias.
La ausencia del anti Vi ha sido investigada con un bacilo de segundo pasaje de cultivo de una cepa de Salmonella typhi, la que en este estado no es aglutinado por ninguno de estos sueros agotados por el bacilo 901 H, en cambio es influenciado ampliamente por otro suero anti Vi y no es aglutinado por suero anti 0 IX.
La lectura de este cuadro es altamente interesante. Demuestra que la acción preventiva no guarda relación ninguna con el poder aglutinante, que el suero agotado por el bacilo 901 H y sin anti Vi es diez veces más activo que el suero intacto, por lo tanto debe intervenir para ello otro factor, pues no teniendo ni anticuerpo 0 ni anti Vi es altamente preventivo. Además hace suponer que la ausencia de los elementos anti 0 y anti H favorecen la acción del suero así tratado. Para dilucidar este punto se absorve un suero antitifoideo con la Salmonella gallinarum que contiene el mismo antígeno IX de EBERTH, sin parcela ciliar. De modo que solo elimina el anti 0. Pues bien un suero así tratado, agotado por el bacilo mencionado conserva su poder preventivo, al mismo nivel que el intacto, lo mismo acontece agotando el suero con la Salmonella enterititis. Demuestran estos hechos un fenómeno altamente interesante, no conocido hasta hoy y que se ha repetido con todos los numerosos sueros experimentales que he estudiado. Todos agotados por .,el 901.H se han mostrado tan activos como antes o mucho más. Este fenómeno no es de extrañar, pues la doctrina clásica nos enseña que ,estos sueros antiendotóxicos o como hoy tenemos que llamarlos antilipidoglúsicos no guardan en la neutralización la proporción de los múltiplos, como sucede con las antitoxinas y las toxinas, razón por la cual podemos neutralizar cualquier cantidad de toxina aumentando proporcionalmente la cantidad de antitoxina. En cambio aumentando la cantidad ,de endotoxina, es decir del antígeno lipidoglúsico o de bacilo muerto o vivo, llega un momento en el cual no es posible neutralizar al veneno o impedir la pululación microbiana como puede verse en el cuadro Nº 2.
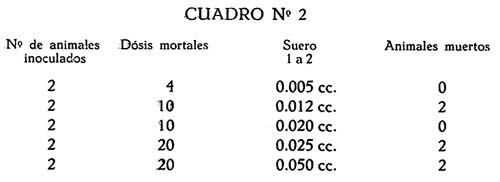
Se ve pues que, para salvar a los animalitos de 10 d.mm. es decir 2 veces y media más que la unidad tomada como base se necesitan 4 veces mayor cantidad de suero que la calculada de acuerdo con dicha base y para 5 veces la cantidad de dosis mortales, no se consigue salvar a los animalitos ni con 10 veces más suero que lo que correspondería por cálculo, a partir de la dosis efectiva para la base tomada en esta experimentación. Con el suero. La 1 se obtiene idéntico resultado.
