










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
Print version ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.4 no.4 Lima Dec. 1945
TRABAJOS ORIGINALES
Catalepsia experimental producida por la Cocaína.
C. Gutiérrez-Noriega1 & V. Zapata Ortiz1
1 Departamento de Farmacología del Instituto Nacional de Higiene.
RESUMEN
Se ha estudiado un caso de catalepsia producida por cocaína en un perro sometido a tratamiento crónico con esta droga. Las reacciones catalépticas se presentaron después de un mes de iniciado el tratamiento, y se hicieron progresivamente más intensas desde su comienzo en el curso de 14 meses de observación. Las conclusiones son las siguientes :
1. La intensidad y duración de las reacciones catalépticas eran proporcionales a la dosis de cocaína. Casi todas estas reacciones eran precedidas y seguidas por un estado de excitación motriz.
2. Con las dosis más altas disminuía el estado de excitación y se intensificaba la catalepsia.
3. La hipertermia cocaínica disminuía con la inmovilidad cataléptica, pero no llegaba a suprimirse.
4. La inyección de barbitúricos durante el estado cataléptico suprimía esta reacción, originando una intensa excitación motriz.
5. La inyección. de benzoato de cafeína practicada antes de la inyección de cocaína, intensificó notablemente la catalepsia producida por la última droga.
6. La inyección aislada de neuroestimulantes (cafeína, cardiazol y coramina) no produjo catalepsia.
7. Durante la catalepsia cocaínica se observó discreto aumento de la glucemia y descenso del fósforo inorgánico sanguíneo. No se demostró modificaciones significativas de la fosfatasa alcalina de la sangre, de la úrea, reserva alcalina y del calcio.
8. Se discute el mecanismo fisiológico de la catalepsia cocaínica, considerándose] e como una excitación de los centros nerviosos más intensa que la excitación que produce la agitación motriz, pero menos intensa que la excitación del mal epiléptico cocaínico.
La cocaína, clasificada en el grupo de estimulantes del sistema nervioso, tiene propiedades farmacodinámicas muy peculiares, que le confieren una posición muy especial en la farmacología del sistema nervioso. Es el único neuroestimulante que tiene acción depresiva sobre el centro respiratorio y que se comporta, a dosis estimulantes, como antagonista de las drogas convulsivantes. 1, También se caracteriza por la intensidad y gran duración de su acción excitante sobre los centros motores, y hasta se puede afirmar que es el estimulante por excelencia del sistema neuromotor.
En algunos casos, la acción neuroestimulante se exterioriza por un fenómeno aparentemente depresivo, la catalepsia. Esta reacción nerviosa ha sido considerada como un estado de inhibición, pues, en efecto, consiste en la parálisis de la actividad cortical -en el hombre por inhibición total o parcial de la conciencia- con inhibición simultánea, de la actividad motriz espontánea, y con fenómenos de flexibilidad cérea.
En el perro es verdaderamente excepcional que la cocaína origine catalepsia (o el síndrome de la catatonía experimental, según la denominación propuesta por DE JONG & BARUE).2 Empero, en el gato la reacción ha sido descrita, demostrándose que la cocaína tiene sobre el sistema nervioso central una acción catatonígena como la bulbocapnina. 3 El empleo de la cocaína en el tratamiento del estupor catatónico, preconizado por ciertos autores (BERGER, FLECK, BYCHOWSKI), y sus favorables resultados son aparentemente paradójicos. Empero. estos resultados no son muy demostrativos según las observaciones de J. M. SACRISTÁN (Archivos de Neurobiología, v. 5, p. 166, 1925).
El caso de catalepsia experimental cocaínica que es tratado en el presente trabajo nos ha parecido interesante y digno de estudio, no solo por ser uno de los pocos ejemplos de la acción cataleptígena de la cocaína, sino porque revela muchos detalles M mecanismo de acción de la droga y de la fisiopatología de la catalepsia.
