










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.4 n.3 Lima sep. 1945
TRABAJOS ORIGINALES
Contribución a la histopatologia del distemper canino
E. Encinas1
1 Jefe del Laboratorio de Anatomía Normal y Patológica del Sistema Nervioso, de la Facultad de Medicina.
Laboratorio de Anatomía Normal y Patológica del Sistema Nervioso de la Facultad de Medicina.
RESUMEN
En un caso de distemper canino el examen anatómico del sistema nervioso ofrece cambios elementales y estructurales sistematizados, no revelados hasta ahora. El hecho de que determinadas unidades estructurales, en evidente relación con los espacios sub-aracnoideos, hayan sido preferentemente comprometidas, en contraste con otras que se han mantenido indemnes por estar fuera de la influencia de aquellos y de su liquor circulante, hace sospechar que aparte de la vía sanguínea, sea el liquor uno de los vehículos del agente infectante. Pero lo que esta influencia no explica es el cambio parcial de las células nerviosas de ciertas zonas, de las formaciones ammónicas, Fig. Nº 44, ni la alteración de los componentes celulares de una unidad arquitectural, de lámina Vª de la corteza, por ejemplo. Para esta especie de variación no cabe al presente, mas que la sospecha, a menudo comprobada, de que determinadas unidades celulares o arquitecturales, poseen cierto grado de vulnerabilidad bis a bis de los agentes agresores. Y esta opinión tan a menudo verificada en la patología humana no tiene porqué no ser extendida a los cambios en los tejidos nerviosos de animales inferiores.
El asta de Ammón humana ha sido con frecuencia estudiada en múltiples procesos patológicos, comprobándose la manera fácil con que agentes vulnerantes la modifican y lesionan, pero, que sepamos, no se han hecho estudios detenidos de ella en procesos sospechosos virulentos como en las encefalitis, ni en aquellos debidos a bacterias. En estos aspectos como en otros de las investigaciones biológicas parece que todos los recursos estuvieran dirigidos a verificar hallazgos ya realizados, o a comprobar sus variaciones, más nó a ir al encuentro de nuevos hechos de información mayor, y de provecho para el conocimiento, Asunto sería de investigar por ejemplo si en la encefalitis humanas ocurren alteraciones semejantes a las mencionadas en este trabajo (de distemper), y, si en efecto las unidades celulares o estructurales reaccionan en la medida y forma señaladas frente a cualquier otro agente alterativo. Posible es que en el caso particular de las lesiones aisladas de la lámina Vª juegue rol determinante la especial susceptibilidad de esta lámina a deficiencias en el aporte de oxígeno, ocasionadas por perturbaciones en el aprovisionamiento de sangre, portador de este gas cuestión que discutieron acertadamente BIELCHOWSKY y PALADINO, MARINESCO y otros; y como alteraciones pericapilares y endocapilares son hechos frecuentes en este proceso, no sería aventurado sostener que las lesiones aisladas de esta lámina se deban a aportes insuficientes de oxígeno por las anotadas alteraciones vasculares. Pero, y repitiendo lo que dijimos en líneas anteriores, factor este vascular, no explica el hecho anatómico del cambio celular aislado como el de las unidades mencionadas de la formación ammónica (Fig. 44). Alguna otra causal citoespecífica debe ser la sola causante de alteración tan particular. Y no es extraño que el virus del distemper, tan notable por la especificidad de las inclusiones citoplasmáticas y nucleares que provoca, sea ajeno a esta modificación.
En lo fundamental, desearíamos que nuestros hallazgos sean verificados y que la autorizada opinión de los cultores del tema se pronuncien sobre ellos.
Numerosas son las publicaciones aparecidas en el mundo sobre distemper canino, y si el interés que se ha tomado en alguno de sus aspectos, en el de su etiología por ejemplo, o en el de ciertos hechos particulares, se hubiera extendido hacia un análisis anatómico de mayor cuidado, conoceríamos hoy mejor, circunstancias orgánicas que han pasado desapercibidas, y que como las que dan materia a este trabajo habrían constituido para la autorizada experiencia de los investigadores del tema, motivos de interés vivo. En efecto, en la bibliografía sobre la materia no hay una información precisa, documentada, referente a las alteraciones del tejido nervioso en el distemper. Las publicaciones se refieren a ellas de modo muy general: se habla de alteraciones agudas de las células nerviosas, de las manifestaciones inflamatorias de los vasos, de las inclusiones citoplasmáticas e intranucleares en diversos endotelios y en células del parénquima, de las reacciones glióticas, etc., etc. Más, las respuestas especiales de los tejidos a la agresión, en este caso probablemente debida al virus, no han recibido conveniente respuesta.
De otro lado en los trabajos publicados hay continuas referencias a las semejanzas de la enfermedad con procesos parecidos humanos, con la parálisis infantil y con las encefalitis epidémicas por ejemplo, pero en adquisiciones citopatológicas nerviosas se ha considerado el tema con la atención conveniente. Deseosos estábamos por esto de desarrollarlo, de averiguar en detalle la manera xepresiva de las alteraciones anatómicas, y del modo cómo unidades tisulares o unidades estructurales pudieran estar comprometidas, como ocurre aveces en los procesos humanos. Y el resultado, por el material iconográfico con que acompañamos este trabajo, no puede ser más sugestivo. Quedan pendientes muchos puntos, y además comprobación única no es de grande valor probatorio, razones éstas que nos comprometen para posterior tarea que desarrollaremos si acaso tiempo y circunstancias han de sernos propicios.
Al doctor T. Battistini, Director del Instituto Nacional de Hígiene, a quien justo es reconocerle el haber señalado la existencia de la enfermedad del distemper, por primera vez en el Perú, deseamos significarle con esta oportunidad nuestra mayor gratitud por las facilidades que nos ha ,proporcionado en la elaboración y publicación de este estudio.
El distemper canino es una enfermedad infecciosa, contagiosa, predominantemente estacional, que compromete a los hurones, armiños, comadrejas, y los perros de corta edad. Son sus síntomas : catarro, trastornos respiratorios, gastrointestinales y nerviosos. Los ingleses y norteamericanos lo llaman canine distemper, canine plague, wheel plague, canine glanders, catarral fever; los alemanes : Hundestaupe, Staupe der Hunde, Hunderkrankheít; los franceses : maladie des chiens o maladie des jeunes chiens; los italianos : cimurro, moccio canino.
Como el sarampión y la tos convulsiva en el hombre, pocos son los animales que escapan a la enfermedad, la que, por otra parte parece conferir inmunidad. Se la encuentra en casi todos los países, y en cualquiera estación del año, aunque la primavera y el verano son las épocas mas favorables para su desarrollo. Se la creyó patrimonio exclusivo de los perros de corta edad, pero LECLAINCHE comprobó que al lado del factor edad, la raza juega papel importante en su desarrollo, siendo los animales de cepa fina, los particularmente susceptibles, y de mayor resistencia, los perros del campo, mestizos o vagabundos.
Parece que fue conocida desde los tiempos de Aristóteles. LAOSSON considera que la epizootia canina que arrasó Bohemia durante el año de 1028 fué distemper canino. Es opinión corriente que fué importada de América a Europa a mediados del siglo XVIII, primero a España de donde se propagó rápidamente. JENNER en 1809 al establecer la naturaleza infecciosa de la enfermedad, fue el primero en diferenciarla de la rabia, y el primero también en comprobar que no era trasmitible al hombre este dominio no se ha penetrado lo suficiente, ni a la luz de las nuevas. D´YOUVILLE a mediados del siglo XVIII, al sospechar su contagiosidad dió indicaciones precisas para evitar su propagación en establos y perrerías. Entre 1818 y 1852 muchas comunicaciones fueron escritas concernientes a su naturaleza contagiosa y a la posibilidad de su trasmisión al hombre. WALDINGER, VON GEMMERN Y MECKE, DELABERREBLAIN, FUNKF y VEITH sostenían que la enfermedad era trasmisible al hombre. RENNER y KERLIE afirmaban lo contrario. La enfermedad fué considerada contagiosa por ROLL y HERTWIG, no contagiosa por TRANSBOT, VENUTA, SEMMER, TRASTOWO y KRAJEWSKI. Para muchos, tenía considerables semejanzas con algunas enfermedades del hombre sobre todo por sus manifestaciones broncopulmonares, intestinales y nerviosas; y en cuanto a las pústulas que aparecen en determinadas oportunidades en la piel del abdomen y en la cara interna de los muslos e ingles, mucho se habló a cerca de sus posibles semejanzas con las pústulas de la vacuna o de la viruela, y hasta se inoculó la serosidad de las mismas con fines de curación o preventivos, con resultado negativo.
Hay grande semejanza entre el distemper canino y la encefalitis de los zorros y aunque en pasadas épocas se clasificaba esta enfermedad en el grupo del distemper canino, parece tratarse de proceso distinto por las circunstancias epidemiológicas y anatómicas que mencionaremos después. De otro lado REUTER y otros, sostuvieron que el distemper canino y el distemper equino reconocían la misma etiología y que podían ser trasmitidas de un animal a otro. Las investigaciones recientes parecen confirmar este punto de vista.
SÍNTOMAS
Los signos clínicos se presenta 7 a 8 días después de la exposición al agente infectante y se traducen por fiebre que dura al principios unas 96 horas, que remite, y se pone a nivel normal, para reaparecer 11 a 12 días después. Acompañan a la fiebre catarro, en oportunidades tan marcado que conduce a la producción de abundante secreción nasal purulenta; bronquitis, que degenera en oportunidades en bronconeumonias; diarreas tan intensas que pueden conducir al animal a graves estados de desnutrición, y, manifestaciones nerviosas de todo orden, desde las simples mioclonias y movimientos espontáneos semejantes a los de la corea, por lo que se la llama también corea canina, hasta parálisis de grandes segmentos del cuerpo, amén de las perturbaciones del comportamiento que se traducen por estados depresivos o de irritabilidad. En oportunidades el animal sufre violentamente la pérdida del conocimiento y de la actividad y es presa de ataques contracturales segmentarios o generales, incluyendo las violentas contracturas mandibulares, y salivación abundante, provocada por estos últimos movimientos. Las convulsiones son en ocasiones tan frecuentes y acentuadas que dan lugar al desarrollo de verdaderos cuadros epilépticos permanentes, después de los cuales el animal permanece privado de movimiento, como adormecido.
La escasa significación de los síntomas puede dar lugar a que en oportunidades la enfermedad pase desapercibida, pero sino es la forma nerviosa, definitiva en sus manifestaciones y mortal, la que se desarrolla, el proceso sigue el cuadro correspondiente al sistema predominantemente comprometido, y así se vé formas broncopulmonares, en gran parte causadas, según algunos autores, por gérmenes de infección secundaria, como los estreptococos, y el mismo B. bronchisepticus, al que como veremos después se le atribuye el origen de la enfermedad; intestinales, en las que el síntoma dominante, la diarrea, conduce a graves condiciones, y, tegumentarias. Más, esta. distribución es arbitraria, y resultado sólo del predominio sintomatológico. La infección es general como todas las debidas a virus; los líquidos y la sangre se les encuentra infectantes una vez que el proceso acusa su más alta manifestación sobre todo febril. La creación de las entidades particulares es artificial por esto, y como lo afirman los conocedores de la enfermedad, fruto exclusivo de la mengua general orgánica y del papel especial de los gérmenes de infección secundaria.
ETIOLOGÍA
Se atribuyó la enfermedad a alimentación insuficiente o impropia, al frío, a condiciones generales que pusieran en peligro sus resistencias orgánicas, y por último a agentes de causalidad mas precisa, sea bajo la forma de organismos bacterianos o de virus. SEMMER (1875) halla un bacilo pequeño y fino en la sangre de los perros afectados por la enfermedad, pocas horas después de la muerte. KRAJEWSKI (1881) halla micrococus. RABE (1883), encuentra cocos uniformes, de pequeño tamaño, juntos o en grupos de 2 o 4, o en series de 4 a 5, teñidos en azul obscuro con el azul de metileno. MATHIS encuentra un diplococus en el fluido de los tejidos, del esputo, y de las pústulas, que cultiva en medios neutrales o ligeramente alcalinos. MARCONI Y MELONI (1888) encuentra cocos similares a los estafilococos. JAQUET y LEGRAIN ( 1890) encuentra en las pústulas micrococos móviles. MILLAIS (1890) halla bacilos largos que liquan la gelatina, y micrococus a los que les atribuye el desarrollo de los cuadros pulmonares. Según este autor los dos agentes, los bacilos y los micrococus serían los productores del pneumo-distemper. SCHANTYE (1892) divide el distemper en otros tres procesos, atribuyendo a cada uno de ellos distinto organismo. JENSEN (1896) cree que la pneumonia y el distemper son producidos por un estreptococo. VALERIO (1896) logra cultivar en agar bacilos positivos de extremos redondeados. gasógenos, de material procedente de los senos frontales, pulmones, medula espina], y del cerebro. PETROPAWLOWSKY (1897) halla bacilos semejantes a los del colon. TATY y JACQUIN (1898) encuentran diplococos en el sistema nervioso central y los sindica como causantes del distemper. COPEMAN (1900) en frotis de material proveniente, de cultivos en caldo, logra observar cocobacilos, a veces en cadena, no tingibles por el Gram, que crecen en el agar a 36º C. en colonias grises, semitrasparentes, circulares, de bordes al principio irregulares. Su crecimiento es lento en el serum; la leche no es coagulada; la gelatina se liqua. LIGNIERS Y PHISALIX (1900, y 1901 ) encuentra bacilos largos; Gram-negativos, movibles, que toman una forma coco-bacilar en los cobayos. CADIOT y BRETON describen la bronco-neumonia de la enfermedad como proceso enteramente distinto. CARRÉ (1905) refiere haber comprobado la existencia de un virus filtrable proveniente de las descargas serosas. Las serosidades obtenidas de la enfermedad acusan un notable poder patogénico y logra trasmitir la enfermedad mediante la inoculación de las provenientes de las cavidades nasal, bronquial, pleural, pericádica y peritoneal, y de la sangre sobre todo, cuando se las torna en momentos culminantes de la ascención térmica. inocula los productos orgánicos mencionados después de haberlos hecho pasar por el filtro Chamberlain por 3 a 4 días, a perros jóvenes, y obtiene la infección. Este hecho establecido por CARRÉ en 1905 fue posteriormente comprobado por el mismo en los años 1924 y 1926, y por una serie de investigadores, entre ellos LÜHRS, ROBIN y VECHIU y sobre todo por DUNKIN y LAIDLAW, todo lo que parece demostrar que el agente patógeno es un ultravirus. La enfermedad pudo ser trasmitida a los perros y gatos. Para estos últimos, LAOSSEN y recientemente A. CHIRILA, ROBIN Y VECHIU se sirvieron de líquido céfalo-raquídeo, tomados de los casos de forma nerviosa aguda. VECHIU logró reproducir la infección inoculando orina filtrada, y líquido céfalo-raquideo y emulsiones de masa cerebral de animales que habían sufrido la infección de forma crónica. LÜHRS (1926) obtuvo inoculaciones positivas con el líquido de las pústulas, DUNKIN y LAIDLAW con sangre de pulpa esplénica inyectadas subcutáneamente. La inoculación es en un gran porcentaje positiva, valiéndose de sangre y de emulsiones de substancia nerviosa y esplénica. HEWER (1906) encuentra bacilos pequeños, gruesos, no movibles, Gram-negativos que se cultivan muy bien en el agar, enturbian el caldo y dejan un sedimento granular; la gelatina no se liqua.
FERRY le dió el nombre de B. bronchisepticus. TORREY y RAHE (1913), después de minucioso estudio afirman que habían logrado reproducir la enfermedad en animales susceptibles a la infección medinate la inoculación de cultivos puros del gérmen de FERRY y M. GOWMAN. A. LOCKCHART (1926) demuestra en el examen de los líquidos de las cavidades internas del cerebro y de las menínges, la existencia de organismos bacterianos peculiares de las clases del diplococo y estreptococo, que inyectados a animales susceptibles reproducían la enfermedad. Se trata según este investigador de estreptococos atípicos capaces de desarrollarse tanto en la sangre circulante como en los fluidos del cerebro y de la médula. Sobre la base de sus comprobaciones y con cultivos de estreptococos polimorfos prepara una bacterina a la cual le atribuye un efecto tan beneficioso que logra hacer descender la mortalidad de un 15 % a un 1/2 a 1%.
El efecto es mayor si la bacterina es inyectada inmediatamente después de las primeras manifestaciones de la enfermedad. P. M. GOWMAN ( 1910) sobre la base de algunas necropsias en perros y gatos y en vista de no hallar excrecencias tisulares, sospecha que la forma paralítica de la enfermedad debe ser debida a la presencia de una toxina.. De dos gatos inoculados intraperitonealmente con una emulsión de cultivos muertos por calentamiento a 560 por media hora, uno de ellos murió 3 horas después con fuertes convulsiones en las extremidades y en las mandíbulas, indicando la presencia de una fuerte neurotoxina. Ninguno de los dos gatos inoculados presentó lesiones peritoneales. Los cultivos que emplea los obtuvo de secreciones provenientes de la base del cerebro de un perro muerto con signos coreicos. Posible es que este hecho, el de la presencia del líquido virulento en tal sitio, se deba, a las interrelaciones comunicativas del oído medio con la cavidad craneal, y a que el virus puede haber emigrado desde el cavuni de la boca, o de las vías aéreas superiores a la región auditiva interna y de aquí hasta la cavidad cefálica. No es extraño que sembríos semejantes se efectúen en el intestino, desde que las manifestaciones diarreicas son frecuentes, a pesar de no haber encontrado el autor lesiones en este órgano. La presencia de esa toxina, tan virulenta, explicaría las manifestaciones paralíticas de los animales infectados natural o artificialmente, sea por compromiso de centros nerviosos vitales, o por lesiones que afecten la médula espinal, o el corazón en su sistema ganglionar nervioso, o en su tejido muscular. De todos los animales examinados el foco inicial parece ser el tractus superior de las vías respiratorias, especialmente la nariz, de cuyas mucosas obtuvo las substancias que sirvieron para los cultivos y para las inoculaciones experimentales. A. S. SCHILINGMAN en comunicación presentada a la American Veterinary Association en Kansas City, Mo., en Agosto 25-28 de 1931, establece que de 100 perros atacados con distemper canino el 81 % muestran la presencia del B. bronchisepticus, un 9 % estreptococos hemolíticos, un 6 % estafilococos albus; en una proporción mucho menor organismos de! tipo tifu-coli. Considera SCHILINGMAN, que de los organismos mencionados solo tiene importancia el primero, pudiendo ser considerados los últimos como invasores secundarios, sobre todo los del tipo coli en períodos antemortem o post-mortem, del proceso, ya que los animales de los cuales se les obtuvo estaban ya moribundos, o había muerto de los efectos de la enfermedad, El autor refiere que perros jóvenes susceptibles, que habían sido criados en localidades libres de la infección desarrollaron síntomas clínicos de distemper, 7 a 9 días después de contacto con perros naturalmente infectados, y que no fué difícil hallar en éstos el B. bronchisepticus a partir del primer día en que se manifestaron los síntomas de la afección. Los resultados fueron negativos en cultivos hechos de productos orgánicos de perros jóvenes susceptibles, sacrificados antes de que se expusieran a cualquier infección. Los síntomas del distemper se desarrollaron en perros jóvenes susceptibles, como resultado de la frotis intratraqueales de suspensiones de B. bronchisepticus recientemente aislados. Por todas estas consideraciones concluye que el B. bronchisepticus debería ser considerado como el factor etiológico primario del distemper.
GREEN en 1925 al referirse a su trabajo sobre la encefalitis del zorro, comunicó que había logrado aislar un organismo del género Salmonella el cual sería capaz de reproducir la enfermedad por pasajes sucesivos a diferentes animales. A causa de la similitud de la encefalitis de los zorros con el distemper canino, considera GREEN que el organismo productor de esta enfermedad debe ser uno semejante a aquél, y en efecto, en comunicación aparecida en SCIENCE anunció el hallazgo de un organismo del género Salmonella en los perros, el cual estaría en condiciones de reproducir la enfermedad en estos animales.
ASHE LOCKHART (1926) al referirse a la variabilidad sintomatología de la enfermedad, sobre todo a las llamadas formasde la misma, considera que el distemper canino es una enfermedad sistematizada, en la que los síntomas aparentes de significación exclusivamente secundaría corresponden a expresiones de alteración en órganos o sistemas desvitalizados por la acción de un gérmen de relativa baja virulencia, de acción general. Sí esto fuera verdad explicaría el bien conocido fenómeno de la inmunidad a los varios tipos de distemper en animales restablecidos de una de las formas del proceso. Los organismos bacterianos que se obtienen de los casos de distemper son variados, pero en general están correlacionados con los síntomas y lesiones presentadas. Así en las alteraciones respiratorias están presentes el B. alcaligenes bronchisepticus, el estreptococo, el estafilococo, y en muchos casos de extensas pneumonias el B. alcaligenis bronchisepticus se hallaría prácticamente en cultivo puro. En la forma entérica se halla un organismo del grupo Salmonella, el cual inoculado en fuertes proporciones estaría en condiciones de provocar síntomas similares y lesiones, en perros adultos que se hubieran recuperado de la enfermedad. En las formas nerviosas halla ASHE LOCK - HART en los líquidos cerebrales y espinales, un organismo difícil de ser cultivado. DUNKIN y LAIDLAW (1926) consideran el B. bronchisepticus como secundario, y responsable de las complicaciones pulmonares del distemper, y sostienen la naturaleza filtrable del agente productor de la enfermedad de que habló CARRÉ en 1905.