Esta favorable eliminación del anti 0 en sueros desprovistos de anti Vi obliga a considerar y aceptar la presencia de otro factor causante de esta acción preventiva. Este factor es al que he denominado, para no prejuzgar nada X, supongo que nace a expensas de la reacción del organismo, ante los nuevos elementos bacterianos, formados por los gérmenes patógenos al adaptarse al organismo y a sus defensas, elementos que las neutralizan y permiten al microorganismo seguir viviendo y prosperando en el medio hostil que le rodea. Si esta premisa es exacta el agotamiento del suero con los bacilos de primer cultivo que son los que más cercanamente se encuentran de los llamados de infección, deberán eliminar esos elementos para los cuales el 901 H no es apto, por haber perdido en el transcurso de los cultivos la contestura de los de infección. Es justamente lo que pasa, vemos por la lectura del cuadro que ambos sueros agotados por el bacilo de primer cultivo, son mucho menos activos que los tratados por el 901 H, tienen solamente una acción igual a la del suero intacto; pero a pesar, de la absorción siguen activos y preventivos, aunque sin ninguna manifestación in vitro, elementos que por lo tanto no son captables en el tubo de ensayo y que los bacilos comunes no son capaces de eliminar. Luego los microorganismos de primer cultivo son biológicamente diferentes de los comunes, tienen un poder neutralizante mayor, superior al de los otros, deben guardar en parte la constitución de los de infección. Si esta idea es exacta, estos tendrán propiedades neutralizantes aún superiores a los de primer cultivo y efectivamente es lo que sucede como puede verse en el cuadro. Uno de los sueros queda totalmente inactivo y el otro actúa con la cantidad en la cual suelen ser activos los sueros normales, vale decir quedan eliminados todos los elementos específicos. Vemos por lo tanto que en los bacilos de infección es tal el poder de absorción que dejan inermes a los sueros específicos, los de primer cultivo conservan parte de ese estado pues eliminan elementos que permanecen, aún después de la absorción por los microbios comunes. Estas experiencias nos demuestran claramente que por el hecho de la adaptación al medio interno, al transformarse en microbios de infección los bacilos de EBERTH, adquieren propiedades biológicas nuevas y son seguramente éstas las que originan los elementos específicos que en el suero no pueden ser, eliminados por el 901 H elementos que he denominado X; elementos totalmente eliminables por el microbio de infección y parcialmente por los de primer cultivo y que podemos llamar para facilitar su examen Xa y Xb.
Puede objetarse a estas experiencias que para ser completa la demostración falta en los sueros examinados, la cuota anti Vi, pero aún con la presencia de ésta los fenómenos son similares como lo demuestra la lectura de los cuadros Nos. 3 y 4.


El bacilo de primer cultivo se consigue de la siguiente manera: se mezcla I u II gotas de sangre del corazón de un ratón blanco, recientemente muerto por infección peritoneal de bacilo de EBERTH GRA, altamente virulento (cuya dosis mortal es de 0.005 cc. de cultivo en caldo de 20 horas de edad, mantenido a 37ºC) con 5 cc. de caldo. De esta mezcla se agrega I o II gotas para cada tubo de agar, o alrededor de 2 o 3 cc. para una botella tipo ginebra con agar, cuando se quiere emplear mucha cantidad de suero para el agotamiento.
Es un suero muy activo, con una fuerte cuota anti Vi, pero aún con, la presencia ésta, los fenómenos son similares. La cuota anti Vi es alta, pues para el bacilo T y 2 mantenido activamente en estado Vi es de1/5000.
Vemos en este caso también la mayor inaglutinabilidad del bacilo, de primer cultivo, tanto, para con el suero puro, como para el que se encuentra en estado Vi. Constatamos también que el suero agotado por bacilo de primer cultivo pierde totalmente su poder aglutinante, en cambio el que ha sido tratado con el microbio Ty2 conserva ligeras cuotas para el 901 H y el 901 O.
Este suero intacto tiene un poder preventivo igual a 0/0001 cc. no pierde su valor por el agotamiento con el bacilo 901.H, vale decir que otra vez la eliminación del anti O y del anti H, no le privan al suero, para nada de su poder preventivo. En este caso puede ron razón aducirse que la acción remanente del suero se debe a la Presencia del anticuerpo Vi y vemos efectivamente que el suero 862 agotado por el Ty2: en estado Vi, pierde parte de su valor, es cinco veces menos activo que el puro, o el tratado por el 901.H, es decir que indudablemente se nota la influencia evidente de la eliminación del anti Vi; pero a pesar de eso ,no queda inerte es activo, demostrándonos que a pesar de la eliminación de los elementos anti O, anti Vi, anti H el suero actúa. Luego esta experiencia comprueba que deben existir otros factores actuantes en este caso, similar a los del suero 538, antes mencionado, vale decir el llamado factor X. En este caso la absorción por primer cultivo debe ,dejar al suero con menor acción y es ciertamente lo que pasa demostrándonos la verdad de la tesis sostenida. En el caso presente no se siguió más adelante el agotamiento porque se analizaba especialmente la ,acción curativa, la que como luego veremos se pierde totalmente mediante la absorción con el bacilo de primer cultivo.
Esta acción de los factores X no se ejercen al igual de los otros directamente sobre los bacilos y solo se efectúa después de un período de incubación de algunas horas y se manifiesta como una verdadera inmunización pasiva, porque ella desaparece al cabo de más o menos tres .semanas. Cuadro Nº 4.