Se trata de un perro que presentó típicas y prolongadas catalepsias en el curso de un tratamiento crónico con clorhidrato de cocaína, al que fué sometido para estudiar los fenómenos de habituación experimental a esta droga.
El tratamiento se inició en diciembre de 1944 con dosis interdiarias de 5 mgrs. x Kgr. de clorhidrato de cocaína en solución al 1 %, inyectada en la vena yugular.
Durante los primeros días sólo se observaron las reacciones que suelen presentar todos los perros que reciben cocaína : gran excitación motora, movimientos de circo, euforia e hipertermia de 40 u 80´ de duración, acompañadas de disnea, midriasis y de algunas reacciones neurovegetativas. Además, fenómenos de habituación a la droga, que en este animal fueron muy ostensibles.
Al llegar al segundo mes de tratamiento, se presentó un cambio notable : inmediatamente después de la inyección se manifestaba la habitual hipercinesia, la que luego era reemplazada por un estado de inmovilidad, con la hipertonía generalizada, predominante en la nuca, con flexibilidad cérea ( fig. 1 ). Tales reacciones eran casi idénticas a las que origina la bulbocapnina, y que DE JONG & BARUK llamaron catatonía experimental, pero que nosotros, por no implicar relaciones con el síndrome clínico del mismo nombre,, denominamos catalepsia experimental.

En primer lugar, se observó un típico estado de flexibilidad cérea; el animal tomaba todas las posturas bizarras y afisiológicas que se le imponían, como lo demuestran las figuras 1 y 2. Tal fenómeno se iniciaba algunos minutos después de la inyección, y duraba 30 a 60 minutos. Al desaparecer la flexibilidad cérea y las otras manifestaciones de catalepsia, se reiniciaba el estado de excitación. La flexibilidad cérea producida por la cocaína no difiere cualitativamente de la producida por la bulbocapnina, siendo en el caso que estudiamos relativamente más intensa y prolongada que esta.

En segundo lugar, se demostró una completa pérdida de los movimientos espontáneos, la que se producía rápidamente, como si el animal experimentara una brusca parálisis en el curso de su excitación inicial. Esta pérdida de la iniciativa motriz siempre se acompañaba de alteraciones posturales : por lo general el perro caía sobre el dorso, con el cuello y los miembros en extensión. Si se le forzaba a estar de pie acababa por adoptar la postura que puede observarse en las figuras 1 y 2, es decir, con el cuello en extensión forzada, los miembros posteriores flexionados y los anteriores en extensión. Si se le imponía posturas bizarras e incómodas, las conservaba indefinidamente.
También fueron evidentes alteraciones del tono muscular. Los autores que anteriormente se han ocupado de la catatonía experimental o de la catalepsia experimental, no han concedido a esta alteración la importancia que merece, pues se trata de uno de los más típicos y significativos fenómenos del síndrome cataléptico. Las alteraciones del tono son en particular notables al iniciarse el estado cataléptico; la hipertonía predomina en el cuello, que como ya indicamos, se mantiene en un estado de hiperextensión, siendo necesario realizar un gran esfuerzo para flexionar la cabeza. También se observó una moderada rigidez de los músculos de las extremidades, en especial de los miembros anteriores. En conjunto, la hipertonía de la catalepsia cocaínica simula la hipertonía de la rigidez de descerebración, y hasta fué posible verificar la existencia de los reflejos tónicos del cuello. En efecto, al flexionarlo se obtenía rigidez en extensión de los miembros posteriores y flexión de los anteriores, mientras que se producía él efecto opuesto, flexión de los miembros posteriores y extensión de los anteriores, con la extensión del cuello. Es de tener en cuenta que en los catalépticos nunca se presentan los reflejos de MAGNUS y de KLEYN.4
Durante el estado cataléptico el perro era casi insensible a los estímulos dolorosos. Los autores que se han ocupado de la catalepsia experimental no han llamado la atención sobre la analgesia cataléptica, que ha sido descrita por primera vez por uno de nosotros.5 Al iniciarse el estado cataléptico se inhiben las reacciones a los estímulos dolorígenos (compresión mecánica de los miembros, corriente farádica), pero a medida que menguan la catalepsia y la flexibilidad cerca, la sensibilidad se recupera progresivamente. Una interesante observación puso en evidencia el fenómeno de la analgesia cataléptica que describimos : nuestro perro sufrió incidentalmente graves heridas, que era indispensable suturar. En vez de recurrir a la anestesia general, se le inyectó 5 mgrs. x Kgr. de cocaína por vía endovenosa, y durante la catalepsia consecutiva a la inyección se pudo realizar la operación quirúrgica sin que el perro manifestara ni el menor indicio de dolor. El hecho indica que la pérdida de sensibilidad al dolor era completa.