J. G. HARDENBERGH, al hacer una recopilación de los trabajos concernientes a la causa del distemper canino y en vista de los hallazgos de diversa índole de organismos posiblemente responsables del proceso, en sí, o de sus complicaciones, termina en que cualquiera que sea la tentativa que tenga por objeto reconocer el origen de la enfermedad, se debe contar siempre con los factores de índole clínica y anatómica y que el hecho de que el restablecimiento de un verdadero ataque de distemper, en cualquiera de sus formas, confiera inmunidad para los otros tipos, es significativo para negar la teoría de más de una entidad patológica. Si los esfuerzos para reproducir el cuadro clínico y patológico del distemper canino con virus purificado fallan, el actual rol jugado por los invasores patogénicos secundarios debe ser afirmado, y debe tratarse de lograr, su identidad correspondiente.
Muchos investigadores dudan aun de las conclusiones de DUNKIN Y LAIDLAW a cerca de la etiología por virus del distemper, señalada por CARRÉ. Al ocuparse BILLINGTON y SPA del tema, la atribuyen al B. bronchisepticus, que hallan en una enorme cantidad de casos. El fracaso de SHLINGMAN de provocar la enfermedad inyectando filtrados de emulsiones de bazo, no sorprende, pues permite que el B. bronchisepticus recupere su personería como agente etiológico de importancia. Sus conclusiones de que este agente debería ser considerado como la primera causa del distemper canino, y que los estreptococos y otros organismos, posiblemente del tipo tifo-coli debieran serlo de otros, caracterizan la confusión que sobre el tema aun persiste. BEMELMANS (1930) atribuye la enfermedad a la invasión de los estreptococos comensales que asumen rol patógeno, La escasa seguridad que los datos logrados proporcionan al interés de diferenciar la enfermedad de sus similares hicieron que D. R. WHARTON y M. W. WHARTON iniciaran de su parte la investigación desde otro punto de vista, desde el del conocimiento de las perturbaciones generales que el virus puede ocasionar en la economía general del animal. Ya SABRAZES Y MURATET habían comprobado en una ocasión, en cuatro perros afectados por la enfermedad, ciertas variaciones en los glóbulos rojos, que les había llamado la atención. WHARTON Y WHARTON volviendo sobre el tema llegan a comprobaciones interesantes respecto a modificaciones químicas de los tejidos y humores. De sus trabajos concluyen que el distemper es una enfermedad predominantemente anemizante e hipoglicemiante; que la pérdida de la hemoglobina y del azúcar sanguíneo puede ser demostrada en los animales pocas horas después de la inyección del virus y que el descenso de la hemoglobina es independiente del número de eritrocitos, cuya reducción es evidente. Demuestran además que la sangre de los perros infectados es glicolítica in-vitro, por acción del virus en los eritrocitos y que es independiente de cambios leucitarios o de otra índole debidos a la actividad hemopoiética acrecentada. La infecciosidad de la sangre cosa simultánea mente con la inhibición del virus cuando los límites del descenso del azúcar y de la hemoglobina han pasado las cifras extremas. La sangre de los perros convalecientes desarrolla actividad glicolítica exagerada in vitto, indicando que el actor inhibidor de la glicolisis ha sido parcial o completamente destruído. La proporción de la difusión del azúcar inyectado en la sangre es constante, e indica que los tejidos ambientes no tienen participación en la causa de su descenso en la enfermedad. La albúmina está reducida y la globulina aumentada; la proporción albúmina-globulina cae con el progreso de la enfermedad, y retorna a su cifra normal en la recuperación. El nitrógeno total plasmático no varía grandemente, pero el nitrógeno no proteínico está sometido a una variación apreciable. Esta sin embargo está lejos de mostrarse constante. Disminuye el calcio del suero, el fósforo inorgánico aumenta ligeramente. La velocidad de sedimentación de las células aumenta considerablemente y parece estar asociada con la gravedad específica, la acidosis, el volúmen de las células y otros cambios. La orina no muestra alteraciones que pudieran estar en relación con cambios en el órgano ni con el hígado. La adición del virus a la sangre normal o a la sangre infectada aumenta la proporción de glicolisis. El calentamiento inhibe la glicolisis. El suero hiperinmune, calentado o nó, inhibe la glicolisis. El virus del distemper canino es intracelular y afecta la enzima glicolítica tanto como los factores que controlan la destrucción de la hemoglobina. En los animales libres de complicaciones, las bacterias son rara vez halladas y no pueden ser consideradas como causa de la enfermedad.
B. ROMÁN (1924) al estudiar cerebros de animales muertos de distemper, y al referirse en particular a sus alteraciones anatómicas, encuentra que éstas no tienen nada de específicas, pues si bien se las comprueban en algunos casos, en otros no, en sus características disposiciones. Los síntomas del proceso, el catarro, las conjuntivitis, bronquitis, etc., tampoco son patognomónicas, pues son comunes en el perro, independientemente o asociadas a otras perturbaciones, Para ROMÁN, la enfermedad anatómicamente es una leuco-poliomielitis. El compromiso del asta anterior de la médula, particularmente en su forma de manguitos de infiltración perivasculares, y los mismos del encéfalo, no tienen nada que diferenciarse de las alteraciones similares de las poliomielitis y encefalitis humanas. Las lesiones de la parálisis infantil son, desde el punto de vista anatómico indistinguibles de las encefalitis letárgicas, y no se está equivocado al decir que en algunas oportunidades, hay analogía entre las lesiones del sistema nervioso central en las enfermedades de los animales y las de las parálisis infantil o encefalitis letárgicas, humanas. En el examen anatómico de los casos, ROMÁN halla hiperemia de la laringe, catarro difuso de los bronquios, inflamación de los senos nasales, hiperemia general del cerebro, médula y puente, reblandecimientos circunscritos en estos órganos, hemorragias puntiformes en los mismos. Comprueba alteraciones meningíticas, encefalíticas, degeneraciones acentuadas en las neuronas corticales. No halla grandes lesiones en el hígado, corazón, pulmones, bazo, riñones. El hecho de que en ocasiones se encuentren las lesiones características y en otros nó, posible es que se deba a la existencia de determinados momentos de la enfermedad, sea a causa de que el agente orgánico acabó con el animal antes de que las lesiones hayan tenido tiempo de organizarse específica mente, o a que la duración de la enfermedad haya sido tan prolongada que la lesión original ha desaparecido o a que el animal puede haber muerto de una enfermedad intercurrente. Otro factor que explica la ausencia de lesiones anatómicas es local exclusivamente, pues en el mismo órgano en que no se las halla, hay porciones que lo poseen. Todo esto indica la necesidad de agotar los recursos del examen antes de llegar a conclusiones que no cumplen su papel por lo incompletas. Y no hay que olvidar además la existencia en el hombre del herpes zoster la concomitancia del cual con lesiones medulares ha sido algunas veces señalada por algunos autores, y del mismo modo, como los trastornos motores tanto del distemper como de la poliomielitis infantil guardan relación con lesiones de las astas anteriores, no sería extraño que las vesículas señaladas en el distemper por algunas investigadores, de la piel del abdomen y de la cara interna de los muslos, guarde correspondencia con las alteraciones de las astas posteriores de la médula, que muestran en ocasiones grandes cambios. A. LOCKHART y S. R. JOHNSON (1931 ) al escribir sobre complicaciones meningo-encefalíticas del distemper, son de opinión de que los llamados tipos de la enfermedad, dependientes de la infección secundaria, son en realidad complicaciones de la verdadera enfermedad a virus filtrables Bajo ciertas condiciones esas manifestaciones secundarias son primarias. y pueden producirse sin la asistencia del virus de CARRÉ. De las varias complicaciones de la enfermedad, las más importantes desde el punto de vista del pronóstico son las nerviosas, de las cuales enferman aproximadamente un 50 % de animales y mueren, de éstas un 75 %. No hay que olvidar, sobre todo cuando se hace el estudio comparativo y experimental, que muchas causas externas pueden provocar espasmos como el parasitismo, la acción de ciertas drogas, las alteraciones metabólicas, la presencia de cuerpos extraños en el estómago o intestino. Preparaciones teñidas del líquido de los derrames intraventriculares encefálicos revelaron, casi constantemente, la presencia de organismos bacterianos peculiares. La primera referencia acerca de estos fué consignada en una comunicación de LOCKHART en 1926 a la American Veterinary Medical Association. S. R. JOHNSON, trabajando independientemente de 1923 a 1927 estuvo en condiciones de comprobar muchos de los hallazgos respecto al organismo al cual se suponía productor de la enfermedad primaria de los zorros. Lo halló en la sangre circulante de una gran cantidad de animales estudiados, y lo comprobó en una gran proporción de las muestras recogidas de los espacios ventriculares. Inyectados estos productos, bajo buenas condiciones, fué posible provocar la enfermedad con los síntomas típicos, y lesiones en el sistema nervioso, de las llamadas especificas del distemper espasmódico.
De todo lo anteriormente anotado, hay de evidente, de un lado, las mayores sospechas respecto a la existencia de un virus como productor de la enfermedad, y de otro, las mejor controladas pruebas para explicar su origen bacteriano. El tiempo con sus adelantados recursos nos dirá por fin cual es el agente fundamental del distemper, o si solo se trata, como algunas tratadistas sostienen, de gérmenes de las mas variadas especies, posiblemente específicos, que se insertan en organismos o sistemas profundamente desvitalizados por el desarrollo de un virus que en su acción patológica general compromete la totalidad de las funciones del organismo.
ANATOMÍA PATOLÓGICA
Es muy significativo el hecho de que todos los sistemas de origen ectodémico estén comprometidos : piel, córnea, sistema nervioso; que las lesiones se manifiesten peculiares en ciertas vísceras como los riñones, hígado, y que ciertos de los componentes del sistema cardio-vascular, vasos, miocardio y bazo muestren alteraciones de importancia, en cierto modo específicas.
No hay duda, de otro lado, que gran parte sino todos de los síntomas nerviosos de la enfermedad, guardan relación con las lesiones mas o menos extensas del encéfalo y sus dependencias. Muchos de los signos paralíticos expresan compromiso de las zonas motoras; muchos otros de carácter involuntario como las mioclonias, el movimiento desordenado de las extremidades, en algunas ocasiones tan semejantes a la corea, razón por la que se dió en llamar a la enfermedad corea de los perros, deben guardar correspondencia con perturbaciones del sistema extrapiramidales, como acontece en la encefalitis letárgica humana. La torpeza de los animales enfermos, su inamovilidad deben ponerse, igualmente, en la cuenta de las manifestaciones extrapiramidales; y en las de los cambios cerebelo-vestibulares, los trastornos del equilibrio, la caída unilateral, etc. y no es extraño que el continuo gemido de los animales sea significativo de perturbaciones en el sistema sensitivo.
La primera investigación histológica del sistema nervioso del distemper canino fue llevado a cabo por PIERRET en 1883, Menciona este autor la atrofia de las células motoras, la acumulación de linfocitos y la infiltración en las vainas periarteriales. MATSULEVICH en 1884 en un cuidadoso estudio de la plaga de los perro observó estos mismos cambios, anotando la existencia de nidos celulares, hemorragias, y proliferaciones neuróglicas y endoteliales. Posteriormente trabajos sobre esta materia fueron verificados por MONTANE, 1897; TATY y JACQUIN, 1898; DEXLER en 1903 y 1904; BATTEN y HOLMES, 1907 y 1913; MARCHAND Y PETIT, 1912; CERLETTI, 1912; FLEXNER y CLARK, 1913; M. GOWAN y DAWSON, 1914; KANTOROVICZ y LEWI, 1923; GOLDBERG y VOLGENAU, 1925; ROMÁN y LAP, 1924: PUGH, 1926, etc.
Es difícil precisar el sitio de las lesiones primitivas. Se creyó en un principio que las manifestaciones catarrales y nerviosas podrían contar con cambios anatómicos correspondientes, pero CERLETTI (1912), comprobó la existencia de lesiones del tejido nervioso en ambas, naturalmente mas intensas en los casos agudos y en las formas predominantemente nerviosas de la enfermedad.
MARINESCO y, colaboradores que han sido los que mas documentadamente han contribuido a la anatomía patológica de la enfermedad señalan cambios notables, de carácter agudo, de las neuronas y de las células glióticas, con la aparición en ellas de las inclusiones nucleares y citoplasmáticas específicas del proceso. Las lesiones no solo toman el componente gris del encéfalo, sino el blanco, acusándose en éste, de notable manera, una particular degeneración de la mielina que da lugar a la formación de imágenes areolares o enrejadas. Para estos autores las lesiones de la corteza varían según haya o no alteración infiltrativa en la misma. Cuando no la hay, las lesiones se reducen a cromatofilia de los cuerpos celulares, engrosamiento de sus prolongaciones, y de la membrana nuclear, discreta hiperplasia glial. Las alteraciones son grandes en cambio, cuando hay compromiso inflamatorio, Entonces, se halla pulverización de los cuerpos de NISSL, degeneración alveolar del protoplasma, transformación del cuerpo celular en verdadera sombra, con intervención de procesos de fagocitosis mas o menos desarrollados, cambios en las células de HORTEGA, y en la microglía de la substancia blanca, sobre todo a nivel de las infiltraciones vasculares. MARINESCO y colaboradores juzgan que el agente productor de las lesiones de las células nerviosas es el mismo que el que ha actuado sobre los elementos de la glía, y que el proceso degenerativo del tejido debe ser reciente por la inexistencia de cuerpos granulosos.
Se trata para ellos, de una leuco-polio-neuroaxitis. Las lesiones son de naturaleza degenerativa e inflamatoria. Los procesos primitivos estarían representados por los cambios de naturaleza inflamatoria perivasculares : manguitos pericapilares linfocito-plasmáticos. Todos los vasos están comprometidos, desde los grandes en que predominan los linfocitos hasta los capilares en los que hay abundantes células plasmáticas. El proceso de pericapilaritis es bastante neto y en su extensión dá al cuadro el aspecto de una verdadera infiltración intersticial. La inflamación está localizada a nivel de las meninqes, en la substancia blanca y en la gris. En la médula el proceso parece propagarse desde la periferie hasta las formaciones periependimarias, provocando en su recorrido una infiltración difusa. A nivel del bulbo y de la protuberancia las alteraciones son sobre todo marginales. Una cierta predilección parece existir por los territorios anexos al locus-niger. La localización casi constante cerebral del proceso inflamatorio en la enfermedad, a nivel de la base, hipocampo, bandeleta óptica, etc., circunstancias a menudo comprobadas en el presente caso, no halla otra explicación que el papel que en la propagación de la enfermedad debe desempeñar el líquido céfalo-raquídeo, sin que se trate de negar por esto, el rol del medio sanguíneo comprobado por las inoculaciones positivas a los animales sensibles, y por el hallazgo de lesiones progresivas de los vasos, e intersticiales de los órganos. Esta doble infección puede ser en cierto modo la causa de la extensión de las lesiones tanto en la substancia blanca como en la gris, con una cierta electividad de la substancia blanca del cerebelo y del cerebro. Por el carácter perivascular de la degeneración inielínica, se aproximaría esta enfermedad a las encéfalomielitis post-eruptivas y post-vaccinales; por su compromiso de la substancia gris, a la poliomielitis, la rabia, y las leuco-neuro-axitis.
De MONBREUN refiere no haber hallado grandes lesiones en la médula espinal y en los ganglios espinales, en los perros que padecían de la forma catarral simple sin manifestaciones nerviosas, ni en los que padecían de la típica forma nerviosa caracterizada por convulsiones generalizadas.
Las meningitis son corrientes. Las membranas se encuentran infiltradas con polinucleares y mononucleares grandes y medianos. La proporción he mononucleares es mayor en las formas crónicas, y en las formas severas; mayor también la infiltración de las meninges, que la de los vasos de la corteza. En los casos corrientes, las proporciones entre los polinucleares o mononucleares es igual.
Pero aparte de las lesiones descritas a cerca de los cuales insistiremos en su oportunidad es halla el hecho muy frecuentemente comprobado, de la existencia de inclusiones citoplasmáticas e intranucleares, al parecer específicas, y semejantes a los corpúsculos de Negri de la rabia, visibles en las células nerviosas, en los endotelios vasculares y en los de algunos otros órganos. LENTZ al ocuparse de la enfermedad, en sus formas nerviosas ( 1907) se refirió a ellas, describiéndolas como corpúsculos desprovistos de estructura, de un diámetro menor que el de un glóbulo rojo, presentes en las zonas destruidas del asta de Ammon. Volviendo sobre el tema en 1909, comprueba sus primitivos hallazgos, y emite la opinión de que como en el caso de los corpúsculos de Negri, se trata solo de masas pequeñas de cromatina, sin la significación parasitaria, que como a los corpúsculos de Negri, se les pretendía atribuir. Algún tiempo después STANDFUSS halla la misma degeneración celular e igoal presencia de corpúsculos en la forma nerviosa de la enfermedad. SINIGAGLIA en comunicación presentada por GOLGI al Congreso de Patologistas de Turín, confirma la presencia de las inclusiones celulares, que difieren sin, embargo de las descritas por los autores alemanes, por poseer una estructura íntima, y presentarse en otros sitios, bajo la forma de cuerpecitos coloreables por el Romanowsky en azul, ser de volumen variable y poseer en conjunto un aspecto murifrome. SINIGAGLIA refiere haberlos comprobado en diferentes partes del sistema nervioso, con excepción del cuerno de Ammon; en las células que tapizan los ventrículos internos del cerebro, en ciertas de las epiteliales de la córnea, en el epitelio bronquial de perros enfermos con distemper que hubieran desarrollado síntomas broncopulmonares o catarrales. BAVES y STARCOVICI reemprendiendo estas investigaciones en perros afectados de esta enfermedad comprueban en unos casos, con lesiones aparentes celulares del asta de Ammon, la presencia de las inclusiones sin estructura descritas por LENTZ, y en otras, con lesiones celulares discretas de la corteza, bulbo, cerebelo y asta de Ammon, la de grandes corpúsculos, mas grandes que los de la rabia, y bastante diferentes de los descritos por LENTZ y SINIGAGLIA.
Estos corpúsculos se caracterizaban, sobre todo, por su estructura interior, por las lesiones citoplasmáticas particulares, concomitantes de las células que los albergaban, y por ser mas refringentes que los corpúsculos de Negri. En este caso, ni el resto del cerebro, ni las células de la cubierta ependimaria, ni las de los bronquios, tenían inclusiones semejantes, razón que justificaría, según BAVES Y STARCOVICI la aceptación de una tercera variedad de corpúsculos en el distemper canino. Refiriéndose a los cuerpos descritos por SINIGAGLIA en el epitelio bronquial, los hallaron en formas coreicas y bronco-pulmonares del proceso, en el espacio que queda entre el núcleo y el borde libre de las células epiteliales bronquiales y en el lugar en que normalmente se hallan los canaliculos de Holmgreen, en una posición semejante a la que adoptar algunos coccidios en las células cilíndricas del intestino. Los hallan también en los frotes de la mucosa de los bronquios, en la posición descrita, y en epitelio descamado de las células alveolares del pulmón. SANFELICI en 1915 halla inclusiones similares citoplasmáticas en las células nerviosas, conjuntivales, intestinales, pancreáticas, de la piel, de los nódulos linfáticos, médula ósea, ovario, pulmón; nó en los riñones, hígado y glándulas salívares. Los describe como constituídos por cuerpos homogéneos, sin estructura o vacuolares, y atribuye su existencia a las reacciones de las células contra los virus. No admite su naturaleza de protozoario, y piensa que puede tratarse de productos protoplasmáticos o nucleares.