Se inyectan subcutáneamente a los animalitos (ratones), con 0.002 cc. de suero Lanz agotado por bacilo 901.H y con diferentes intervalos de tiempo se infestan en el peritoneo con cuatro dosis mortales mínimos de primer cultivo en caldo de 24 horas de edad de la cepa Gray, que se encuentra en estado Vi.
Se demuestra que la acción no es directa sobre los bacilos, con la mezcla de los microbios con el suero e inyectándolos después. Los anticuerpos comunes en este caso impresionan de tal manera al bacilo lo apronizan, de modo que ellos son fácilmente fagocitados, al ser inyectados dominándose la infección. Cuadro Nº 5.

El dispositivo experimental fué el siguiente: se mezclaron 4 dosis mortales mínimas con diferentes cantidades de suero y se mantuvieron en contacto cinco minutos a la temperatura ordinaria y luego se inocularon en el peritoneo de lauchas. En el cuadro siguiente se dan los valores medios obtenidos en doce experiencias:
Vemos que el suero ha mezclado en la cantidad de 0.005 cc. con cuatro dosis mortales de primer cultivo e inyectado peritonealmente a la laucha, es un procedimiento que salva al animalito. En cambio el agotado por el bacilo 901 H ni aún en la cantidad de 0.01 mezclado con cuatro dósis mortales e inyectado peritonealmente es capaz de salvar a los ratones inoculados. Sin embargo en la experiencia clásica de acción preventiva, inyectando los sueros 24 horas antes en el tejido subcutáneo y los bacilos en el peritoneo la dósis del suero puro activo es de 0.005 cc. ,siendo la misma para el agotado por bacilo 901.H, es decir que el agotado se muestra tan activo como el intacto. En cambio no es capaz de impresionar directamente a los bacilos. Parece por lo tanto que la acción del suero en función X se ejerce merced a la intervención del organismo inyectado, o que este suero se reactiva de alguna manera en él. Los fenómenos observados en el peritoneo de los animalitos vacunados pasivamente con los sueros agotados por bacilos 901.H, son completamente diferentes de la de aquellos que se manifiestan en los inyectados con sueros intacto. En este caso se ve una franca reacción fagocitaria, una intensa reacción polinuclear, fenómenos que dominan rápidamente la situación. En cambio en los inyectados con suero en función X se vé también una acumulación, de leucocitos, pero en la mayoría son monoculeares, observándose una fagocitosis sumamente escasa, pero se notan modificaciones en los microorganismos, los que se encuentran en pequeñas agrupaciones o formando cadenas de hasta diez elementos, constituyendo filamentos sin división aparente, como si el proceso de la multiplicación estuviese detenido por alguna causa integrante del medio en el cual se desarrolla el microbio. Por otra parte se ve en estos animales tratados con suero en función X que los bacilos inyectados en el peritoneo pasan rápidamente a la sangre, en la cuál se encuentran ya a los cinco minutos y permanecen ahí hasta las 72 horas después de la infección. En los órganos especialmente en el bazo se nota una intensa reacción linfocitaria con intensa congestión y sin embargo a pesar de estos cambios el animal soporta fácilmente este estado sin trastornos aparentes a pesar de la permanencia de los bacilos en la sangre y de las profundas alteraciones de sus órganos; los microbios van desapareciendo paulatinamente mediante un rnecanismo que aún no he podido establecer. Basándose en la imagen peritoneal puede pensarse en una acción bacterio-estática, que impidiese al microorganismo pueular libremente y transformarse en microbio de infección, pero es lógico que este concepto por el momento solo puede exponerse hipotéticamente. Por el estudio dela acción curativa de los sueros en función, X veremos que se refuerza ese modo de ver.
Acción curativa. Para estudiarla hemos empleado el suero 682 ya mencionado y su acción ha sido medida inyectando en el peritoneo de ratones blancos dos dosis mortales mínimas de cultive, en caldo de primer cultivo, mantenido durante 20 horas a 37º. Dos horas después de la infección, momento en el cual pululan los microbios en el peritoneo, y que ya se encuentran en la sangre, se les inyecta en la misma cavidad las diluciones del suero a dosar. Véase el Cuadro Nº 6.