Finalmente, se registraron alteraciones neurovegetativas, en especial sialorrea y midirasis.
La reacción cataléptica fué constante durante todo el largo período de la intoxicación crónica, siempre precedida por un estado de intensa excitación motora, y seguida por una excitación motora de menor intensidad. Por lo regular, con la dosis de 5 mgrs. x Kgr. de peso de clorhidrato de cocaína, la excitación inicial duraba de 5 a 10 minutos, la catalepsia 30 ó 60 minutos, y la excitación postcatalépticá 10 ó 20 minutos. Esta reacción fué observada con absoluta regularidad durante seis meses. En los intervalos de las inyecciones, no se registró ninguna anomalía en la conducta del perro.
Relación entre la dosis de cocaína y la catalepsia. Con el objeto de averiguar si existía una relación entre la dosis administrada y la reacción cataléptica, sometimos al animal a un tratamiento con dosis progresivas, iniciándolo con 0.5 mgrs. x Kgr. Los resultados se han sintetizado en la fig. 3, en la que puede apreciarse el progresivo aumento de la reacción proporcional al aumento de dosis.
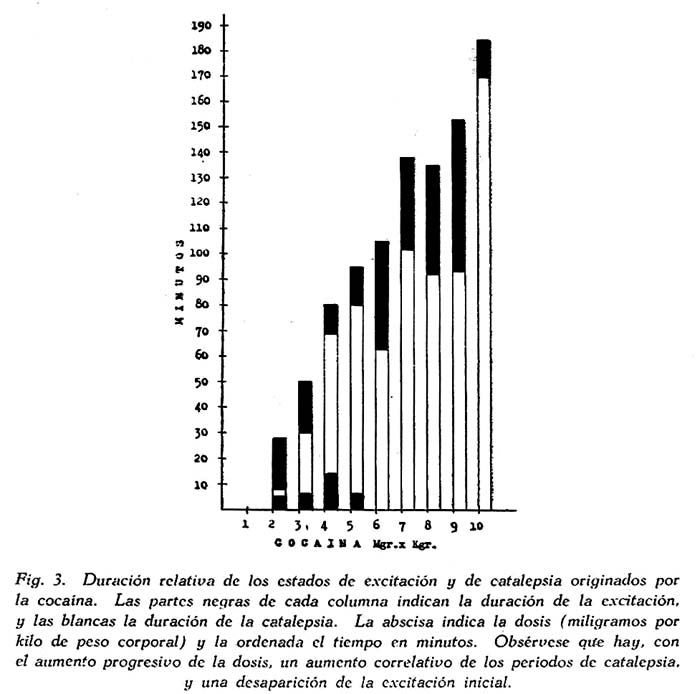
En primer lugar, es digno de tenerse en cuenta que la pequeña dosis de 2 mgrs. x Kgr. determinara la reacción, aunque breve y de poca intensidad. Es de advertir que esta dosis sólo produce un estado de leve excitación en los perros normales. Empero, al repetir esta prueba con la misma dosis, se obtuvo una intensa reacción cataléptica que duró 98 minutos, precedida y seguida, como siempre, por estado de excitación. A medida que se aumentó la dosis la reacción cataléptica era más prolongada, desapareciendo la excitación motora precataléptica. Con dosis muy alta, y cercana ya a la dosis convulsiva umbral (10 mgrs. x Kgr.) la excitación se reduce al mínimo, y la catalepsia llega a 170 minutos de duración.