KANTOROWIVZ y LEWY en 1923 describen inclusiones intracelulares en el cerebro de perros que habían padecido de la forma nerviosa del distemper. Según la opinión de estos autores se trataría del parásito Encephalitozoon cuniculi, que comunmente se presenta en el cerebro de los conejos. ROMÁN y LAPP en 1925, al estudiar el sistema nervioso de perros que habían sufrido de distemper, hallan, de 27 animales examinados, 10 con alteraciones del sistema nervioso central bajo la forma de procesos diseminados, no supurativos de la médula espinal, cerebro y leptomeninges, sin inclusiones asociadas. PUGH en 1926 en un estudio de 50 casos de encefalitis epidémica canina comprueba lesiones en los ganglios basales de los casos en los que la afección estaba acompañada de trastornos letárgicos, anotando la presencia de reacciones perivasculares extensas asociadas con hemorragias minúsculas. A causa de la extraordinaria similitud de esta enfermedad con ciertos procesos en el hombre, consideró la posible relación entre ambos. DUNKIN y LAIDLAW en sus investigaciones de la enfermedad en los hurones, concluyeron en que el examen histológico no proporcionaba suficientes testimonios de alteración característica; que si bien es cierto que en algunos animales habían comprobado la existencia de alteraciones en las células nerviosas, no hallaron las reacciones inflamatorias que comunmente se les asocian, y que, en algunos casos de la enfermedad provocada experimentalmente no se encontraban las inclusiones celulares con la regularidad y número de las mencionadas por otros investigadores. NICOLAU en 1935 describe cuatro casos de distemper fatal, en los que comprueba la existencia de inclusiones protoplasmáticas similares a las de LENTZ, ampliamente distribuídas en todas las células derivadas de los tres tipos embrionales. Las inclusiones nucleares en cambio solo se comprobaron en las células nerviosas y neuróglicas. Según NICOLAU se trata de inclusiones,etimológicamente hablado, y no de parásitos libres, menos aun, del grupo del Encephalitozoon. No hay analogía como lo piensa MANOUELIAN, ni menos semejanza entre las inclusiones propias de la enfermedad, excesivamente variables, en forma y dimensiones, y el Encephlitozoon cuniculi. La sola propiedad común es la afinidad tintorial. Es igualmente la impresión de MARINESCO y sus colaboradores. Por estas razones NICOLAU considera justo rechazar las calificaciones de Negria canis (SINIGAGLIA) y de Encephalitozoon Negri (MANOUELIAN), que implica la idea de parásitos definidos. NICOLAU propone llamarlos, simplemente, corpúsculos de LENTZ. del nombre del primero que los observó: ellos caracterizan la enfermedad de CARRÉ lo mismo que los corpúsculos de Joest-Degen la enfermedad de Borna, y los corpúsculos de Negri, la rabia. BENJAMÍN excluye el origen parasitario de las inclusiones, los cree productos de destrucción de los nucleolos, o nucleolos que han emigrado al protoplasnia. BENDORF refiere haberlos hallado en seis casos nerviosos de distemper, graves, no en las formas ligeras ni en los perros normales. Para MARINESCO y sus colaboradores las inclusiones descritas en la enfermedad de los perros entran en el mismo grupo de otras, comprobadas en enfermedades a ultra-virus. Según estos autores, SINIGAGLIA Y MANOUELIAN Y VIALA carecen de pruebas para afirmar que se trata de fases panesporoplásticas del virus, tanto más cuanto que los corpúsculos intracelulares de estas infecciones no ofrecen las reacciones de la cromatina nuclear como ocurre con los núcleos de los protozoarios Encephalitozoon cuniculi, Klosiella muris, y el hematozoario de Laveran. Tampoco creen MARINESCO y sus colaboradores que sean productos no específicos de degeneraciones de nucleoplasma como parece que admiten ABNER, WOLF. y SAMUEL ORTON, y termina expresando que probablemente sean debidas a la acción del virus sobre las células, pues no las hallan en los perros normales.. Debido a su presencia no sólo en las células nerviosas sino en las neuróglicas, estas formaciones tienen la Particularidad de que no se encuentran en otras infecciones neurotropas. Por su localización sobre todo intranuclear que denota que el virus es particularmente karyotropo, y en menos escala cytotropo, se aproximan a las halladas en la en cefalomielitis equina (Enfermedad de Borna). H. R. KREISEL en sus investigaciones sobre la encefalitis experimenta¡ de los zorros, y sus comprobaciones histológicas comparativas con el distemper canino, encuentra, en los tejidos de los perros jóvenes inoculados experimentalmente con el virus de la encefalitis de los zorros, inclusiones nucleares. En se ries de 54 perros jóvenes experimentalmente infectados con el virus del distemper canino halla inclusiones nucleares similares en el 10 % de los casos, inclusiones citoplasmáticas en el 63 %. Las inclusiones nucleares fueron observadas en las células hepáticas, en las de Kupfer, en el retículo-endotelio de las glándulas linfáticas mesentéricas y del bazo, y en el epitelio glomerular del riñón. Inclusiones citoplasmáticas fueron halladas en el epitelio de los bronquios, de los conductos biliares, en el retículo endotelio del hígado, de las glándulas linfáticas del mesenterio y bazo. KRIESEL refiere que las inclusiones nucleares de la encefalitis de los zorros y del distemper, canino están constituidas de gránulos mi núsculos que se tiñen en rosado y que su agregación dá el aspecto homo géneo con que se les vé comúnmente. Inclusiones citopiasmáticas fueron observadas en 7 hurones infectados con el distemper canino. ASHE LOCKHART y S. R. JOHNSON en su comunicación sobre complicaciones me ningocerebrales del distemper canino refieren haber hallado imágenes de degeneraciones celulares en los elementos motores de la corteza principalmente, con la concurrencia de cuerpos intracelulares que pueden confundirse con los corpúsculos de Negri. Estos cuerpos pseudo-Negri se diferenciarían de los verdaderos, por su independencia de los gránulos nucleares y la vaguedad de sus contornos. De MONBREUN en 1937 publica uno de los más completos trabajos sobre el distemper. Estudia detalladamente la enfermedad en sus expresiones clínicas y anatómicas tanto en la forma natural, como en la provocada experimentalmente por inoculaciones de emulsiones de virus filtrados, por vía intravenosa, intraperitoneal, intracerebral, nerviosa, subcutánea, intradérmica, intranasal, intratraqueal, etc., etc. Encuentra en la mayor parte de los órganos. en sus componentes endoteliales, como en las cubiertas de los vasos, y también en los elementos de los parénquimas, alteraciones de tipo degenerativo, pero fundamentalmente las inclusiones específicas intracitológicas e intranucleares a las que se refieren la mayor parte de los tratadistas de la enfermedad.
Para sus comprobaciones en la enfermedad natural, emplea De MONBREUN tres grupos de perros: el grupo I consistente en animales que han desarrollado la forma catarral de la enfermedad sin presentar síntomas que sean referibles al sistema nervioso central; el grupo II representado por perros que han sufrido la forma típica nerviosa, caracterizada por las convulsiones severas generalizadas; el grupo III por animales que desarrollan un proceso extraordinariamente rápido, de horas, y que mueren en coma, sin evidencias de parálisis o convulsiones, Refiere así sus hallazgos en el examen del sistema nervioso central. Las secciones de la médula espinal de los grupos I y II, y las de los ganglios espinales del grupo II no revelaron nada significativo. En un caso las meninges que cubrían el cerebelo cerca de los troncos cerebrales estaban infiltrados con mononucleares, algunas de las cuales contenían inclusiones nucleares similares a aquellas de las células endoteliales del bazo y de los ganglios linfáticos. En la capa cerebelar subyacente, mas allá de la línea de las células de Purkinje, se comprobó la existencia de una manifiesta proliferación de las células micro y macrogliales, en las cuales, se halló inclusiones nucleares y protoplasmáticas; corrientemente ambos tipos de inclusión en la misma célula. Las inclusiones nucleares aparecen como cuerpos simples, homogéneos, de contornos irregulares y fácilmente demostrables en las tinciones por la hematoxilina-eosina. Las inclusiones citoplasmáticas, algunas veces en número de tres en la misma célula aparecen como cuerpos pequeños, vacuolados, acidófilos. Se muestran mejor con el carbol-anilina-fuchsina de Goodpasture y con las tinciones con el método de Wright. En este grupo no fueron comprobadas inclusiones en las células nerviosas. En dos casos del grupo II, de animales que habían muerto como consecuencia del desarrollo de la forma nerviosa típica de la enfermedad, fueron halladas inclusiones de ambos tipos, nucleares y citoplasmáticas, tanto en las células nerviosas como en la macro y microglía. El examen microscópico de porciones del cerebro y de la médula espinal de perros del grupo III, presentó numerosas y minúsculas hemorragias, con edema y proliferación de las células del endotelio capilar. Algunas de estas células, hinchadas, contenían las inclusiones nucleares idénticas a las de cualquier otro tipo de células endoteliales. Comprueba las referidas inclusiones, también en el retículo endotelial del bazo, hígado, ganglios linfáticos del mesenterio; en los grandes mononucleares que forman las infiltraciones perivasculares de los tejidos, en las células de descamación de los endotelios alveolares del pulmón, en el epitelio descarnado de los pequeños bronquios. La especificidad de esta lesión está indicada –dice- por la presencia de inclusiones citoplasmáticas y nucleares en el epitelio bronquial y alveolar. Estas inclusiones han sido observadas en los casos experimentales sólo en aquellos en los que, la inoculación se ha efectuado por vía respiratoria, lo que unido al hecho de la frecuencia con que la enfermedad aparece naturalmente en animales que no tienen otro medio de contacto que el del ambiente hace suponer que la común puerta de entrada de la infección sea el tramo respiratorio. No es extraño que lo sea también como consecuencia de las mordeduras ya que De MONBREUN logró producir la enfermedad por inyección subcutánea o intradérmica y que otros investigadores hubieran mencionado la existencia del virus en la saliva de los perros enfermos. La patología microscópica de los casos descritos por De MONBREUN son casi similares a la llamada encefalitis epizoótica de los zorros de GREEN, tal como la describieron GREEN, ZEIGLER, GREEN y DEWEY. Estos investigadores no mencionan las inclusiones citoplasmáticas y nucleares de las células bronquiales y de los alveolos pulmonares, que corrientemente halla De MONBREUN en los perros afectados por distemper natural adquirido. Últimamente, en un posterior estudio sobre la encefalitis de los zorros, GREEN y sus colaboradores, encuentran inclusiones nucleares, pero nó inclusiones citoplasmáticas en las células del epitelio bronquial de los tramos superiores del árbol respiratorio, de zorros que han muerto de enfermedad natural. En los casos de enfermedad provocada en estos animales, las inclusiones aparecían sólo cuando la enfermedad experimental tenía un curso prolongado. GREEN y SHILLINGER, logran trasmitir la enfermedad de los zorros al perro, y expresan la opinión que la enfermedad probablemente ocurre naturalmente en estos animales. GREEN y DEWEY no logran infectar hurones con la encefalitis del zorro, pero lo mismo que DUNKIN y LAIDLAW comprueban que estos animales son extremadamente susceptibles al distemper canino. Con excepción de los abscesos de los labios, lengua y paredes abdominales, las lesiones histológicas presentadas por estos animales son las mismas que las comprobadas en el dístemper experimental de los perros. Entre ellas se logra comprobar las características lesiones de! cerebro y la presencia en las neuronas y elementos glióticos de las inclusiones intracitoplasmáticas e intranucleares. Para DE MONBREUN el virus del distemper es evidentemente citotrópico. juzgando por la presencia de las inclusiones en la mayoría de los componentes de los tejidos, las células derivadas de todos los componentes embrionales son susceptibles de afectarse. El hecho de que los endotelios se comprometan especialmente es signo de que la infección toma la vía sanguínea para propagarse en el huésped.
Tanto en los animales que enferman espontáneamente, perros, hurones, como en aquellos en los que se ha provocado la enfermedad experimentalmente, se comprueba la existencia de proliferaciones endoteliales vasculares, infiltraciones a mononucleares, pericapilares. En ambos tipos de lesiones se halla inclusiones nucleares tanto en los mononucleares pericapilares como en las células del endotelio de los vasos. Iguales inclusiones se halla en las células nerviosas, astrocitos y microglía. Hay mucha semejanza entre las inclusiones citoplasmáticas del distemper y las de la rabia, pero la existencia concomitante de inclusiones nucleares en la primera sirve para establecer el diagnóstico. Este dato vale mas que las particularidades tintoriales y de contorno que no son seguros siempre. DE MONBREUN cree que las células nerviosas son menos afectadas que las de los endotelios, debido al hecho de su elevada diferenciación funcional y a que hay que esperar que se desarrollen las formas nerviosas de la enfermedad, sólo en aquellos animales cuyas respuestas inmunológicas al virus son particularmente lentas. Además de los tipos clínicos corrientemente aceptados en la enfermedad tales como el catarral, nervioso o encefalítico, intestinal y tegumentario, DE MONBREUN describe otro que propone llamar encefalítico agudo, o mas propiamente, tipo agudo encéfalo-mielítico cuya característica es tener curso fulminante y comprometer en forma extensa el endotelio vascular, principalmente del cerebro, y de la médula, y el retículo endotelio y células del hígado. Muchos investigadores creen que las diferentes formas de la enfermedad se deben a la existencia de diversos virus, pero las observaciones de otros, y en especial las de DE MONBREUN, comprueban, que las manifestaciones catarrales se hallan en casi todas las formas, y que si en algunas oportunidades no se presentan, se debe a la circunstancia del predominio de ,otras; que con la misma cepa de virus ha podido producirse en unos animales la forma catarral exclusiva, en otras la forma nerviosa, en otras por último la forma encefalítica aguda; que en el tractus gastro-intestinal, especialmente en el colon de algunos animales en experiencia, se comprobó la existencia de actividad del virus, un hecho que podría explicar las diarreas en la enfermedad, Raras veces se comprobó en la forma del proceso natural, la existencia de abscesos de la pared abdominal, y en la experimental, en ninguna ocasión. El hecho de desarrollarse abscesos iniciales en los labios, lengua y paredes abdominales, y la presencia en <ellos de las inclusiones nucleares y citoplasmáticas lo mismo que en los tejidos adyacentes a los mismos proporcionan el testimonio de aquellos han sido provocados por el virus. El virus del distemper es uno de los pocos en determinar el desarrollo de inclusiones nucleares y citoplasmáticas. Otros los hacen como los de la viruela, paravaccinia, y alastrina. Para muchos investigadores se trata aquí sólo de formas modificadas del mismo virus. A. LOCKHART y S. A. JOHNSON, (1931) señalar, como muy discretas las alteraciones del tejido nervioso en el cerebro, entre las que habría que señalar la presencia de una gran cantidad de líquido en las cavidades ventriculares, de apariencia opaca y conteniendo proporción considerable de flóculos. El corazón presenta hemorragias endocardiacas y endopericardiacas. El bazo es grande y presenta también hemorragias subcapsulares. En el cerebro dominan las infiltraciones perivasculares a base de polinucleares y mononucleares, más frecuentes en la corteza si las infiltraciones meningíticas son mas acentuadas y predominantemente a mononucleares si las infecciones son de algún tiempo. Las recientes en cambio muestran mas abundantemente las polinucleares, Los autores refieren haber comprobado lesiones degenerativas en las células nerviosas de la corteza gris del cerebro, y la presencia de cuerpos, intracelulares semejantes a los cuerpos de Negri, de los que se diferenciarían por la escasa nitidez de sus contornos y por su independencia de sus gránulos del núcleo. Aparte de las alteraciones nerviosas de la enfermedad, cuya bibliografía hemos examinado con algún detalle, casi la mayor parte de los restantes tejidos del organismo han sido señalados como portadores de alteraciones generales, y en veces, particulares, con la presencia de las inclusiones celulares a las que nos hemos referido en anteriores líneas. Así se ha comprobado en los pulmones lesiones de pneumonia intersticial, y en la mucosa de los bronquios medianos y pequeños, infiltración mono y polinuclear y desprendimientos epiteliales. Tanto las células que tapizan los alveolos como las de los bronquios presentan inclusiones citoplasmáticas y nucleares, de forma oval o redonda, únicas o varias, mas frecuentes las primeras que las segundas, ocupando las citoplasmáticas la porción intermedia entre el núcleo y la pared citoplasmática, y con las mismas características -según DE MONBREUN que las descritas por GOLGI y SINIGAGLIA. A causa de la constancia con la que incluiones de esta especie aparecen en las cubiertas epiteliales de los bronquios de perros muertos de distemper, se ha considerado a esta alteración como característica de la enfermedad. Con el fin de comprobar esta sospecha se examinaron varios animales sanos y otros sospechosos de haber muerto de rabia. En ninguno de ellos se llegó a comprobar inclusiones de esta naturaleza.
Las lesiones hepáticas varían según la calidad y magnitud de la infección. En los perros que padecen de manifestaciones catarrales simplemente, y en los que han presentado síntomas evidentes de alteración nerviosa traducida por manifestaciones contracturales generalizadas, las células hepáticas muestran evidencias de un hinchamiento enturbiado de sus protoplasmas, y de necrosis parcelares con acentuación de sus expresiones infiltrativas a base de mononucleares. Ni las células hepáticas, ni las células de KUUFER presentan inclusiones; solo el epitelio de los canalículos biliares muestra inclusiones y de la misma especie de las de los bronquios. En perros que han muerto de infección violenta, de 30 horas, con marcados trastornos nerviosos, las lesiones hepáticas eran distintas: las inclusiones se manifestaban evidentes en las células hepáticas y en las de KUPFER.
El bazo presenta en la mayor parte de los casos una considerable hipertrofia folicular malpighiana a base de masas de mononucleares proliferados y teñidos pálidamente. El tejido linfoide está notablemente reducido, confinado a la periferie de los folículos. Muchos de los mononucleares centro foliculares presentan signos de necrosis, contienen restos de hematíes o leucocítos fagocitados, o albergan inclusiones citoplasmáticas o nucleares de la misma especie que las de las células de KUPFER o las del hígado de perros muertos con infección aguda. No es extraño encontrarlos en las células endoteliales de los senos del órgano.
En los ganglios linfáticos del mesenterio las células del retículo-endotelio muestran evidencias de gran actividad, lo mismo que una notable disminución de los linfocitos. Son frecuentes las inclusiones nucleares de las células del retículo-endotelio, de la misma especie de las mencionadas en el bazo, lo mismo que cuerpos pequeños, vacuolados, rojos, en el citoplasma, que es difícil diferenciarlos satisfactoriamente de los glóbulos rojos.
En lo s intestinos y el estómago es frecuente hallar hemorragias minúsculas en la mucosa, e infiltraciones de mononucleares de la sub-mucosa, muchas de las que contienen inclusiones nucleares. Semejantes inclusiones se hallan también en las células epiteliales glandulares M colon y en las células basales de la mucosa gástrica.
DISTEMPER CANINO Y ENCEFALITIS EPIZOOTICA DE LOS ZORROS
En el curso de esta exposición nos hemos referido continuamente a la encefalitis de los zorros, enfermedad con la que el distemper tiene grandes semejanzas, tanto clínicas como anatómicas. Para ilustrar este trabajo, como para sugerir, entre nosotros, investigaciones sobre la materia, exponemos en el siguiente capitulo el estado de la cuestión actualmente.
El distemper canino y la encefalitis de los zorros han sido consideradas a menudo como la misma enfermedad. LAIDLAW y DUNKIN lo anotaron así, y refirieron que LAOSSON no encontró en 1882 dificultad en trasmitir el distemper a dos zorros cautivos. GALLI y VALERIO hicieron mención del desarrollo de un caso de distemper canino en un zorro que había sido inoculado con una bacteria procedente de un caso de distemper canino. ALLEN en el Canadá estudiando las diferentes formas clínicas de la enfermedad sospechó que podía ser provocada por un virus ROMÁN y LAPP al comprobar que la patología cerebral de la encefalitis de los zorros era similar al distemper, sugirieron la posibilidad de que ambas enfermedades podían ser provocadas por el mismo gérmen. R. G. GREEN, N. R. ZEIGLER, B. B. GREEN y E. T. DEWEY establecen que la enfermedad es común a los zorros blancos y rojos en cautiverio, que es provocada por un virus que invade la sangre y que puede ser hallada en diferentes tejidos del organismo. Las alteraciones patológicas son pronunciadas en el cerebro y médula espinal. Por sus síntomas le conviene la designación de encefalitis, pero por la naturaleza de los cambios anatómicos convendría llamarla mejor mielo-encefalitis. No son de opinión de que se trate de la misma enfermedad, pues en las investigaciones experimentales la enfermedad del zorro si bien era fácilmente trasmisible a los perros no lo era a otros animales de laboratorio como las ratas, cobayos, conejos. Los hurones tan susceptibles al distemper canino no lo son a la encefalitis de los zorros; en cambio el distemper canino es fácilmente trasmisible a los zorros. Las investigaciones de GREEN y sus colaboradores que abarca una población de cerca de 30,000 animales, con 1,000 zorros que sirvieron para el estudio experimental y aproximadamente 2,500 autopsias les dieron oportunidad para concluir en que las dos enfermedades eran distintas; que los perros que se habían recuperado de una infección con el virus de LAIDLAW y DUNKIN no manifestaron inmunidad al virus de GREEN; que el serum hiperinmune que se había mostrado tan eficaz para la supresión del virus de la encefalitis del zorro no producía similares resultados a los del virus del distemper canino, probado en las inoculaciones de zorros y hurones. Considerables dosis de virus de la encefalitis del zorro no ocasiona síntomas de la enfermedad en los hurones Jóvenes y tales inyecciones fallan para estimular la inmunidad contra la subsecuente inoculación con el virus de LAIDLAW-DUNKIN. Los síntomas de la enfermedad fueron observados en los animales inoculados experimentalmente lo mismo que en los que desarrollaron la enfermedad naturalmente. Se traducían por pérdida del apetito, convulsiones de diversa índole, hiperexcitabilidad, letargia, parálisis flácida y espástica, temblores musculares, estados semejantes a las alucinaciones, intranquilidad, miedo, gritos semejantes al aura epiléptico y coma. Los animales acusaban marcada desnutrición, descargas oculares y nasales bajo la forma de secreciones simples o purulentas, cámaras mucosas a veces teñidas de sangre, o francamente hemorrágicas.
Lo mas notable en el examen anatómico fueron las escasas lesiones anatómicas de los órganos, pues a no ser por las hemorragias de diversa. intensidad en los órganos, unas veces tan escasas que pasan desapercibidas, o no existen, y otras tan marcadas que constituyen el cuadro íntegro de la alteración, no se comprobaron otras. Las hemorragias eran tan marcadas en oportunidades que las cápsulas suprarrenales aparecían como simples coágulos. Hemorragias intensas llenaban las cavidades bronquiales, lo mismo que las cavidades abdominal y toráxico, o la cámara interna del ojo bajo la forma de gotas en el humor acuoso, o mezcladas en forma tal que daban a la pupila un brillo rojo. El corazón estaba aveces particularmente afectado por hemorragias intramurales, intrapericárdicas u sub-endocárdicas, y considerables hemorragias podían verse en el pancreas, timo, glándulas sub-maxilares y pulmones. El tejido nervioso era asiento de estas mismas hemorragias, desde las puntiformes, en diversas porcione s de la substancia gris y blanca, hasta las grandes extravaciones que comprometían hemisferios íntegros o zonas mas o menos extensas. Todas ellas no tenían otra significación que la acentuación de las extravasaciones pericapilares, elemento del cambio anatómico en esta alteración. Era natural suponer la calidad y extensión de la parálisis en el caso en que hemorragias de tal magnitud se produjeran. Cuando la epizootia era de curso largo, no era raro comprobar pneumonia, icteria, marcadas infiltraciones gastro-intestinales, todo lo cual significaría o una complicación simple del proceso, o la actuación de un agente bacteriano que acompaña al virus en su pasaje de animal a animal.