Estas cifras han sido obtenidas cuando la dosis mencionada salvaba a los 5 animales en experimentación y que los testigos inoculados morían todos.
El estudio de la acción terapéutica de este suero nos demuestra también que para su eficacia no es necesaria la intervención de los anticuerpos O y H, pues el suero desprovisto de ellos mediante la absorción por el bacilo 901.H, no solo es eficaz, sino que lo es tanto o más que el intacto.
Otra vez pudiera aducirse en este caso que el anticuerpo Vi, es el causante de esta acción, pues él queda remanente en el suero así tratado. Nos demuestra lo contrario la actividad del suero agotado por el bacilo Ty2 en estado Vi el cual elimina los anticuerpos O, Vi, H, a pesar de lo cual, él sigue altamente efectivo, con un valor de 0.0005 cc.; es verdad que resulta con un valor 16 veces menor que el agotado por el 901.H, cuyo límite de acción es de 0.00003, demostrándonos, sin lugar a dudas, la existencia de la acción anti Vi. Pero hay que tener presente que su eliminación mediante el agotamiento con el Ty2 no deja inerte al suero, este queda bien activo lo que demuestra evidentemente que esa actividad se debe a otros factores que el anti Vi y que el bacilo mencionado no ha podido eliminar. Elementos que son casi totalmente absorbidos por los bacilos de primer cultivo, pues el suero agotado por este microbio, ni aún en la cantidad de 0.01 cc. es capaz de actuar terapéuticamente. Es decir que el suero tratado por Ty2 es por lo menos 20 veces más activo, que el absorbido por bacilos de primer cultivo. Esta experiencia pone de manifiesto que el bacilo de primer cultivo elimina del suero los anticuerpos O Vi, H, y que aquellos que permanecen en él y lo dejan activo, a pesar de haber sido agotado por, microbios en estado V. Elementos éstos que ni el 901.H. ni el Ty2, son capaces de eliminarlos, por carecer de actividad para ello, la que posee el bacilo de primer cultivo, fenómeno que demuestra la diferencia biológica entre ellos. Son justamente esos cuerpos que escapan a la acción de los microbios comunes, los que a mi juicio, como lo he manifestado, constituyen los factores esenciales de la función X. Nacidos ellos por la excitación provocada en el organismo por los antígenos fundamentales o antígenos de adaptación, que he llamado antígeno fundamental, formados por los bacilos al transformarse en microbio de infección, resistente a las defensas orgánicas, elementos que se conservan en gran cantidad en los bacilos de primer cultivo.
La existencia de estos elementos X se manifiestan también por la diferente manera de acción sobre los bacilos, lo que se observa perfectamente bien estudiando la evolución de las manifestaciones que tienen. lugar en el peritoneo de los animales infestados. Esta investigación nos presenta diferentes cuadros según los anticuerpos que actúan. En los animales tratados con el suero intacto con todos sus anticuerpos se nota en la cavidad peritoneal una activa fagocitosis, con un máximo a las dos horas de efectuada la inyección terapéutica, para luego ir disminuyendo lenta y paulatinamente y llegar a O a las seis horas, momento en el cual no se pueden poner en evidencia los microbios mediante el examen microscópico. En los ratones tratados con el suero agotado por el bacilo 901.H, es decir conteniendo solamente los anticuerpos Vi y X, la curva fagocitaria es semejante, pero tiene un segundo repunte manifiesto a las cuatro horas y media después de la inyección terapéutica para caer luego lentamente a las seis y media horas coincidiendo con la desaparición bacterioscópica de los bacilos. En cambio en los tratados con el suero agotado por el bacilo Ty2 es decir conteniendo solamente elementos X el cuadro peritoneal es muy diferente. Se encuentra muy poca fagocitosis, la que desaparece a las cuatro horas después de la inyección tratante, momento en el cual hay un repunte bacilar en el peritoneo, estos microbios van luego desapareciendo poco a poco hasta que a las siete horas no se les puede ver más por el examen microscópico. De modo pues que mediante un procedimiento en el cual no interviene la fagocitosis los bacilos desaparecen y son destruídos entre las cuatro y las siete horas después de la introducción del suero. Fenómeno, que en parte, se cubre con los observados en el tratamiento preventivo con suero en estado X, en cuyo caso tampoco se nota la fagocitosis como elemento preponderante de eliminación bacilar, constataciones experimentales que obligan a pensar en otro medio de acción antibacteriana. Coinciden también ambas experimentaciones en demostrar la existencia de factores microbicidas en los sueros agotados por el bacilo Ty2 y cuya acción es diferente de aquella que caracteriza a los anticuerpos comunes, elementos que solo pueden ser eliminados por el agotamiento del suero mediante bacilos de primer cultivo.