Puesto que el grado de excitación del sistema nervioso producido por la cocaína aumenta con la dosis, de las observaciones experimentales que hemos referido se deduce que la catalepsia cocainica es un estado de excitación, y no de inhibición, como podría inferirse por sus manifestaciones sintomáticas.
También el hecho de que la catalepsia sea precedida y seguida por estados de excitación sugiere que ella también es un estado de excitación. Se trata, aparentemente de una excitación progresivamente más intensa a medida que la cocaína se fija en el sistema nervioso a partir de la inyección, y que se transforma en catalepsia cuando la excitación llega a su clímax; luego, cuando la excitación disminuye debido a la eliminación de la droga, la catalepsia desaparece y se reinicia el estado de excitación motora. La figura 3 revela muy objetivamente este punto de vista.
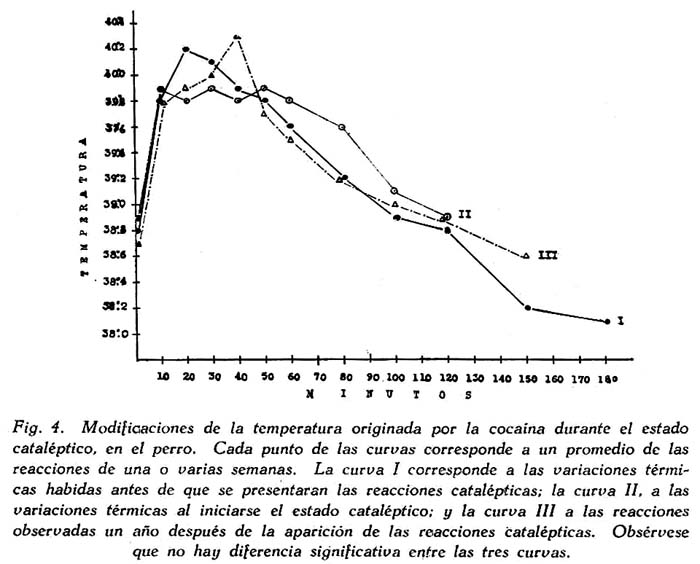
Modificaciones térmicas. Durante la excitación, cocaínica se produce un aumento de temperatura, que en algunos casos es una verdadera reacción febril, y que se ha atribuido al estado de agitación motriz. En tal caso, la reacción cataléptica que implica una completa inmovilidad, debería suprimir la fiebre cocaínica. Esto no se observa; la catalepsia disminuye simplemente la reacción febril, pero no la suprime, como puede observarse en la fig. 4, Es muy probable que la hipertonía que hemos mencionado sea uno de los factores del aumento de temperatura durante la catalepsia cocaínica.
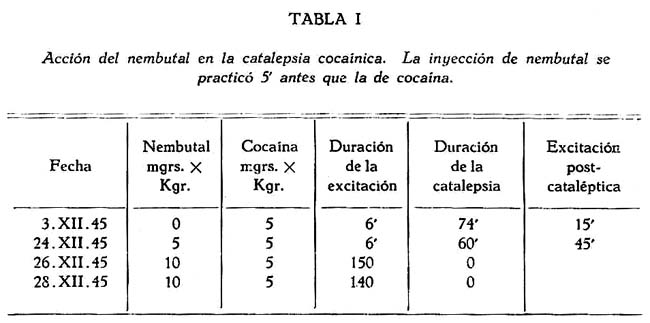
Catalepsia cocaínica y barbitúricos. Con el objeto de averiguar si la catalepsia cocaínica es un estado de inhibición del sistema nervioso, o un estado de excitación, como parece demostrarlo el experimento ya referido de las dosis progresivas, sometimos al perro en estudio a la acción del nembutal y del luminal. Si la catalepsia se debe a una depresión de los centros corticales era de esperar su refuerzo por acción de estos barbitúricos.