Como se esperaba los cambios histológicos en el sistema nervioso eran marcados. Al lado de los derrames sanguíneos ya descritos, frecuente era encontrar infiltraciones de células redondas, perivasculares, que significaban más la reacción del tejido nervioso a la enfermedad, que alteración típica de ella. Todas presentaban el característico aspecto de los manguitos perivasculares, tan frecuentes en diversas lesiones del tejido nervioso, tanto en las infecciones como en las tóxicas. Nidos celulares aparentemente asociados con capilares, constituídos por elementos del tejido fundamental hiperplasiado, eran frecuentes, lo mismo que las infiltraciones de las meninges en el fondo de las cisuras o fuera de ellas. Era frecuente también el hallazgo de las proliferaciones endoteliales vasculares, tan intensas en oportunidades que llegaban a oclusiones verdaderas. Raros eran los trombos. El canal central estaba dilatado y proliferado su epitelio.
Para estos autores la causa de la enfermedad es un virus filtrable que en solución salina tenía la propiedad de pasar el filtro Berkefeld. Cultivos bacterianos de gérmenes anaerobios y aerobios de zorros muertos con la enfermedad se mostraron negativos, excepto para los estreptococos, los que con sorprendente frecuencia podían ser aislados en cultivos puros, de los cerebros de los animales. Tales cultivos no provocaban. una infección semejante a la determinada por el virus proveniente del, animal del cual derivaban los cultivos. El virus tiene una distribución amplia, pues se le halla en la sangre, médula, bazo, cerebro.
Posteriormente en 1933, R. G. GREEN, N. R. ZIEGLRE, W. E. CARLSON y colaboradores, en un estudio referente a las propiedades generales y patogénicas del virus de la encefalitis epizootica de los zorros, concluyen en que el virus de esta enfermedad permanece viable en el cadáver del animal muchos días después de su muerte, y que puede ser conservado en una emulsión glicerinada al 50 % durante muchos años, sin, pérdida de su virulencia. E! virus es fácilmente filtrable a través de un filtro Berkefeld y las inclusiones específicas típicas de la enfermedad ocurren en los animales infectades, con los filtrados. En las infecciones experimentales producidas en coyotes mediante inyecciones cisternales. las inclusiones intranucleares se presentan con bastante frecuencia, en las células endoteliales de las meninges, con menos frecuencia en la substancia. cerebral, y rara vez en las células ependimarias. Se las halla también en las células hepáticas y en las endoteliales de los cordones hepáticos, de los senos del bazo, del riñón, de las, cápsulas suprarrenales; en los elementos del retículo-endotelio del bazo, del hígado, ganglios linfáticos y en las células corticales de las cápsulas suprarrenales. El zorro gris es resistente a la infección lo mismo que el visón. Las tentativas de producir infección con este virus en los conejos, incluso empleando procedimientos especiales, fracasaron enteramente. El virus parece no patogénico para las ratas blancas, cobayos, ardillas y gatos. Los perros acusan ser extraordinariamente susceptibles al virus. Las inclusiones intranucleares típicas del virus se producen abundamente en el perro, no solo en el sistema nervioso central sino en los órganos toráxicos y abdominales. Los hurones aparecen resistentes a la infección con el virus. Los carneros enteramente inmunes, lo mismo que los monos.
GREEN y SHILLINGFR (1933), en la descripción de la infección experimental en los perros, por el virus de la encefalitis de los zorros, terminan en que los perros pueden ser experimentalmente infectados con el virus de la encefalitis de los zorros. En algunas oportunidades hay alguna irregularidad en la trasmisión de la enfermedad que explican sobre las bases de una inmunidad familiar natural. La infección fatal es de curso breve, de una semana aproximadamente, lo mismo que en la enfermedad natural de estos animales. La secreción lagrimal y nasal de los animales enfermos tienden a ser purulentos. Es frecuente que el compromiso del sistema nervioso se traduzca por síntomas generales que se manifiestan por excitación y verdaderos ataques de carreras. La patología general de la enfermedad consiste en la infiltración celular en el sistema nervioso central y en la necrosis focales del hígado. Las inclusiones intranucleares específicas se hallan en las células del endotelio vascular, de las meninges, del retículo-endotelio, y ocasionalmente en las células corticales de las cápsulas suprarrenales.
Posteriormente R. S. GREEN, B. B. GREEN y colaboradores en sus investigaciones acerca de la encefalitis epizootica de los zorros, demostraron que bajo condiciones naturales y experimentales, era grande la afinidad del virus por el tramo respiratorio superior. La inoculación muscular de las secreciones de esta zona provocaban la enfermedad en los animales en experiencia, afinidad anatómica por el virus, demostrada además de un lado por la presencia de las inclusiones intracelulares específicas de las encefalitis en las células epiteliales de esta zona, y porque una inoculación del virus intramuscular en un animal provoca en éste la aparición del virus en la porción superior del tramo respiratorio. Todo parece indicar que este tramo es la puerta de entrada del virus de la encefalitis de los zorros como, según opinión de algunos autores, acontece con el distemper, el cual gana su entrada a favor de las lesiones que causa en las células epiteliales que cubren la región, lesiones que de otra parte no significan hecho particular anatómicos, sino expresión parcial de la infección general. La persistencia del virus en los tejidos del árbol respiratorio superior sería la explicación de los portadores del virus, en el aspecto epidemiológico de la enfermedad.
H. R. KRIESEL (1930) en un estudio comparativo clínico e histopatológico del distemper canino y de la encefalitis experimental de los perros provocad¿ por el virus de la encefalitis de los zorros, encuentra que las observaciones de DE MONBREUN, clínicas y anatómicas son similares a las obtenidas por el. El virus empleado para la inoculación a los perros fue obtenido de un zorro que se llevó al laboratorio para examen. La investigación microscópica demostró las inclusiones nucleares típicas en el endotelio vascular del cerebro, similares a las referidas por GREEN y colaboradores. La inyección intracisternal de una suspensión de material cerebral probó ser fatal para un perrito, en 5 días. El comienzo de la enfermedad fué brusco, con elevación de la temperatura, inapetencia, inquietud, debilidad de las patas posteriores, abundante descarga mucosa de los ojos y de la nariz, descargas que se observaron durante las 24 a 48 horas, seguidas rápidamente de muerte. A la autopsia se comprobó la existencia de las inclusiones nucleares específicas, en el endotelio de los vasos cerebrales, en las células hepáticas y en las de Kupfer, en el retículo-endotelio del bazo, y de los ganglios linfáticos. La autopsia demostró además petequias y edema del timo, edema y hemorragias de los ganglios linfáticos y mesentéricos, hemorragias subendocárdicas, equimosis de los pulmones, serosidad del peritoneo. La inoculación intracisternal del virus provocó hemorragias en las leptomeninges del cerebro, mientras que la inyección intraperitoneal determinó marcada exudación fibrinosa en la cavidad abdominal. Una comparación de sus observaciones en ambas enfermedades revela que hay una marcada disimilaridad en el curso clínico de estas dos infecciones; que la patogenicidad de los virus productores de ambas infecciones es distinta respecto de los hurones experimental ni ente infectados; que los perritos infectados con el virus de la encefalitis epizoótica de los zorros muestran inclusiones nucleares en una proporción del 100 %, mientras que los infectados con el virus del distemper muestran una proporción de estos elementos en sólo 18 %; que la proporción de inclusiones intracitoplasmáticas en los perritos infectados con el distemper era de 63 %, en tanto que sólo en un perrito fué posible descubrir inclusión de esta especie, empleando el virus de la encefalitis de los zorros. Estas y otras comprobaciones conducen a KRIESLER a considerar la posibilidad de la existencia de una sola cepa de virus, cuya patogenicidad puede estar marcadamente aumentada para los perros y considerablemente disminuida para los hurones, la ocurrencia de infecciones espontáneas de los perros mediante el virus de la encefalitis de los zorros, y la existencia de alguna otra relación entre el virus del distemper canino y la encefalitis de los zorros.
RESULTADO DE NUESTRAS INVESTIGACIONES
El material orgánico de que nos hemos servido para este análisis histológico proviene de un perro de corta edad, que murió como consecuencia de una forma breve de la enfermedad, y con signos predominantemente nerviosos.
CORTEZA
Dos son los tipos de reacción meníngea hallados : uno reciente, predominantemente inflamatorio, con proporción igual de polinucleares y mononucleares grandes y medianos, ubicuo, carente de inclusiones celulares : figuras Nº 1, 2, 3, 4; y otro de tipo edematoso, corrientemente en los surcos, con el natural aflojamiento de las mallas de su componente conjuntivo, enturbiamiento de su substancia fundamental y escasa o ninguna presencia de elementos de infiltración: Fig. 5. Muy interesante es el aspecto de la Fig. 6, acerca de las concomitancias patológicas de elementos del mesenquima y del parénquima nervioso cortical. Se trata de una preparación lograda con el método argéntco de Río HORTEGA, en caliente, que permite ver la alteración reciente meníngea, de tipo inflamatorio, la respuesta gliocítica reaccional predominantemente astrocitaria de la corteza, que se advierte mejor en la Fig. 7, y el aspecto marcadamente fibroso del vaso aracnoideo y de sus ramificaciones corticales. En lo principal estas imágenes permiten afirmar el desarrollo de una anormalidad meníngea de tipo reciente y reacciones edematosas de la misma que bien pueden corresponder a momentos en sus cambios patológicos.
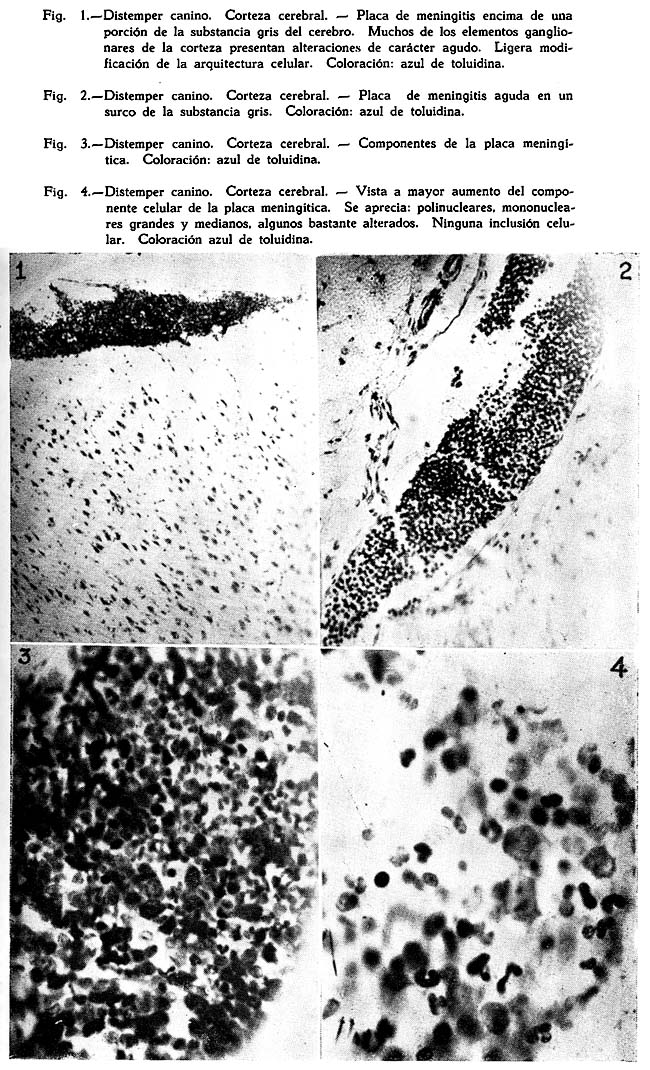
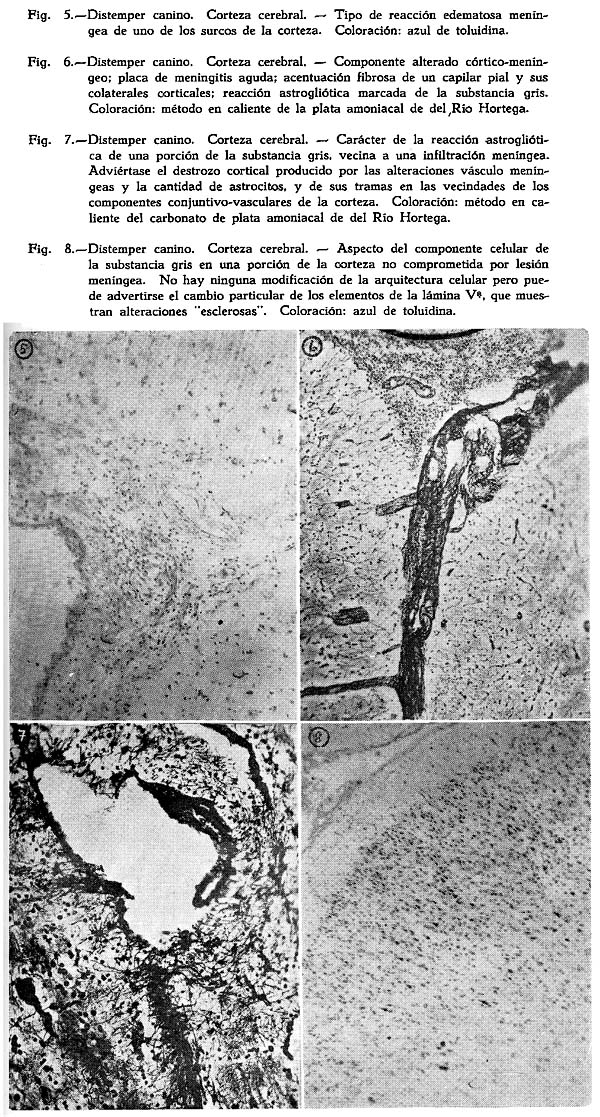
Las alteraciones en los elementos de la substancia gris son dignos dc anotar. Se trata unas veces de cambios notables de las células ganglionares que comprometen láminas o regiones circunscritas de la corteza; otras, de alteraciones difusas, pero que lesionan determinado tipo de elementos; y otras, de cambios, en cierta forma circunscritas, dependientes eveces de la distribución vascular. En la figura 8 se distingue con precisión la normal distribución de las células ganglionares en las diferentes capas de la corteza, sin el menor cambio arquitectural de sus componentes celulares, ni la presencia de vacíos celulares como acontece en otros lugares: Figs. 12, 17. Pero a poco que examinamos la referida Fig. 8 sobre todo con la ayuda de una lupa distinguiremos que muchos de los elementos de la lámina piramidal IIIa2, y IIIa3, esto es, los elementos piramidales medianos y grandes, lo mismo que los de la lámina Va, muestran cierta particular densidad en su cuerpo celular y cierto estado de agudeza en sus bordes, significativas de cambios estructurales de importancia. Este cambio se ve mejor en la Fig. 12. Modificaciones de esta especie testimonian un tipo de alteración celular sistematizado laminar, pues los cambios comprometen sólo las células de determinadas láminas dejando indemnes las de otras. A este especial tipo de alteración le hemos llamado sistematizado laminar.
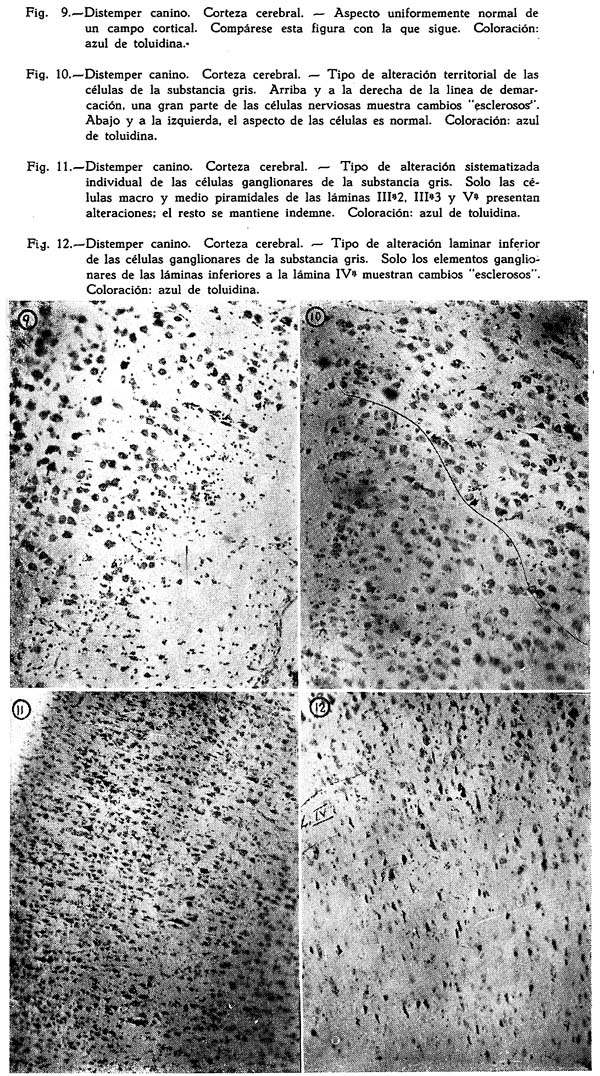
En otros sitios no ocurre este tipo de variación sino otro, segmentario, de menos conformidad con la distribución laminar, pero que compromete en cambio considerable número de elementos de toda una zona de la corteza, dejando a la vecina sin tocar. Esto es lo que se ve en las figs. 10 y 14. En la Fig. Nº 10, al lado derecho de la línea de demarcación se ve numerosas células modificadas en el sentido comunmente llamado escleroso; al lado izquierdo ninguno de los componentes celulares muestra cambio de ninguna especie. La Fig. Nº 9, de un territorio vecino al examinado, en el que no hay modificación ninguna en sus elementos, sirve de prueba comparativa para afirmar nuestro acerto. A este tipo de modificación le llamamos segmentario.
En algunos sitios el cambio ganglionar es predominantemente inferior a la capa granularis interna: Fig. Nº 12; en otros, superior a la misMa: Fig. Nº, 13. En ciertos sectores, además de la presencia de células en alteración fibrosa hay desaparición lagunar de las mismas: Fig. Nº 14: en otros hay verdaderas bandas de elementos alterados ganglionares, solos: Fig. Nº 15, o con intervención acentuada de elemento gliocítico: Fig. 16. En otros sitios el cambio esta vez predominantemente isquémico es marcado en las últimas hileras de células corticales de la lámina VI, vecina a la sustancia blanca: Figs. 17 y 19.
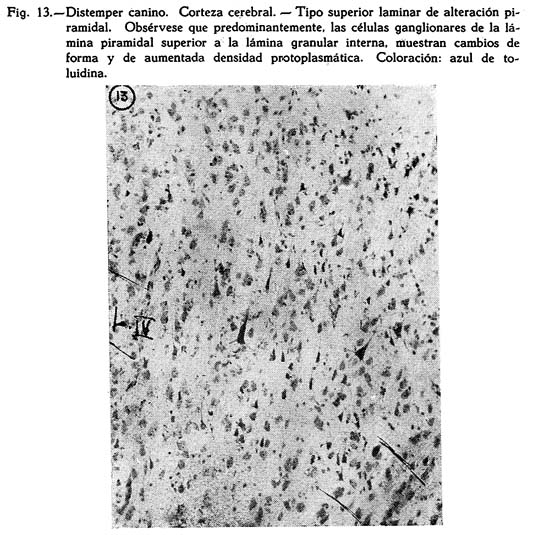
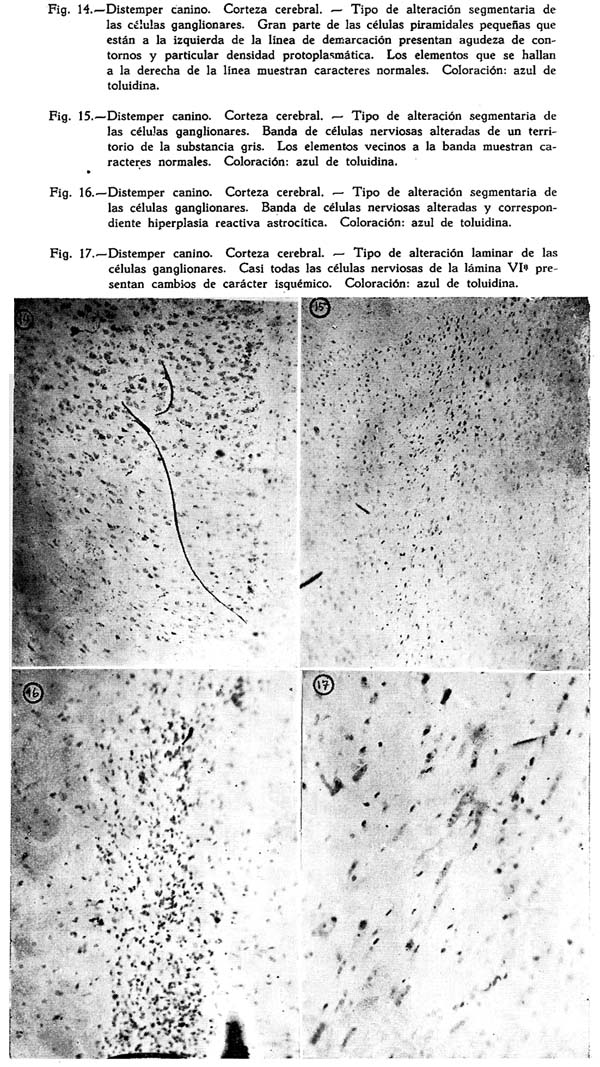
Tales tipos de alteración, en oportunidades de tanta conformidad como se vé con la estructura laminar de la corteza, corno ocurre con la lámina IV, o con demarcaciones territoriales definidas, se podrían explicar sólo por las cualidades tópicas de reaccionabilidad, inherentes a sectores o unidades celulares, circunstancia que anotó O. VOGT en el cerebro humano. Que tales circunstancias ocurren lo demuestra el hecho de la especial respuesta de la corteza en sus diferentes láminas, a un agente químico, el carbonato de plata en este caso, que se comporta de manera evidentemente diferenciada en las distintas láminas de la corteza: Fig. 20.