En conjunto se nota que en los animales inyectados con suero agotado por el bacilo Ty2 es decir con función X una acción bactericida diferente de la ejercida por el suero íntegro se nota un gran predominio de lisis celular y desaparición de los microbios fuera de las células, con alteraciones en su morfología y multiplicación y por un mecanismo por ahora no conocido, hipotéticamente bacteriostático y que acantonan al microbio, que permanece vivo sin manifestarse, sin provocar lesiones, durante tres o cuatro días en que puede constatarse su presencia culturalmente y en tan escasa cantidad que no se le puede poner en evidencia, ni con el más cuidadoso examen microscópico.
En el esquema siguiente podemos resumir el conjunto de los resultados experimentales expuestos en este trabajo.
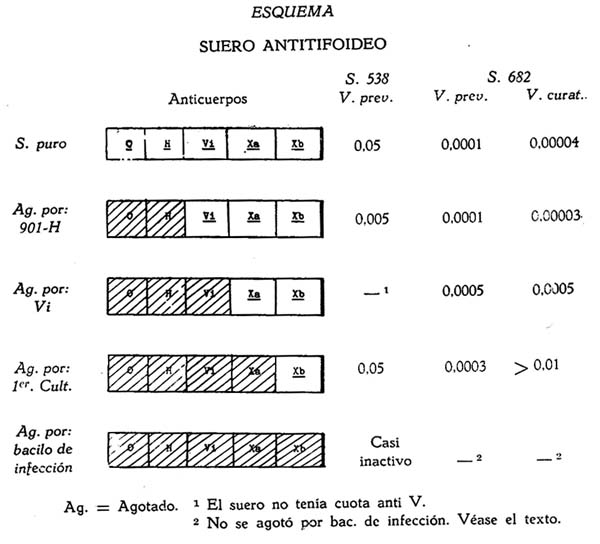
Comentarios al esquema:
El suero Nº 538, obtenido en mulo, por inyecciones endovenosas de bacilos de primer cultivo, está desprovista de acción Vi y sin embargo agotado por bacilos 901 H, a pesar de perder todo en poder aglutinante, pues no actúa ni sobre microbios en estado Vi, ni sobre los de primer cultivo, lejos de quedar inactivo por la absorción de los anticuerpos O1X y H y por no tener anti Vi, es sin embargo 10 veces más activo que el puro. En este caso solo puede pensarse en la existencia de otros elementos, remanentes en el suero y a los cuales debe su actividad, son justamente a los que llamo función X.
Este suero agotado por microbios de primer cultivo, no pierde totalmente su valor, se eliminan mediante este tratamiento los anti-O y anti-H y como tiene menor acción que el tratado por 901 H debe haber absorbido parte de los elementos respetados por éste y solamente parte porque conserva algo de su actividad, la que en el caso particular que exponemos llega a ser igual a la del suero puro. Como los bacilos de infección anulan toda la actividad del suero, quiere decir que la porción de elementos activos dejados intactos por los microbios de primer cultivo son eliminados por los bacilos de infección. Por este hecho supongo, y con razón que para los elementos que constituyen la función X debe admitirse que están formados por dos partes: una eliminada por los bacilos de primer cultivo y los de infección y la otra solamente por estos últimos.
El suero 612 ha sido suficientemente comentado en el texto y sus resultados corroboran y amplían los del 538.