En los primeros experimentos el nembutal fué administrado por vía endovenosa, a dosis subnarcótica, antes de la cocaína. Se observó que con pequeñas dosis (5 mgr. x Kgr.) se presentaba una reducción de la catalepsia, y prolongación del estado de excitación; y que con dosis mayores ( 10 mgr. x Kgr.) la catalepsia desaparecía por completo, reduciéndose la reacción a la cocaína a un estado de excitación (tabla I). Este inesperado y paradójico resultado hace aún más probable la hipótesis de que la catalepsia sea una excitación del sistema nervioso de mayor intensidad que la excitación que da lugar a las reacciones hipercinéticas.
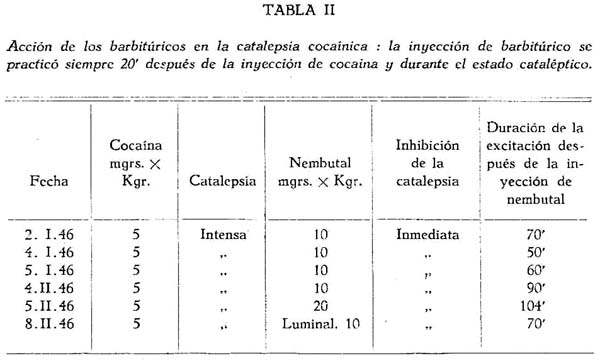
En una segunda serie de experimentos, en los que ya se inyectó el nembutal por vía endovenosa durante la catalepsia, el resultado no pudo ser más espectacular e interesante. El barbitúrico actuaba aparentemente como un verdadero estimulante, pues inmediatamente después de inyectado se desencadenaba una intensísima excitación motora, qué solía durar una hora, u hora y medía. Si sólo se inyectaba 10 mgr. x Kgr. de nembutal (el tercio de la dosis narcótica del perro) se suprimía al instante la catalepsia y la excitación cocaínica se revelaba como en un perro normal que ha recibido una dosis de 5 mgr. x Kgr. Este experimento se repitió tres veces con el mismo resultado. Si se inyectaba una fuerte dosis de nembutal (20 mgrs. x Kgr.) el perro se manifestaba atáxico y muy excitado durante los primeros minutos; luego desaparecía o disminuía la ataxia, y la excitación cocaínica continuaba hasta después de una hora con gran intensidad (tabla II).
La espectacular acción anticataléptica del nembutal, cuando se administra por vía endovenosa, se relaciona con la acción terapéutica de los barbitúricos en las neurosis de guerra,6 en especial en casos de estupor o estados de depresión similares, en que la inyección endovenosa de aquel barbitúrico a dosis subnarcótica exteriorizaba la situación correspondiente al trauma patógeno, es decir, al estado de excitación inicial precursor de la depresión subsiguiente. También en este caso surge la sospecha de que tales estados depresivos, o mas bien pseudodepresivos, enmascaran intensas excitaciones cerebrales.
Conviene recordar, para interpretar este sorprendente resultado de nuestros experimentos, las dos teorías fisiopatológicas de la catatonía producida por la bulbocapnina. La escuela francesa, con DE JONG & BARUK, afirma que aquella droga tiene una acción cortical difusa, y que, en consecuencia, la catalepsia se debe a una alteración general de la corteza. La escuela rumana, con SAGER principalmente, sostiene que la alteración es local, que la corteza cerebral no es indispensable para que se manifieste la catalepsia, y que esta depende principalmente de una intoxicación de los centros diencefálicos y mesencefálicos, con consiguiente liberación del romboencéfalo ( metencéfalo y mielencéfalo), lo que da lugar a un estado análogo al de la rigidez descerebrada. La catalepsia por inhibición cortical obtenida por PAVLOV concordaría con este punto de vista.