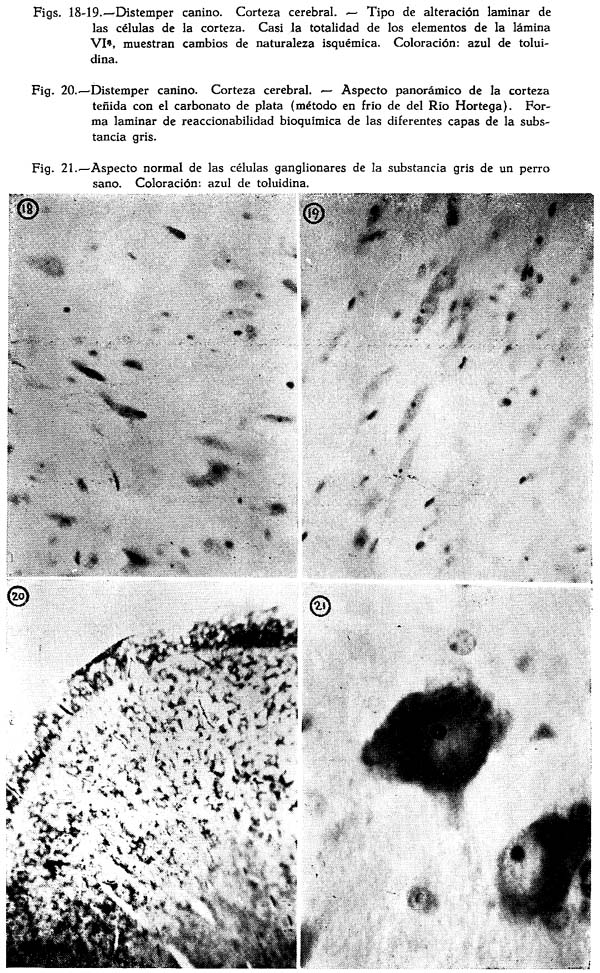
Las ilustraciones que siguen, han de servirnos para mostrar los considerables cambios individuales en la porción gris de la corteza. El tipo de la célula ganglionar normal de la corteza del perro sano es el de la Fig. 21. A pesar de su forma redondeada, no por esto comparable a las que van a servirnos de análisis, conserva todos los atributos de un elemento que podríamos considerar genérico, por la calidad y distribución normal de sus elementos constitutivos, y por la disposición de sus contornos, naturalmente mejor apreciados al examen microscópico. La célula ganglionar de la Vª lámina, Fig. Nº 22, rodeada por elemento sano en toda su extensión, muestra ya modificaciones que podríamos llamar iniciales: homogeneización y acentuación cromática parcial de su protoplasma, empequeñecimiento y tendencia a excentricidad de su núcleo, minimización nucleolar. La notable del conjunto celular aquí es que una sola célula es la afectada, y ya fuertemente afectada, y que el tipo de cambio es el mismo, primordial, de la alteración fibrosa que estudiaremos después.
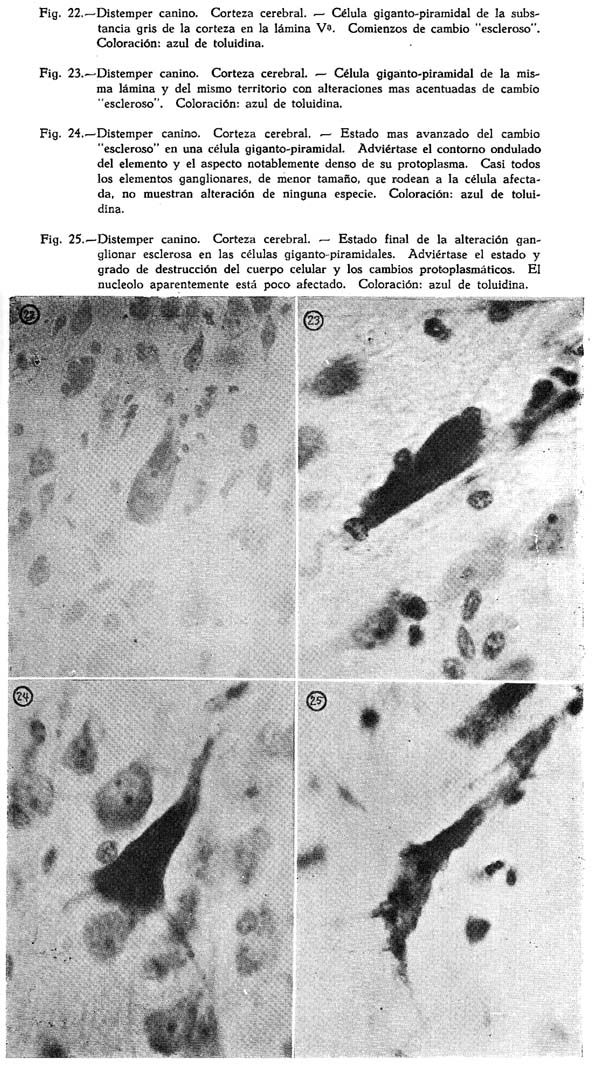
Los cambios son mas marcados en las células de las Figs. 23 y 27, tanto por las profundas modificaciones de su contenido protoplasmático, que van, desde el estado de acentuada homogeneización y densidad, hasta el de su destrucción, dándole el aspecto de masa esponjosa, Fig. 25, cuanto por la deformidad en sus contornos que se vuelven sinuosos, afilados o con prominencias inusitadas. Las alteraciones son aisladas unas veces, comprometen determinados individuos celulares: Figs. 23-27; en grupo otras: Fig. 28; o predominantemente en ciertas unidades de un componente estructural: Fig. 29, destacándose la anormalidad celular del resto, normal, de los elementos del contorno. La mayor parte de estos cambios son de carácter grave, por la magnitud y especie de las alteraciones individuales.
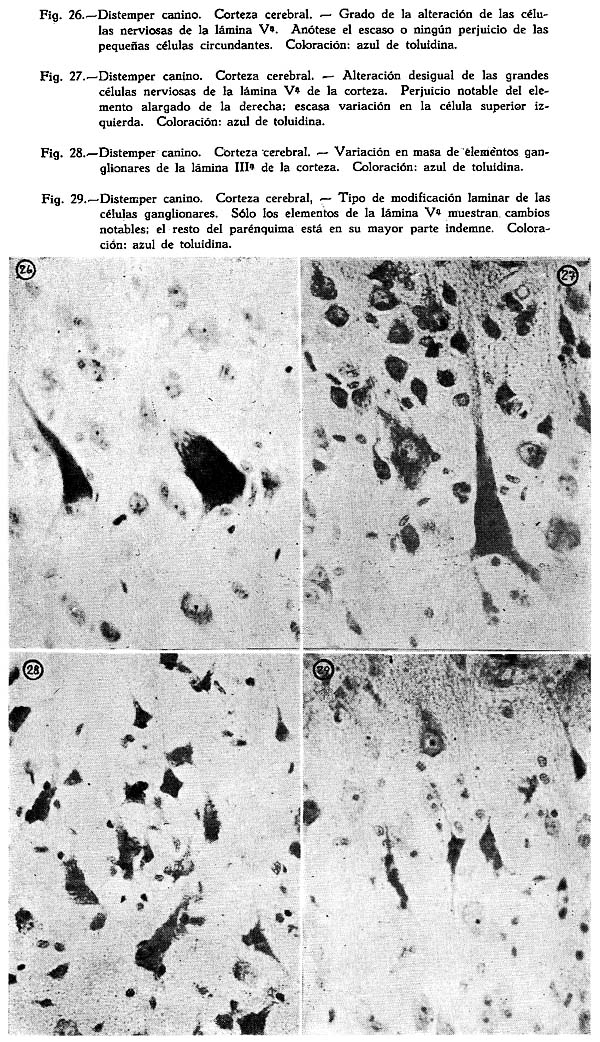
El componente astrocitario ha reaccionado fuertemente como lo demuestran las Figs. 6, 7, 33 y 34. Las dos primeras corresponden a porciones de la corteza con especial reacción gliótica astrocitaria concurrente con marcadas lesiones de la meninge; la Fig. 34, al especial tipo de astrocito, y su peculiar distribución en la substancia blanca.
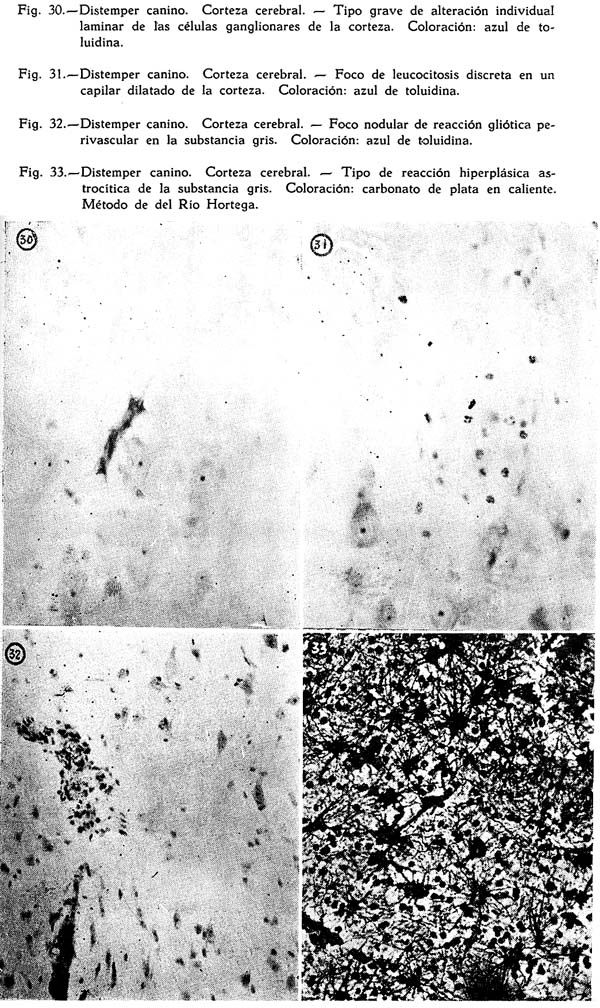

Casi la mayor parte de la corteza presenta las reacciones que acabamos de anotar, en unos sitios mas que en otros, y tanto en la porción blanca como en la gris, Además hay proliferación vascular de la misma y la consiguiente formación de manguitos de infiltración poli y mononucleares, pericapilares, que se desarrollan como expresión del cambio reaccional inflamatorio de la enfermedad.
CUERNO DE AMMON
Para darnos cuenta de la ubicación de las alteraciones que vamos a describir consideramos en esta zona las siguientes porciones: las flechas verticales de la parte superior de la Fig. Nº 35 muestra la separación de este territorio en dos campos, uno a la derecha que corresponde al campo piramidal subicular simple, y otro a la izquierda, al piramidal ammónico: HE1α y HE2 de la carta de von Economo, respectivamente. En la parte inferior de la fotografía se vé la fascia dentata f. d., dividida mediante una flecha en dos zonas, una superior x, correspondiente a una porción del cuerno de Ammon cubierta por las formaciones vecinas, y otra inferior, correspondiente a la porción descubierta de la misma, en relación con los espacios subaracnoideos, y por ésto bañada con el líquido circulante en ellos. Además en el codo de la porción terminal de la lámina piramidalis, campo HE2, de von Economo, señalamos dos porciones a, y b, a las que nos referiremos en el curso de esta descripción.
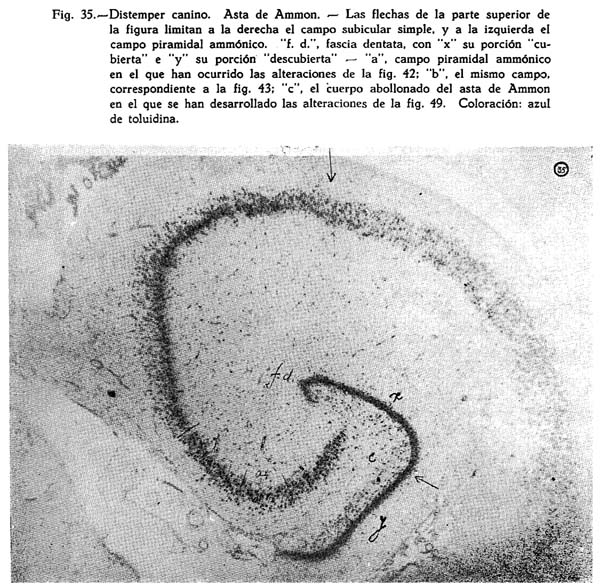
Todas estas zonas son asiento, de alteraciones celulares individuales, aislados, o en grupos. Las Figs. 36 y 37 señalan modificaciones totales en los componentes celulares de la lámina superior de uno de los campos de la formación ammónica. No sólo elementos celulares aislados han quedado comprometidos, o masas íntegras, sino que su disposición citoarquitectural ha quedado perturbada, por la presencia de vacíos celulares en los que todo elemento neuronal ha desaparecido, o por la penetración abundante e inusitado de células glióticas. Así en la Fig. 38 se nota claramente cómo las células ganglionares, justamente de la agrupación uncinada a, muestran particulares cambios estructurales, en tanto que la formación b subyacentes conserva sus características normales. Además el espacio normalmente existente, estrechísimo, entre agrupaciones uncinadas vecinas, y porciones subyacentes, está aquí considerablemente ensanchado, y sobre esto, vacío,libre de elemento neuronal. En ocasiones, como en el caso anterior de la Fig. 38, se trata sólo de modificaciones en las células nerviosas ganglionares, sin participación de elemento gliótico; en otras en cambio, al lado de alteraciones marcadas y graves en aquellas, se encuentra proliferación de gliocitos, muchos de los que rodean a las neuronas, en plena neuronofagia: Fig. 39, dando al conjunto neurogliótico una apariencia nodular a, que se destaca por su coloración generalmente intensa, y por su composición, del resto del cuadro. De la magnitud de las lesiones anteriores puede darnos idea la Fig. 40, tomada con mayor aumento de una de las láminas anteriormente examinadas, en la que puede apreciarse el grado profundo alterativo de las células nerviosas en esta zona. Prácticamente se nota aquí, un cambio total de los elementos de un territorio cortical. Conviene recordar que se trata, en este caso, de porciones del órgano en relación inmediata con los espacios subaracnoideos y por esto enteramente bañados por el liquido cefálico circundante.
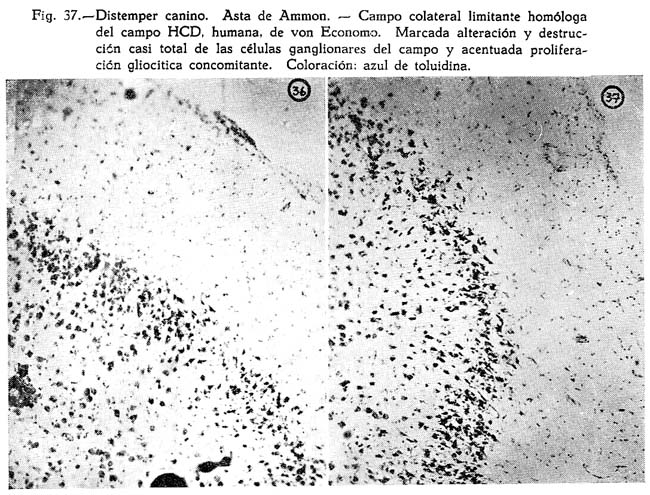
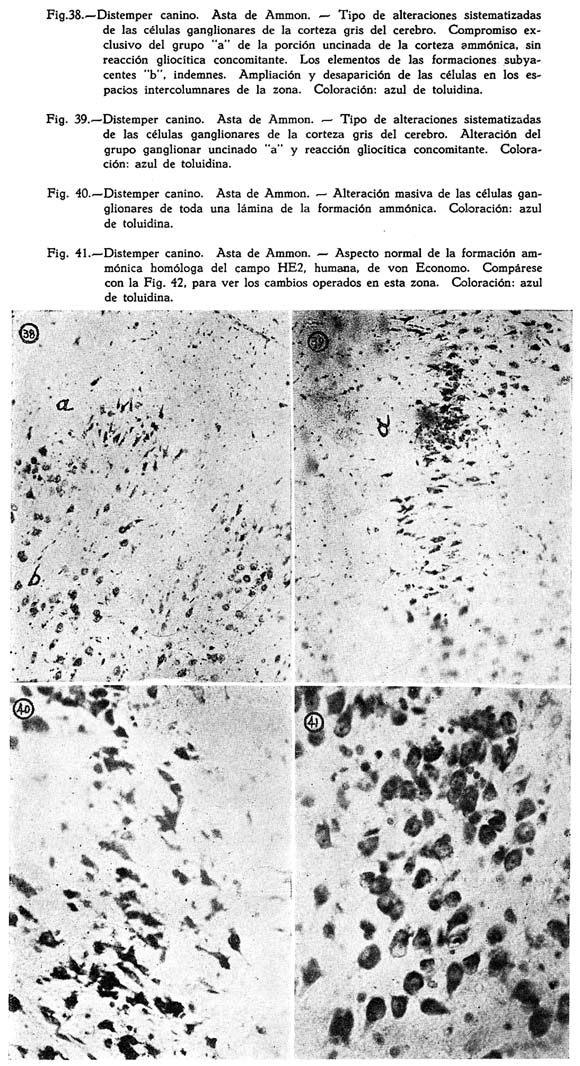
Las ilustraciones que siguen muestran otro tipo de alteración, esta vez individual, en ciertos de los territorios y láminas de la formación ammónica. La Fig. 41 correspondiente a una zona del campo HE2. de von Economo muestra una zona normal de la región; la Fig. 42, vecina a la anterior, y correspondiente al mismo campo, tiene alterados muchos de sus elementos. Se advierte, en efecto, la presencia de numerosas células alargadas, de contornos agudos, de cuerpo protoplasmático denso, las extremidades de algunos de los cuales, penetran, a manera de agujas en las capas inferiores de la corteza. El contraste no puede ser mayor al comparar una imagen con la otra. Pero la comprobación es aun mas evidente al examinar las Figs. 43 y 44. La primera corresponde a una porción normal de la corteza ammónica de determinada zona; la segunda, a una porción alterada, Claramente se vé cómo los elementos de la primera carecen de toda significación patológica, y como, en cambio, en la segunda están, algunos, acentuadamente deformados, en formas serpiginosos, lanceoladas u ovoides, dejando a las del rededor, casi enteramente normales en su forma, aunque ya, se advierte en éstas, en algunas, particular densificación parcial protoplasmática, y cambios de contornos preliminares.
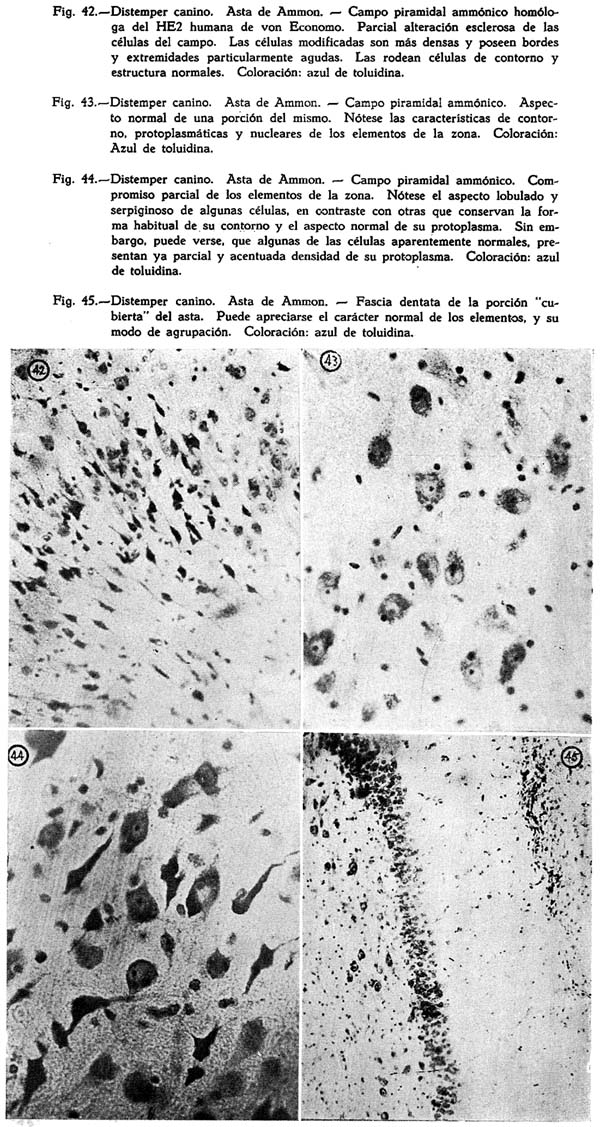
Veamos ahora los cambios en la fascia dentata. La Fig. Nº, 45 muestra la porción cubiertade esta zona, esto es, aquella inmediata a las formaciones vecinas, sin solución de continuidad ninguna. Todas las células, uniformemente, se presentan con caracteres normales. La Fig. Nº 46, en cambio, correspondiente a una porción de la fascia dentata libre esto es en relación inmediata con el espacio subaracnoideo, y por esto bañada con los líquidos circulantes de la cavidad craneana, muestra una imagen distinta. Una gran parte de los elementos se presentan particularmente encogidos, de bordes agudos, de un aspecto enteramente distinto en fin al de los pocos normales circundantes.