DEDUCCIÓN
Estas experiencias demuestran, a mi juicio, que la Salmonella typhi virulenta al penetrar en el organismo animal se modifica biológicamente de tal manera que le permite seguir viviendo en el medio interno hostil del huésped. Modificación constitucional, que tiene como respuesta del organismo infestado la producción de elementos o anticuerpos para expresarse con la nomenclatura actual y que para no prejuzgar sobre su esencia, he llamado, al conjunto función X. Elementos que no pueden ,ser eliminados del suero, sino por los microbios de infección o los de primer cultivo los que aún conservan en parte el estado constitucional de los de infección y que los comunes, aún los llamados V, no lo pueden ..hacer por haber perdido ese estado, en el curso de los cultivos, lejos de .la influencia hostil que lo provoca, faltándoles por eso la capacidad para eliminar del suero los elementos de la función X. Elementos que .actúan preventiva y curativamente, sin necesidad de los anticuerpos comunes y mediante un mecanismo diferente.
DEDUCTION
These experiences prove, in my own judgement, that the virulent Salmonella Typhia upon entering into the animal organism, modifies itself biologically in such a way as to continue living in the hostile medium afforded by the host. This constitutional modification gets an ,answer from the infected organism in the form of a production of elements, antibodies to express myself in the current nomenclature to which in order not to be prejudiced about its esence, I have given the name of function X. These elements can not be eliminated from the serum except by the infection germs or the ones of the first culture, which still have to a certain degree the constitutional state of the infection, and can not be eliminated by the common germs, even by the so called V germs, which have: lest that state throughout the cultures removed from the hostile influence that provokes it. Due to this fact fact they den't have the capacity to eliminate from the serum the elements of function X, which act in a preventive and curing way without the need of the common antibodies and by means of a different mechanism.
BIBLIOGRAFÍA
1. WHITTE : Jour. Path. Bac., t, 35, p. 77.
2. KAUFFMANN & WHITTE: Third. Int. Cong. f. Microbilogie, p. 834.
3. KAUFFMANN: Zeitschf. f. Hyg., t. 117, p. 778.
4. FÉLIX PITT: Jour. Path. Bact., t. 38, p. 409.
5. FÉLIX PITT The Lancet, t. 227, p. 186.
6. FÉLIX BATHNAGER : Brt. Jour. Exp. Pathl., t. 15, p. 346.
7. BOIVIN & MESRABEANU : Rev. d´inmunologie, t. I, p. 551; id. t. II, p. 113; id. t. III, p. 319; id. t. IV, p. 40 y 197, id. t. V, p. 86.
8. RAISTRICK & TOPLEY : Brit. Jour. Exp. Path., t. 15, p. 113.
9. HENDERSON & MORGAN : Brit. Jour. Exp. Path t. 19, p. 82.
10. BAIL: Weichhard. Jaresbericht, t. 7, p. 81 y Handbuch. der Path. Mi krorgrnidmen, 3 edc., t. II, p. 635.
11. LOEHLEIN : Gruber. u. Futaki Zentrabl, f. Baktr. Ref., t. 38, p. 901.
12. WEIL & BRAUN : Folia Serologica, t. III, p. 271.
13. SPAET: Zeitschf. f. Hig., t. 69, p. 46.
14. BAIL & PETERSEN : Zentrlbl. f. Bactr. I ori., t. 34, p. 4.
15. MATSUMOTO: Zeitschj. f Immunitf., t. 40, p, 402.
16. LANCEFIELD: Jour. Exp. Med., t. 47, p. 91.
17. BACHMANN Bol. Academia Nac. de Medicina, 1936, p. 1099; id. Junio. Agosto y Setiembre de 1937; id. Abril - Mayo y Octubre de 1944; Día Médico, t. X, Nº 31; Rev. Asoc. Med. Arg., t. LX, Nº 461 y 462; VI Congreso Nac. de Med. Soc. Bio. Arg., 1937; IV Cong. It. Path. comp., Roma 1937, t. II, p. 437; C. R. Soc. Boil., t. II, p. 1217; id. t. II, p. 345; Rev. Sud Americana de End. Inmunología, t. XX, p. 629, id. t. XXI, p. 751; id. L XXII, p. 73.