La acción anticataléptica y pseudoestimuladora del nembutal plantea, en relación a estas explicaciones, nuevos problemas. Sí la catalepsia es, como afirma SAGER, una intoxicación diencefalomesencefálica con liberación romboencefálica, es inexplicable que un barbitúrico, que deprime principalmente el diencéfalo, suprima el fenómeno en vez de intensificarlo. La contradicción es más notoria si se tiene en consideración que los centros de la marcha se encuentran situados en el perro en la región diencefalomesencefálica, es decir, en la zona sobre la que ejerce principalmente su acción depresiva el barbitúrico.
Por el momento nos concretamos a señalar que nuestros experimentos contradicen la teoría de los fisiólogos humanos, reservando para el final la discusión general del problema.
Catalepsía cocaínica y neuroestimulantes. Los fenómenos observados al inyectar neuroestimulantes en el perro con catalepsia cocaínica, no son menos interesantes que los que acabamos de referir.
Se demostró un notable sinergismo entre la cafeína y la cocaína (tabla III). La primera droga, aisladamente y a la dosis de 30 mgrs. X Kgr., sólo producía un moderado estado de excitación, pero si en estas condiciones se inyectaban pequeñas dosis de cocaína, se producía una intensa catalepsia. Así la dosis de 2 mgrs. x Kgr. de cocaína, que por lo regular sólo producía una ligera catalepsia, daba lugar a intensas reacciones de este tipo en el perro con cafeína. Aun con la dosis de 1,5 mgrs x Kgr. de cocaína se obtuvo una catalepsia de 43 minutos de duración y hasta con la ínfima dosis de 1 mgr. x Kgr. fué posible obtener una breve catalepsia cuando se estimulaba al perro previamente con cafeína. También la inyección de cafeína en el curso de la excitación cocaínica produjo un gran incremento de la catalepsia ya existente, en un caso con la notable duración de dos horas veinte minutos.
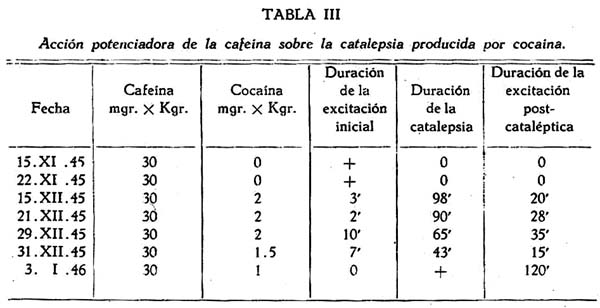
Es singular que sólo la cafeína, entre los estimulantes ensayados, actuara como potenciador del efecto cataleptígeno de la cocaína. El cardiazol y la coramina, al contrario, lo disminuían o suprimían, a la dosis de 10 ó 50 mgrs. x Kgr. respectivamente.
La administración de cardiazol a dosis convulsivante (14 mgrs. x Kgr.) durante la catalepsia cocaínica, dió lugar a un acortamiento de esta reacción. No se pudo demostrar, como en el caso de la cafeína, sinergismo entre cocaína y cardiazol con respecto a la reacción cataléptica.
Tampoco se pudo obtener catalepsia por administración aislada de cafeína, cardiazol o coramina (a la dosis de 100 mgrs. x Kgr. en el último caso), lo que indicaría que tal reacción es específica con respecto a la cocaína, y no para otros neuroestimulantes.
Inyectándose bulbocapnina (15 mgrs. x Kgr. por vía endovenosa) se obtuvo una catalepsia que no difería de la que por lo regular se produce en los perros por aquella droga y a la dosis indicada, pero mucho menos durable y menos intensa que la catalepsia cocaínica en el perro estudiado.
No se observó ninguna alteración significativa de la dosis umbral convulsivante de cardiazol, ni en el curso de la catalepsia cocaínica ni fuera de ella.