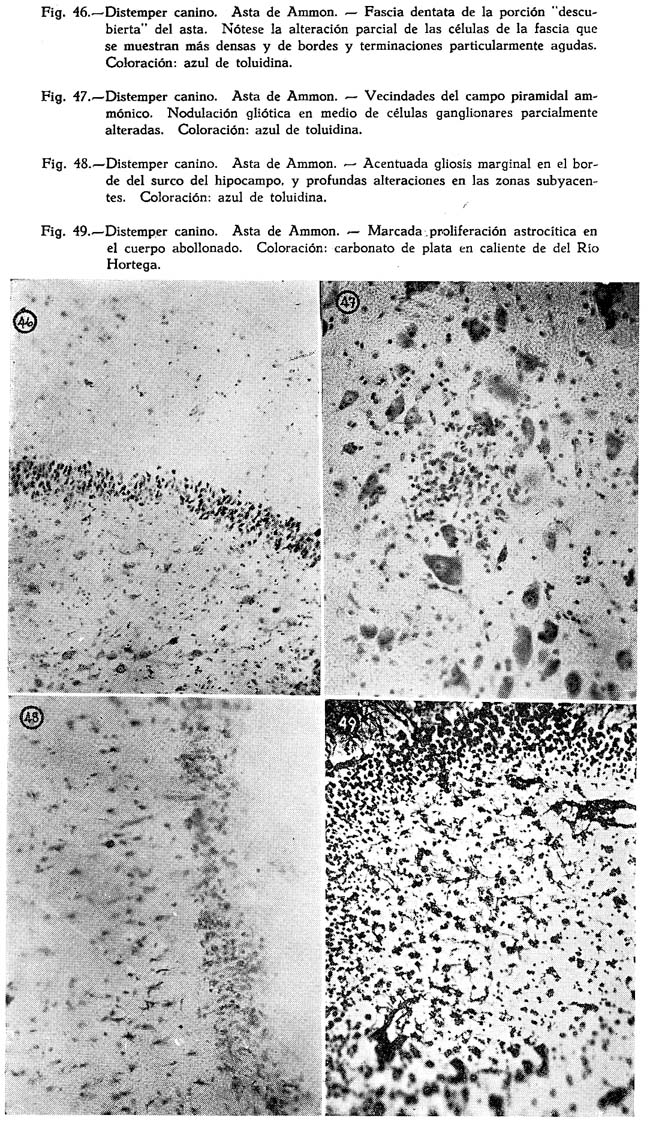
El agente de la enfermedad, en este caso de la fascia dentata como en las formaciones de Ammón anteriormente estudiadas, ha tenido la particularidad de lesionar profundamente, a sólo determinados elementos de un territorio, dejando al resto indemnes. El hecho de que sea justamente la zona en relación con el liquor la particularmente comprometida, en contraste con la porción cubiertaque no ha sido tocada, justifica la presunción de que sea el liquor el medio actuante patológico, alterador de las células nerviosas, por la acción de algún agente, bacteriano, virulento o tóxico, encerrado en su seno. Mas esta explicación, clara para el caso de la totalidad de la formación comprometida no justifica el compromiso particular y aislado de las células. De la zona alterada, algunas, sólo, sufren manifiestos cambios estructurales, el resto en cambio permanece indemne o ligeramente alterado. Esta modificación tan visible en el estudio de las formaciones ammónicas, que apoya con toda precisión la Fig. Nº 44, solo puede explicarse o por una vulnerabilidad particular elemental de las células nerviosas frente al factor patogénico actuante, o por un estado de peculiar meiopragia celular, o como uno de los tantos momentos del cambio anormal anatómico. Sobre este particular conviene recordar la extremada vulnerabilidad del asta de Ammón en los procesos neurológicos, tanto en la especie humana como en la animal inferior, como tuvimos oportunidad de comprobarlo en las alteraciones producidas por la intoxicación estrícnica y otras. Que la vulnerabilidad es en efecto, particular en el asta de Ammón, lo demuestran en este caso las imágenes dé nodulaciones glióticas en pleno seno de formaciones ganglionares: Fig. 47; las marcadas gliosis marginales y el perjuicio grave de las células gangliona res de las láminas subyacentes: Fig. 48; las reacciones fuertemente astrocitarias del campo abollonado HF de von Econono: Fig. 49; y las manifiestas reacciones de infiltración perivascular en las vecindades de un gran vaso de la región: Fig. 50. Son pues evidentes y particulares, por la calidad de los perjuicios ocasionados, sea en las unidades estructurales como en las celulares, las alteraciones del asta de Ammón en el caso en estudio así como los moldes de reaccionabilidad de los elementos que la integran.
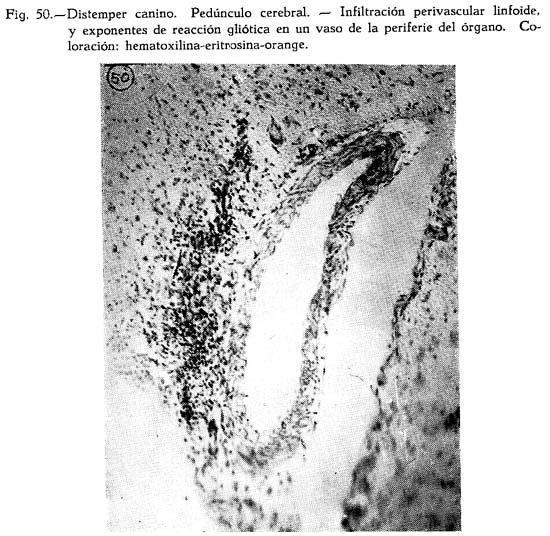
TÁLAMO
Manifiesto engrosamiento y discreta infiltración de la membrana de la pía que recubre el tálamo en su superficie lindante en el IIIer ventrículo, lo mismo que infiltraciones nodulares subpiales a base de elementos linfoides, y concomitante aparición de gliocitos en la masa misma del parénquima, en sus láminas más superficiales, con perjuicio marcado de las células ganglionares de casi toda la zona, en particular de aquella que está mas en relación por las porciones afectadas por la infiltración es lo que con frecuencia se halla en el tálamo: Fig. Nº 51. Otras veces, imágenes reccionales a base de elemento gliocítico, corrientemente perivasculares o también difusas, en porciones mas profundas de la formación, como ocurre en la Fig. Nº 52. En otras, se aprecia el compromiso perivascular de pequeñísimos capilares rodeados de células de infiltración de tipo linfoide como en la Fig. Nº 53. Como resultado de estos cambios se comprueba la existencia de alteraciones profundas en los elementos ganglionares del tálamo, en todos sus segmentos, predominantemente en los vecinos a la cavidad del IIIer. ventrículo. Unas veces tienen las neuronas aspecto retraído, denso, vacuolar o reticular, con manifiesta deformación de sus cuerpos, que los conduce hasta presentar imágenes serpiginosas, ondulados, aciculares, etc., como en la Fig. Nº 54. En otras, el aspecto celular, de suyo bastante deformado, tiene un especial carácter : agudeza de los bordes celulares y extremada finura, casi filiforme de sus prolongaciones, con desaparición, además, de muchos elementos y su sustitución por acúmulos glióticos reccionales de mayor o menor densidad: Fig. Nº 55. Es frecuente a veces el hecho de la aparición de modificaciones especiales celulares en el sentido de deformaciones aciculares alargadas, filiformes que acompañan a los vasos, o corren independientes de: ellos: Fig. Nº 56; o la presencia de deformaciones considerables y modificaciones protoplasmáticas correspondientes como las de la Fig. Nº 57. No es infrecuente por último el hecho de encontrar vasos pequeños de paredes ligeramente engrosadas con reacciones especiales de sus componentes adventiciales o endoteliales: Fig. Nº 58. En resumen, en el tálamo se hallan expresiones de Infiltración difusa o circunscrita, predominantemente en las vecindades de marcados perjuicios meníngeos, o vasculares, y como resultado, diversidad y gravedad de alteraciones de las células ganglionares muy en relación con el tipo del elemento propio de la zona.
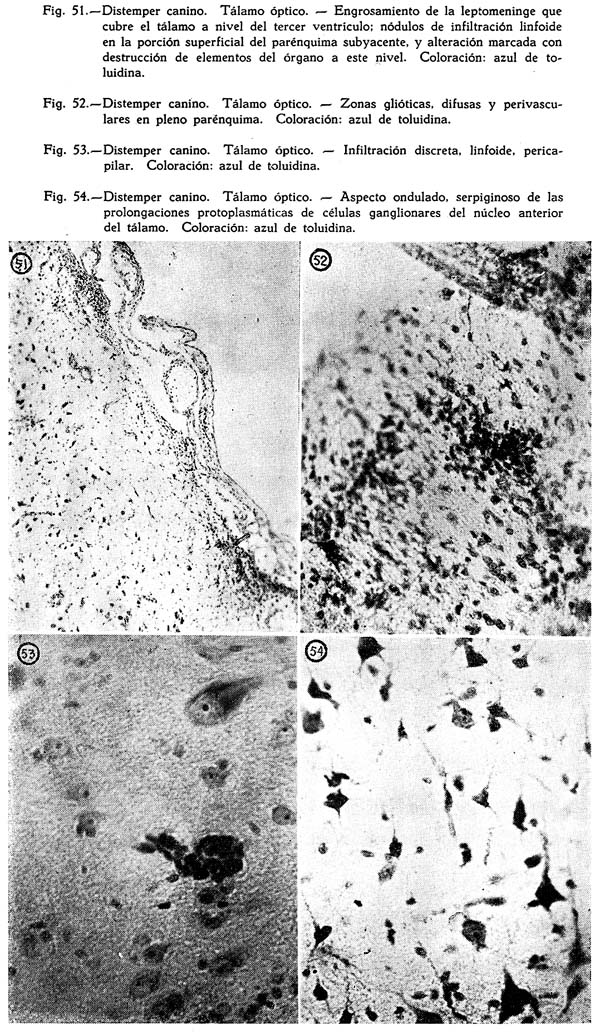
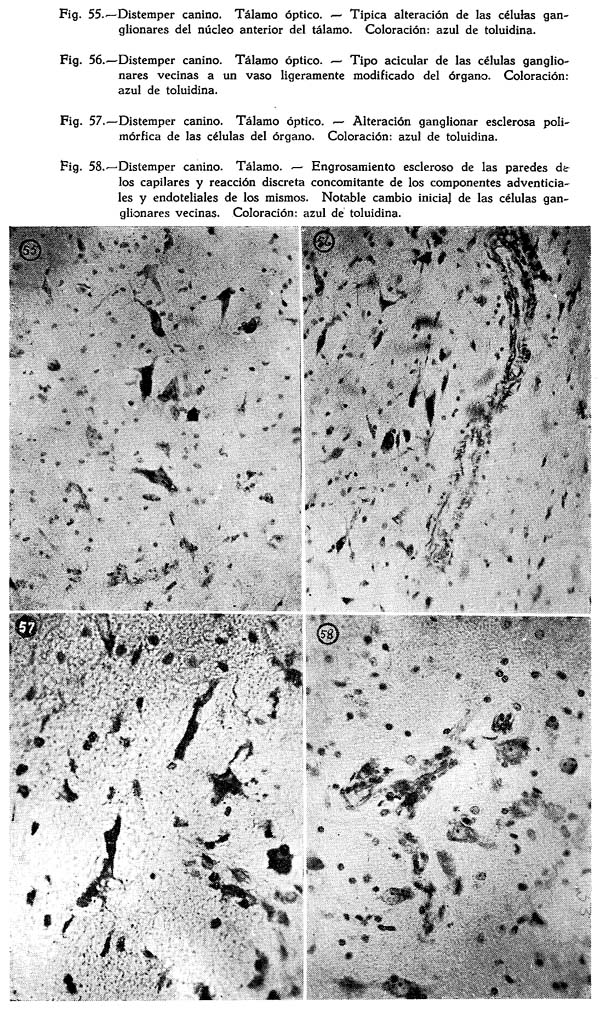
PEDÚNCULOS CEREBRALES
Nódulos gliocíticos circunscritos, Fig. Nº 59, o zonas de gliosis difusas, más o menos extensas, unas veces en relación con vasos enfermos, otras nó, Fig. Nº 60. Reacciones marginales glióticas de trama densa y de composición especial por la presencia de unidades astrocitarias pequeñas y especiales, Fig. Nº 61, es lo que corrientemente se aprecia en porciones de esta formación vecinas a la periferie, o algo profundas. En otros lugares, se halla expresiones de alteración vásculo-parenquimatos de diversa índole. Así no es raro encontrar imágenes reactiva-vasculares plasmocíticas y linfoides, periféricos a los capilares, en cuya composición intervienen, reaccionalmente, unidades propias nerviosos, gliocíticas predominantemente, Fig. Nº 62. En otras ocasiones: Fig. Nº 63, la reacción adventicial es plasmocítica, con alguno que otro elemento de cepa linfoide. En oportunidades lo que se observa no es ya la infiltración adventicial anotada, sino sólo, la reacción proliferativa celular de los componentes propios de los capilares: células adventiciales y endoteliales: Figs. Nº 64 y 65; y, en otras, hermosas formaciones en ramillete o agrupaciones como en las Figs. Nº 66 y 68 en cuya constitución intervienen elementos vasculares del capilar, hipertrofiados e hiperplasiados, y células plasmáticas, en conjunto difuso, o mas o menos circunscrito a los trayectos vasculares. Las alteraciones celulares ganglionares se traducen, sea por alteración en grupo, de células ganglionares vecinas a la periferie y a zonas meningíticas de infiltración, adoptando entonces formas agudas y afiladas particulares: Fig. Nº 69, uno de cuyos elementos fotografiado en grande, muestra la Fig. Nº 70, o a cambios individuales de mayor magnitud, en su protoplasma y núcleo, que los conduce a profundos cambios degenerativos: Fig. Nº 71. En resumen evidencia, en el pes pedúnculi de procesos infiltrativos difusos o circunscritos, linfocíticos o linfoides, o plasmáticos, en diversas zonas, con el daño consiguiente y diverso de células ganglionares, cualquiera que sea su calidad y la consiguiente reacción hiperplásica e hipertrófica de los elementos propios a los capilares, reacción que se traduce en la formación de ramilletes celulares perivasculares, a base de células adventiciales y endoteliales y de elementos de infiltración plasmática o linfocítica. Respecto de la presencia de las células ganglionares de extremidades punteagudas o casi filiformes como las de las Figs. Nº 56 en el Tálamo, y las de las Fig.Nº 70 en el pedúnculo, es muy sugestiva su aparición en las vecindades de alteraciones periféricas meningeas, o en la de vasos en alguna forma modificados. No sería extraño por lo demás que se tratara de formas especiales de elementos ganglionares propias de la zona en cuestión, en animales de la especie del que estamos estudiando.
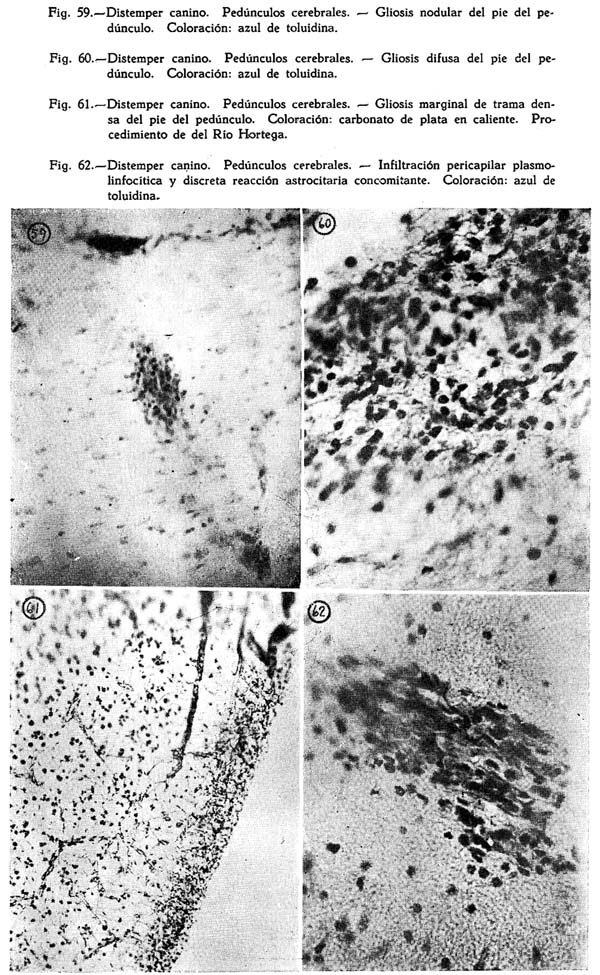
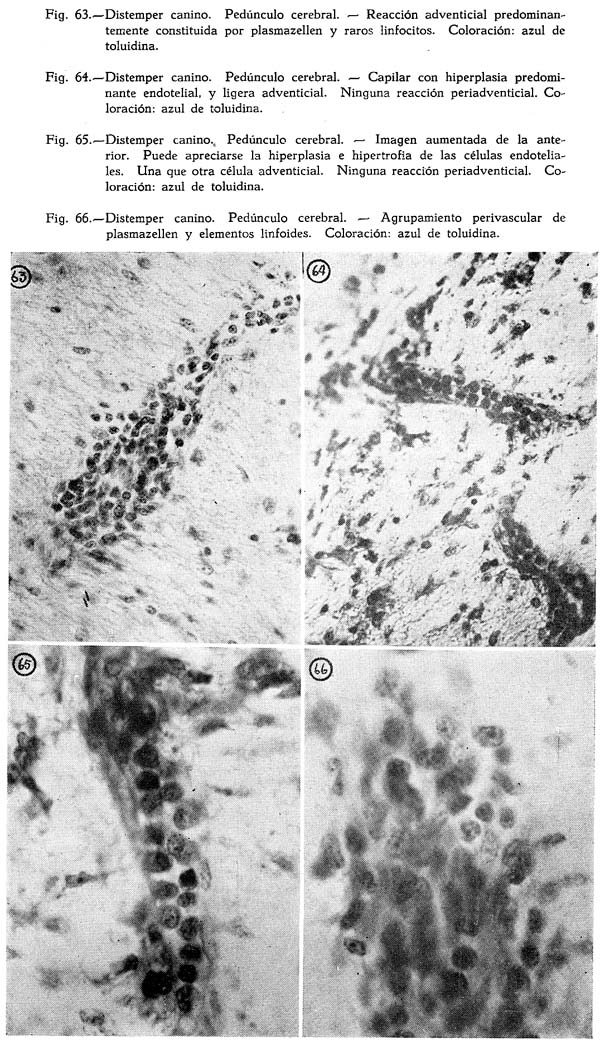
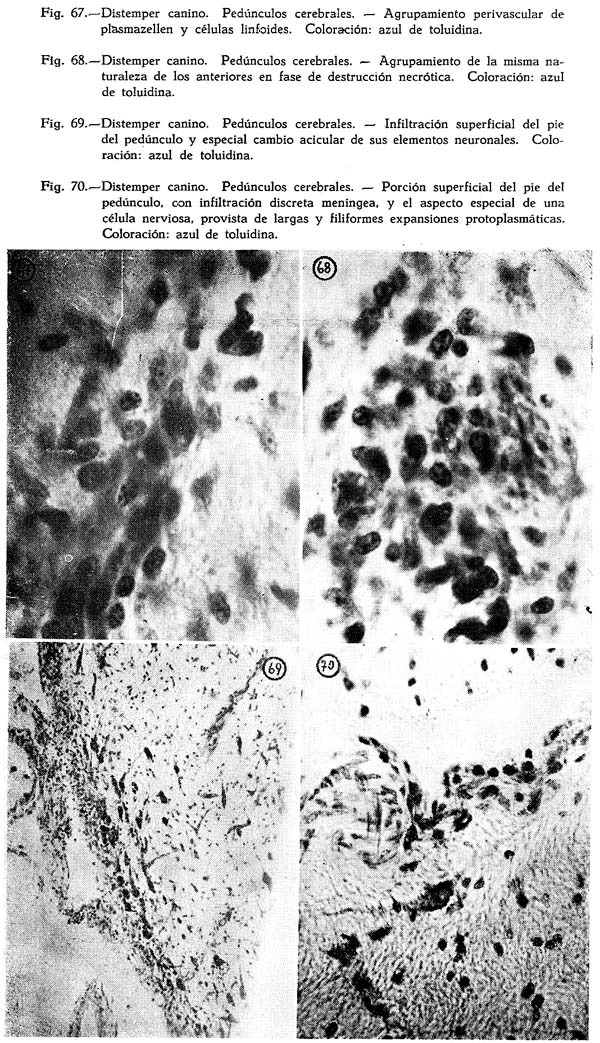
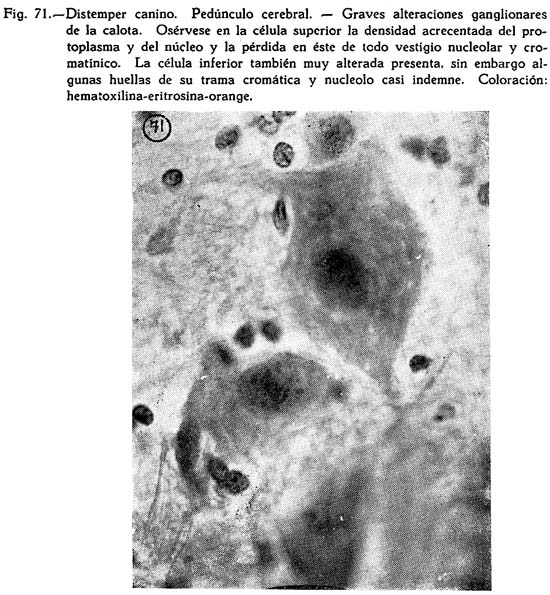
CAVIDADES INTERNAS DEL CEREBRO
La Fig. Nº 72 muestra una porción de una cavidad interna del cerebro en la que su epitelio ependimario es normal, tanto por la calidad de sus elementos, como por la forma de su agrupamiento, y estado de la lámina subependimaria subyacente. En la Fig. Nº 73 se aprecia ya un hecho patológico notable: la presencia de abundantes elementos de infiltración linfocítica o linfocitoide que han formado un nódulo, justamente debajo de la membrana ependimaria, indemne, sin provocar grande reacción en el componente subependimario o en las zonas vecinas. Lo curioso es que es discreta, sino nula, en algunos sitios, como en el fotografiado, la alteración de las células de la cubierta ependimaria, y, ausente. enteramente, toda expresión de manifestación reaccional en la cavidad que tapiza. La Fig. Nº 74 muestra, en otro sitio bajo una cubierta ependimaria también indemne una porción subependimaria infiltrada por elementos linfocitoides y plena de componente gliocítico reaccional. La reacción sin embargo es limitada justamente al límite inferior de la lámina sub-ependimaria. Trátase pues, aquí, de nódulos reaccionales, circunscritos, de distribución particular, según que ocupen el espacio subepitelial inmediato, o la porción subependimaria propiamente dicha. En otras ocasiones, la manera reaccional de los tejidos es distinta, difusa, tal como se vé en la Fig. Nº 75; o marcadamente, reactiva, exudativa, proliferativa como representa la Fig. Nº 76, de la cubierta ependimaria del fornix. El papel de los agentes patológicos parece que hubiera sido en este sitio particularmente intenso, según se deja colegir por los cambios anotados.
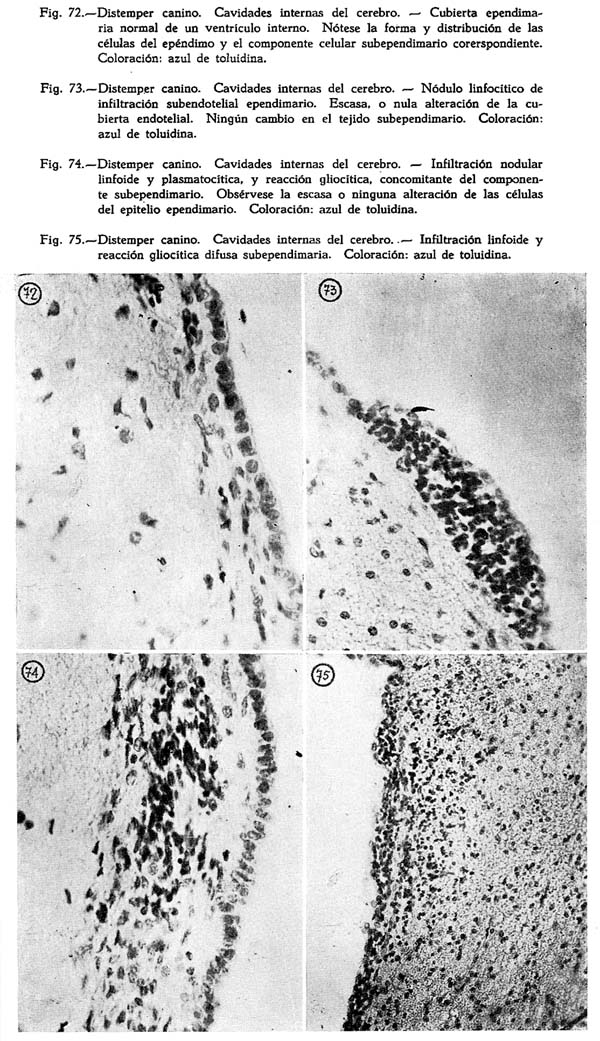
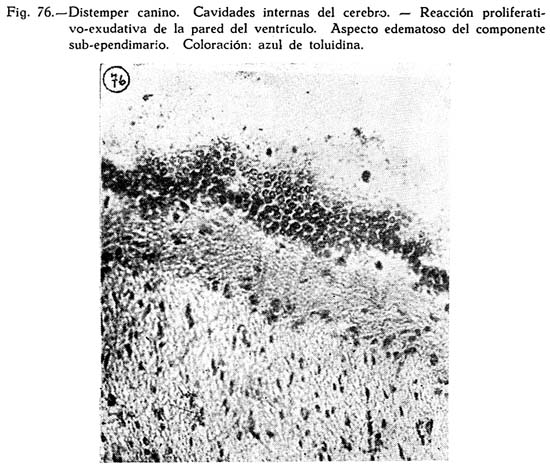
CEREBELO
Frecuentes son las porciones del órgano en sus láminas periféricas, comprometidas, por lesiones meníngeas de vecindad. Pero lo interesante es el desarrollo de alteraciones, directamente, en relación vascular. Una vista de una hojuela cerebelar Fig. Nº 77, muestra un trayecto vascular que desde la periferie avanza hacia la capa de los granos, en la cual es visible la destrucción de éstos en la inmediata vecindad del vaso penetrante, y la formación en a (Fig. Nº 77) de una zona de infiltración difusa en plena porción blanca del órgano. Las alteraciones en ésta, siempre de carácter vascular, son en otros sitios, en cierta manera circunscritos a los trayectos vasculares: Fig. 78; inmediatamente en relación con la cubierta de los mismos, constituyendo formaciones apendiculares linfocitarias y plasmocíticas: Fig. 79; o nódulos de estos mismos elementos, rodeando, en manguitos densos a vasos también lesionados en su endotelio: Fig. 80. Grandes zonas de infiltraciones, y necrosis circunscritas o difusas, en relación o nó con vasos, aparecen frecuentemente en la porción de los granos del órgano, destruyén dolos totalmente: Figs. 81-82. Como consecuencia, no es infrecuente encontrar expresiones de reacción gliótica, como las de la Fig. 83 donde se aprecia típicas formaciones de ramilletes glióticos, gliastrauchwerk de Spielmayer. Una vista a distancia como la de la Fig. 84 nos informa ya respecto de las variaciones de las células de Purkinje. Se advierte en ellas, además de las modificaciones en su cubierta nuclear, discretas, cambios mayores en su protoplasma. Células de Purkinje, normales, de un perro sano están fotografíadas en las Figs. Nos 85 y 86: se nota, sobre todo en la última, las características de forma, protoplasmáticas y nucleares, típicas, y el aspecto de los elementos corpusculares de la porción granulosa vecina. Cambios profundos en las mismas, del perro con distemper, pueden apreciarse en las Figs. Nº 87-90. La célula de la Fig. 87, considerablemente deformada, muestra una acentuada densidad protoplasmática en su parte inferior, y modificaciones de homogeneidad fluída de su porción superior, en un cuerpo celular de contorno irregular y difuso. Una apariencia pulverulenta fina o reticular, con homogeneización evidente del contenido protoplasmático, engrosamiento acentuado de la cubierta nuclear, y aspecto obscuro y denso de la cromatina es lo que se ve en la célula de PURKINJE de la Fig. 88. Mayor modificación por la deformidad del cuerpo celular y el aspecto reticular, irregular, denso, del protoplasma, unido a evidentes modificaciones de la cromatina. y acentuación de la cubierta del núcleo es lo que se nota en la célula de PURKINJE de la Fig. Nº 89. Elemento profundamente alterado, retraído, homogeneizado, en picnosis nuclear es lo que ofrece la Fig. Nº 90. En resumen imágenes de infiltración perivascular con predominio de elementos linfoide; extensas zonas de necrosis, formación de figuras de ramillete gliótico, gliastrauchwerk de SPIELMAYER, y notables modificaciones de las células de PURKINJE es 10 que se nota al examen del cerebelo.
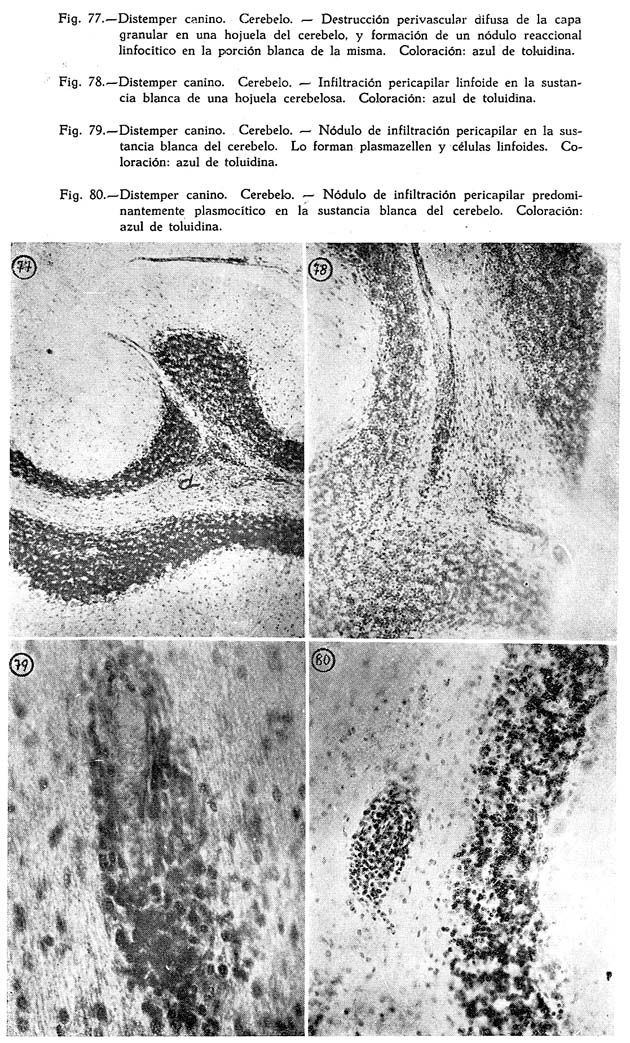
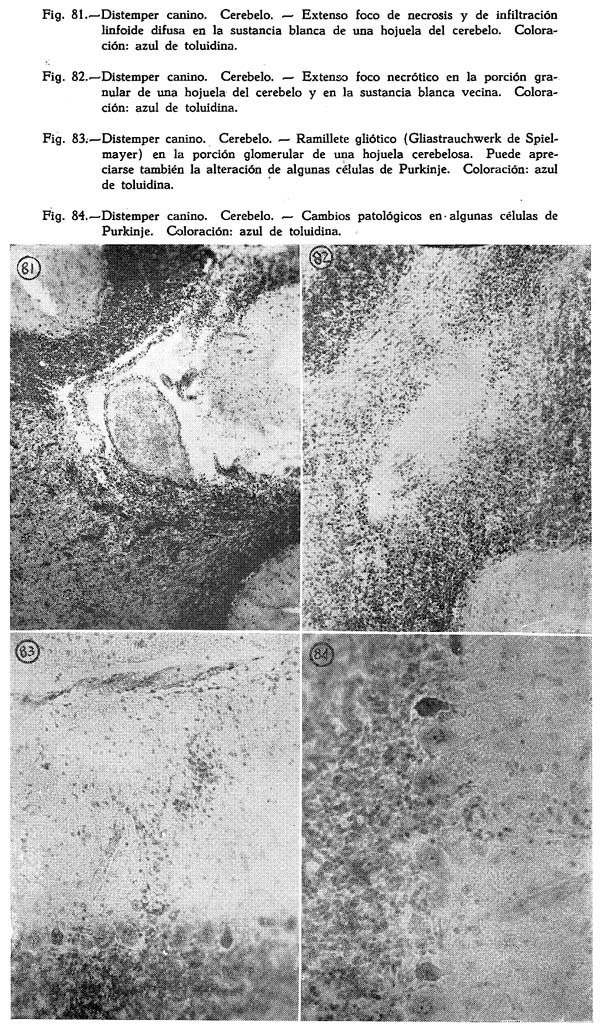
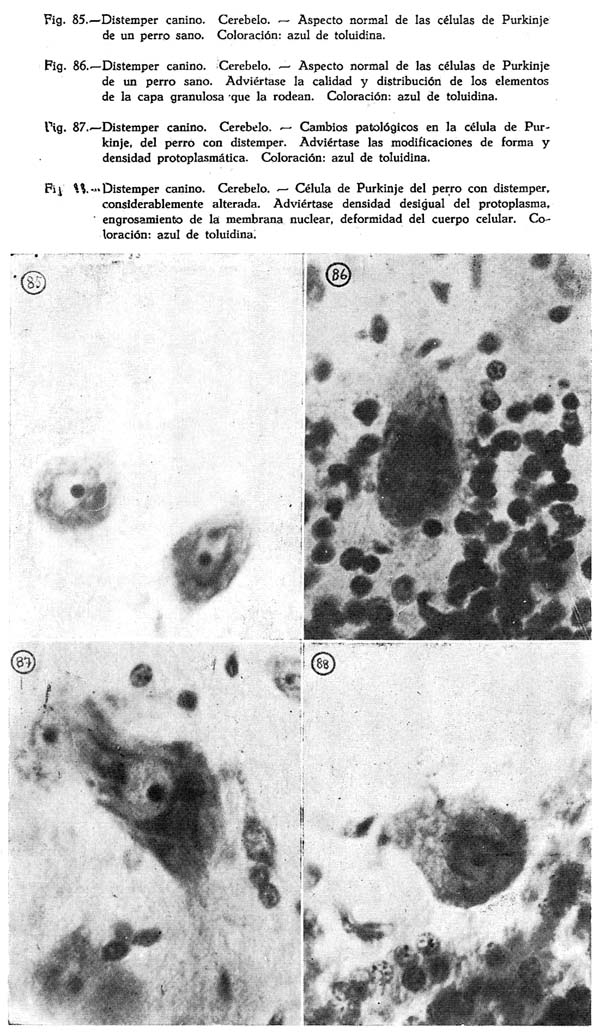
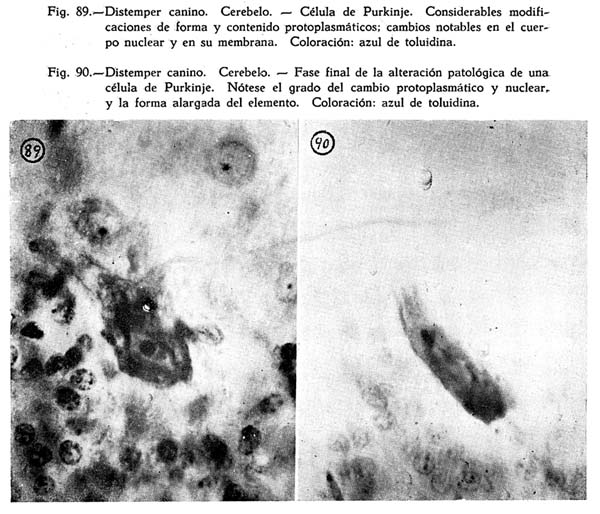
MÉDULA ESPINAL
Una vista panorámica del asta anterior de la médula, Fig. Nº 91, nos da ya materia para sospechar la especie y grado de las alteraciones estructurales de sus componentes. En efecto, en la Fig. Nº 92 puede apreciarse, de un lado, la alteración del epitelio ependimario del surco anterior del órgano, que muestra lesiones de carácter irritativo-exudativo, con modificaciones y soldadura mútua de los elementos que lo forman, y de otro, el carácter notable de las infiltraciones vasculares, y sus respectivos cambios endoteliales. El hecho patológico vascular no se reduce a la infiltración, y a las endotelitis anotadas, sino, a una alteración mayor, total del vaso, y al consiguiente perjuicio de la zona en la que se halla insertado, o riega. Es así corro en el vaso de la Fig. Nº 93 por ejemplo, podemos anotar un particular enturbiamiento y mayor densidad de la pared vascular, tumefacción e hipercromatosis del endotelio, y un especial estado de aflojamiento del parénquima nervioso, en donde la trama habitual, suelta, engarza apenas, cuerpos de glía, o de neuronas de pequeño tamaño, las que, además están alteradas. En la Fig. Nº 94 se destaca el típico manguito, linfocítico, o linfocitoide, pericapilar. La luz vascualr está estrechada, tanto por la densa masa circundante de los mencionados elementos, cuanto por los cambios del endotelio. La reacción gliótica es unas veces nodular pequeña, independiente de trama vascular: Fig. Nº 95; otras veces, siguiendo cursos capilares: Fig. Nº 96; otras, difusa, en zona mas o menos extensa Fig. Nº 97.
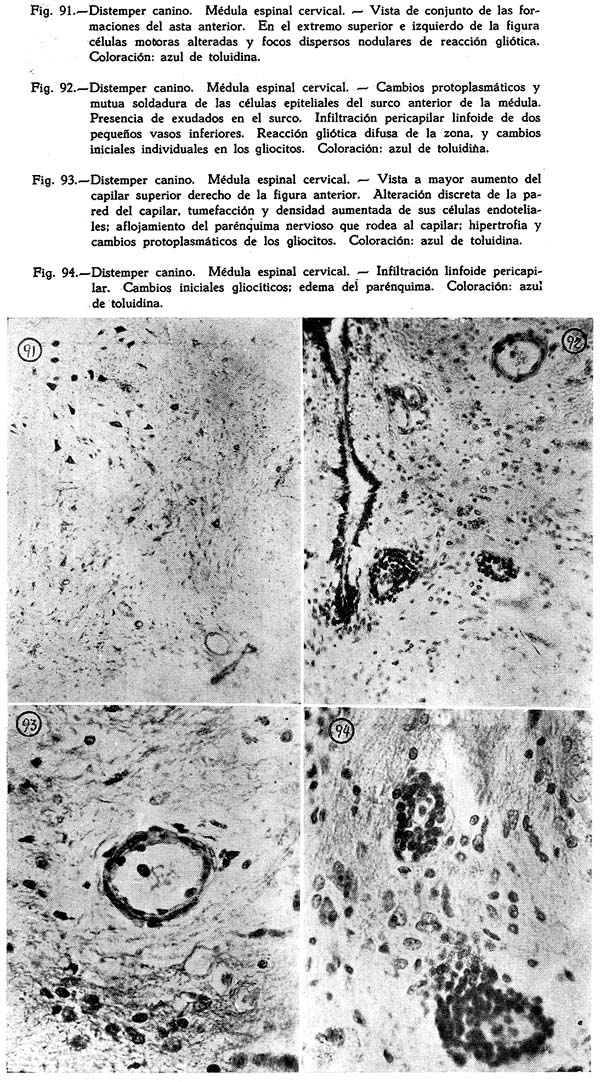
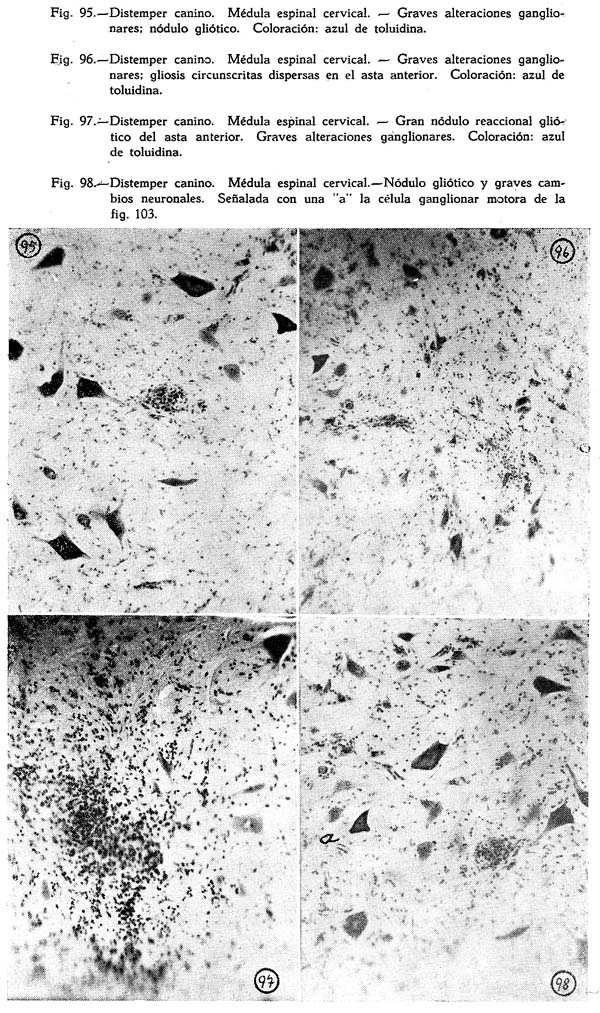
Las Figs. Nº 98 y 99, muestran las alteraciones de las células ganglionares vecinas a las zonas de infiltración o de reacción gliáticas. En la Fig. Nº 98, marcada con una a se halla la misma célula de la Fig. Nº 99, señalada con idéntica letra en la Fig. Nº .103. Estas imágenes permiten medir de antemano los cambios neuronales del órgano. La Fig. Nº 100 muestra la imagen de una célula nerviosa del asta anterior de la médula espinal de un perro sano.: puede apreciarse el aspecto de sus contornos, las formaciones protoplasmáticas que le dan el típico aspecto tigroide; la forma y caracteres de la red cromática, su nucleolo, el aspecto de su engarce en el tejido ambiente. La Fig. Nº 101 del perro enfermo, muestra ya un estado marcado de cambio patológico: se halla desprendida de su contorno, en parte; su protoplasma se ha homogeneizado totalmente, perdiendo el carácter tigroide que le fuera característico, con cierto estado de mayor fusión en la parte central del cuerpo celular; su núcleo enteramente excéntrico presenta un estado de densidad parcial de su cromatina, bajo la forma de bloques irregulares que se destacan del resto del núcleo, claro y transparente. El tejido nervioso de la periferie está manifiestamente laxo. Un aspecto semejante, con mayor desprendimiento del tejido ambiente, presencia de bloques pequeños de cromatina nuclear y pérdida absoluta de las inclusiones protoplasmáticas típicas es lo que se ve en la Fig. Nº 102. La densidad protoplásmica es mayor, la pérdida de los afilados contornos del cuerpo y su sustitución por límites contorneados irregulares y difusos, la acentuada densidad de la cromatina nuclear y el manifiesto aumento de volumen del nucleolo, es lo que tiene el elemento de la Fig. Nº 103 notablemente alterado. Resto sólo de unidad histológica, cadáver casi celular, enteramente desprendido del tejido ambiente, con desaparición de las características habituales de célula nerviosa, es lo que se ve en el elemento fotografiado de la Fig. Nº 104.
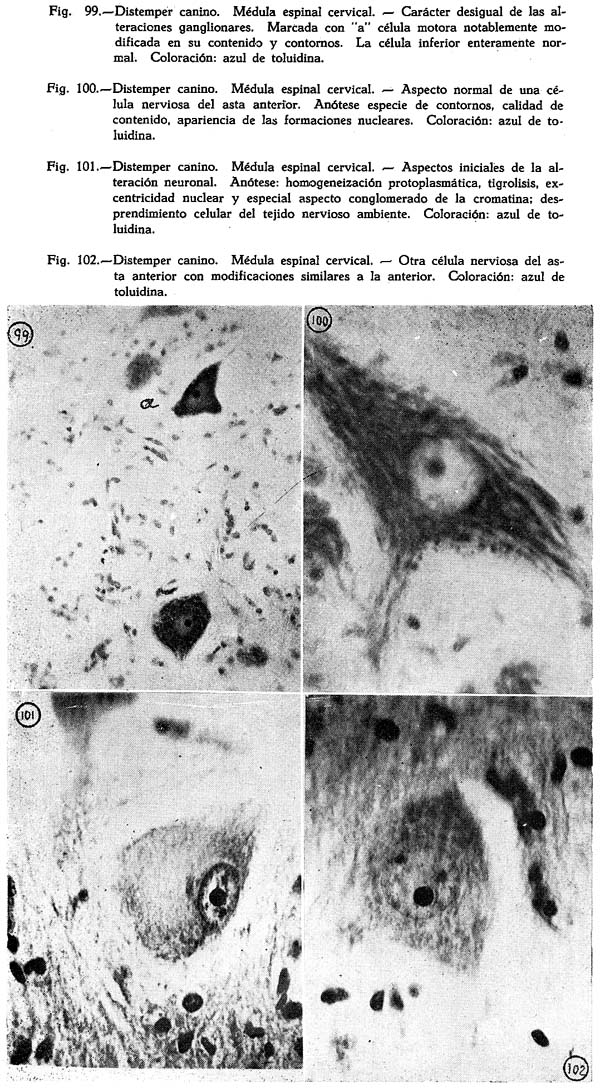
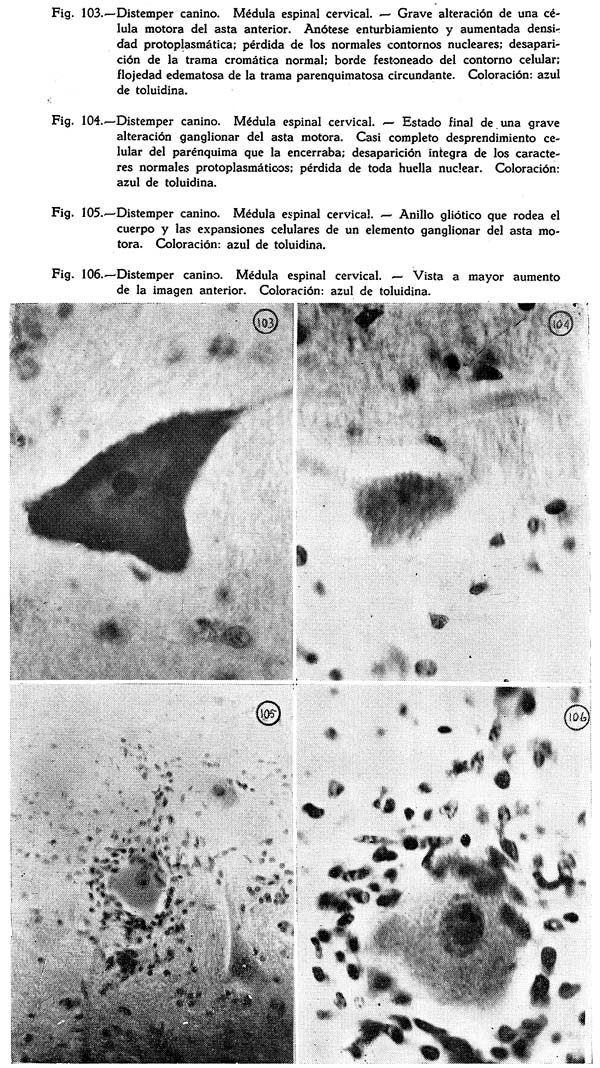
En lo referente a las alteraciones glióticas de la médula, aparte su acentuada proliferación y correspondientes concomitancias vasculares, se han presentado imágenes especiales, pocas veces comprobadas en la histopatología nerviosa. En las Figs. Nº 105 y 106, células motoras del asta anterior de la médula sino con grandes modificaciones -aunque las tienen evidentemente- se presentan rodeadas, no solo en su cuerpo sino en sus prolongaciones de elemento gliótico proliferado y alterado. Antes que papel alterativo neuronofágico parece, en este caso haberse organizado la glía en verdadera barrera reaccional defensiva, pues solo así se explica el hecho de los cambios de tan escasa significación en la célula nerviosa, pues se la ve conservando gran parte de sus caracteres morfológicos y estructurales normales. La reacción astrocitaria se manifiesta con la misma intensidad, tanto en la substancia gris como en la blanca, en la primera mas intensamente que en la segunda y en las astas afectadas con mayor intensidad, que en las menos alteradas. La Fig. Nº 107 nos muestra riqueza de tejido astrocítico difuso en un sector de la substancia gris, con relaciones evidentes vasculares bajo la forma de los conocidos pies chupadores de CAJAL y la Fig. Nº 108 un aspecto de la substancia blanca de la médula en la que, al lado de la expresión infiltrativa se advierte el exponente de una rica formación astrocitaria al lado derecho de la figura. Las formaciones de pies chupadores se notan con bastante claridad en las Figs. Nº 109 y 113; un estado de particular discreta fibrosis vascular y constitución de anillos glioconectivos perivasculares en la Fig. 110, de una asta enferma. La intensidad que adquieren las formaciones antedichas y la parte diferenciada que en la constitución de estos complejos vásculo-glióticos toman los vasos y los elementos glióticos se manifiestan ostensiblemente en la Fig. 113. Pero por sobre todo esto, lo interesante fué comprobar cómo la reacción gliótica ostensible en las porciones de la médula particularmente afectadas por el trastorno es pobre, nula, o normal, en la zona de la médula que no ha sido tocada. En efecto, si examinamos la Fig. Nº 114 correspondiente a un segmento de médula espinal histológicamente normal, comprobaremos que la rica trama está constituida predominantemente por las prolongaciones profusas de escasas células gliáticas. Si observamos en cambio las Figs. Nos 115 y 116 fácilmente se advierte la riqueza de producción astrocitaria fibrosa, lado a lado de neuronas, como se ve, manifiestamente enfermas. Notable es pues la evidencia de la reacción del componente gliótico en concordancia con porciones particularmente afectadas del órgano.
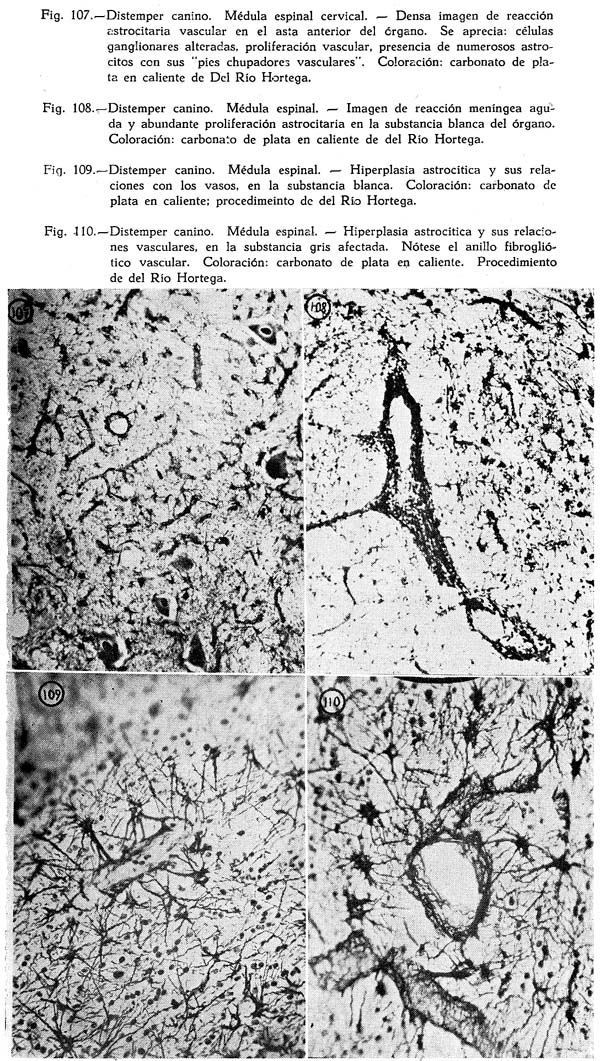
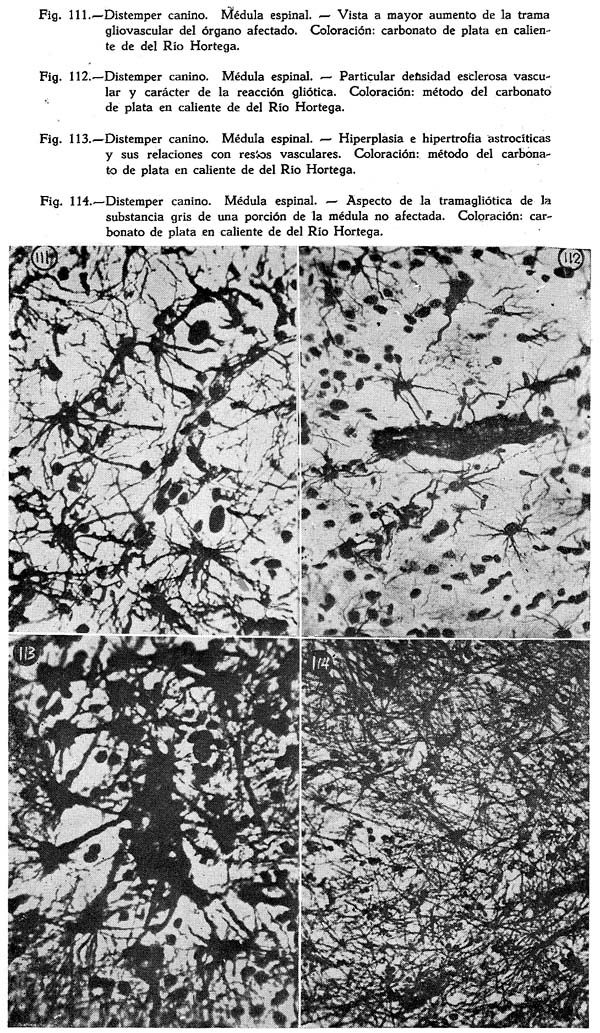
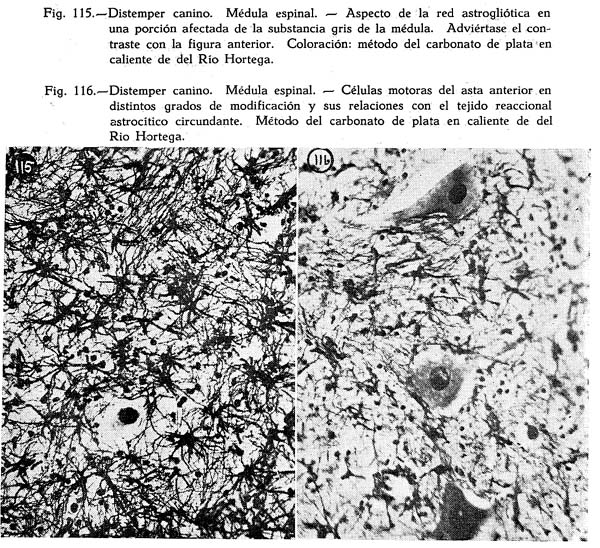
En lo referente a las alteraciones glióticas de la médula, aparte su acentuada proliferación y correspondientes concomitancias vasculares, se han presentado imágenes especiales, pocas veces comprobadas en la histopatología nerviosa. En las Figs. Nº 105 y 106, células motoras del asta anterior de la médula sino con grandes modificaciones aunque las tienen evidentemente se presentan rodeadas, no solo en su cuerpo sino en sus prolongaciones de elemento gliótico proliferado y alterado. Antes que papel alterativo neuronofágico parece, en este caso haberse organizado la glía en verdadera barrera reaccional defensiva, pues solo así se explica el hecho de los cambios de tan escasa significación en la célula nerviosa, pues se la ve conservando gran parte de sus caracteres morfológicos y estructurales normales. La reacción astrocitaria se manifiesta con la misma intensidad, tanto en la substancia gris como en la blanca, en la primera mas intensamente que en la segunda y en las astas afectadas con mayor intensidad, que en las menos alteradas. La Fig. Nº 107 nos muestra riqueza de tejido astrocítico difuso en un sector de la substancia gris, con relaciones evidentes vasculares bajo la forma de los conocidos pies chupadores de CAJAL y la Fig. Nº 108 un aspecto de la substancia blanca de la médula en la que, al lado de la expresión infiltrativa se advierte el exponente de una rica formación astrocitaria al lado derecho de la figura. Las formaciones de pies chupadores se notan con bastante claridad en las Figs. Nº 109 y 113; un estado de particular discreta fibrosis vascular y constitución de anillos glioconectivos perivasculares en la Fig. 110, de una asta enferma. La intensidad que adquieren las formaciones antedichas y la parte diferenciada que en la constitución de estos complejos vásculo-glióticos toman los vasos y los elementos glióticos se manifiestan ostensiblemente en la Fig. 113. Pero por sobre todo esto, lo interesante fué comprobar cómo la reacción gliótica ostensible en las porciones de la médula particularmente afectadas por el trastorno es pobre, nula, o normal, en la zona de la médula que no ha sido tocada. En efecto, si examinamos la Fig. Nº 114 correspondiente a un segmento de médula espinal histológicamente normal, comprobaremos que la rica trama está constituida predominantemente por las prolongaciones profusas de escasas células glióticas. Si observamos en cambio las Figs. Nº 115 y 116 fácilmente se advierte la riqueza de producción astrocitaria fibrosa, lado a lado de neuronas, como se ve, manifiestamente enfermas. Notable es pues la evidencia de la reacción del componente gliótico en concordancia con porciones particularmente afectadas del órgano.
ORGANOS EXTRACEREBRALES
Interesante es la formación de un gran nódulo de tejido intersticial hiperplásico que ha ahogado en gran proporción al parénquima secretor del órgano, y que en su porción central está necrosado. La Fig. Nº 117 muestra al nódulo antedicho y la Fig. Nº 118 a su porción modificada. Hay hiperplasia del conectivo intertubular, sobre todo del que rodea a los tubos contorneados; hiperplasia mas aparente aún por la intensidad con que se han coloreado sus células componentes. El epitelio de los tubos secretores muestran alteraciones degenerativas, granulosas, de su protoplasma, y en las vecindades una considerable hipertrofia de los núcleos de sus componentes conectivos: Fig. Nº 120. El pulmón presenta focos difusos o circunscritos de alveolitis catarrales exudativas: Fig. Nº 121, y el hígado, la presencia de bandas fibrosas pericentrolobulillares con alteración considerable de los cordones hepáticos, inmediatos: Fig. Nº 122.
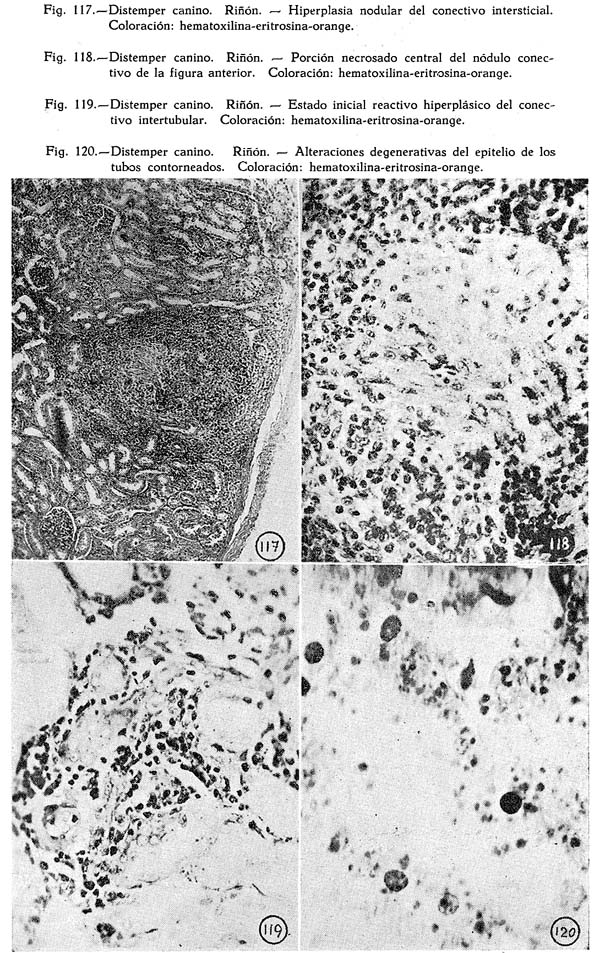
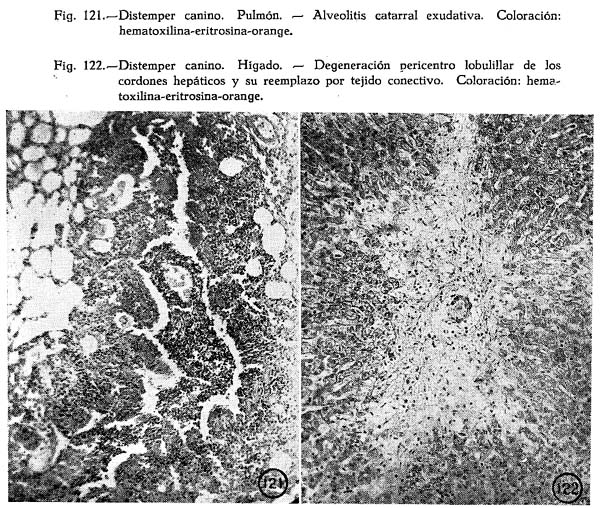
BIBLIOGRAFÍA
1. V. BABES & C. STARCOVICI: Compt. rend. Soc. de Biol., 1912, 73, 228. [ Links ]
2. H. CARRÉ: Compt. rend. Acad. d. sc., 1905, 140, 589. [ Links ]
3. H. CARRÉ: Compt. rend. Acad. d. sc., 1905, 140, 1491. [ Links ]
4. G. W. DUNKIN & P. P. LAIDLAW J. Comp. Path. & Therap., 1926. 39,201. [ Links ]
5. G. W. DUNKIN & P. P, LAIDLAW J. Comp. Path. & Therap., 1926, 39,213. [ Links ]
6. J. P. M'GOWAN J. Path. & Bact., 1910-11, 15, 372. [ Links ]
7. R. G. GREEN, N. R. ZIEGLER, B. B. GREEN & E. T. DEWEY : Am. J. Hyg., 1930, 12, 109. [ Links ]
8. R. G. GREEN, B. B. GREEN, W. E. CARLSON & T. E. SHILLINGER Am. J. Hyg., 1936, 24, 57. [ Links ]
9. R. G. GREEN & J. E. SHILLINGER : Am. J. Hyg., 1934, 19, 362. [ Links ]
10. J. G. HARDENBERGH : University of Illions, February, 16-17, 1926. [ Links ]
11. H. R. KREISEL : Departament. of Patology and Bacteriology, New York State, Veterinary Collage. [ Links ]
12. OTTO LENTZ : Ztschr. f. Hyg. u. Onfektionsk, 1908-09, 62. [ Links ]
13. A. LOCKHART & S. R. JOHNSON : Veterinary Medical Association. August, 25-28, 1931. [ Links ]
14. A. LOCKHART: Veterinary Medicine. March, 1934, 136. [ Links ]
15. A. LOCKHART: Veterinary Medical Association, August. 17-20, 1926. [ Links ]
16. G. MARINESCO, DRAGANESCO, STATE & G. STROCESCO : Ann. d. l´ Inst. Pasteur., 1933, 51, 215. [ Links ]
17. W. A. DE MONBREUN : Am. J. Path., 1937, 13, 187. [ Links ]
18. FERRY S. NEWELL : J. Infec. Dis., 1911, 8, 399. [ Links ]
19. S. NIVOLAU : Compt. rend. Soc. de Biol., 1935, 119, 269. [ Links ]
20. B. ROMÁN & CH. M. LAPP : Veterinary Medical Association, July, 1924. [ Links ]
21. A. S. SCHLINGMAN : Veterinary Medical Association. August, 25-28, 1931. [ Links ]
22. R. STANDFUSS : Arch. f. Wissensch u. prakt. Tierheilk., 1908, 34,109. [ Links ]
23. C. TORRES MAGARINOS & J. TEXEIRA DE CASTRO: Instituto Oswaldo Cruz, 1935, 30, 215. [ Links ]