Modificaciones químicas de la sangre. Se investigó la glucemia, el fósforo, la fosfatasa, el calcio, la úrea y la reserva alcalina (tabla IV)
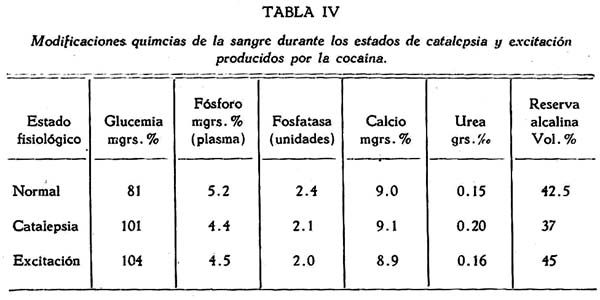
Por lo menos se tomaron tres muestras de sangre; antes de la inyección de cocaína, durante el estado cataléptico y durante el estado de excitación post-cataléptica.*
En dos observaciones se comprobó un discreto aumento de la glucemia, lo que en realidad no es significativo en relación a la catalepsia, pues está demostrado que la cocaína produce este efecto; mas es de destacar en este caso que la hiperglucemia se presentó a pesar de la inmovilidad cataléptica, no siendo dable atribuirla, como piensan ciertos autores, a la excitación motora originada por la cocaína.
Más interesantes son los datos obtenidos del análisis del fósforo inorgánico en la sangre. Durante el estado cataléptico se observó una discreta disminución del fósforo. No se obtuvieron cambios significativos de la fosfatasa.
Las modificaciones del calcio, de la reserva alcalina y de la úrea, en, realidad fueron poco importantes y no las consideramos como significativas.
En resumen, de las modificaciones químicas de la sangre durante la caatlepsia cocaínica sólo tienen alguna importancia las del fósforo y la glucemia, pero estas modificaciones no son privativas de la catalepsia, pues son peculiares de la acción general de la cocaína.
Examen anatomo-patológico. Después de la muerte del perro, que tuvo lugar incidentemente a consecuencia de un colapso cardio-circulatorío producido por una inyección de cafeína, se practicó la autopsia y el examen anatomo-patológico que exponemos a continuación,** con excepción del cerebro que aún está en estudio.
Se demostró la existencia de lesiones hemorrágicas en el páncreas y en la suprarrenal. En los focos hemorrágicos de la suprarrenal se encontró hemosiderina, lo que permite suponer que eran antiguos y sin relación con la causa de la muerte. Los focos hemorrágicos del páncreas, al contrario se produjeron probablemente antes de la muerte.
DISCUSIÓN
El caso de catalepsia producida por cocaína que hemos estudiado es muy interesante y justifica el estudio especial que le hemos dedicado por muchas razones. En primer lugar, porque aparece en el curso de una intoxicación crónica, y, aparentemente, como expresión de los fenómenos de sensibilización a esta droga que suelen manifestarse durante su administración prolongada.
En segundo lugar por ser el caso único en la literatura farmacológica de un estimulante que produce intensas reacciones catalépticas, aun a dosis muy pequeñas. En efecto, algunos neuroestimulantes producen reacciones catalépticas solo a dosis muy altas, casi siempre convulsivantes, manifestándose la catalepsia como secuela postconvulsiva. El ejemplo que hemos referido se caracteriza, porque las dosis cataleptígenas fueron 4 ó 5 veces menores que las dosis convulsivantes.
En tercer lugar, la constancia de la reacción y la forma como fué estudiada permitieron observaciones del mayor interés para comprender el mecanismo neuro fisiológico de la catalepsia.
Tanto las observaciones sobre la relación de la dosis con los estados de excitación motriz y de catalepsia, como el hallazgo de la espectacular acción anticataléptica y pseudoestimulante de los barbitúricos, indican que la catalepsia es un estado de excitación y no un estado de inhibición del sistema nervioso. Nuestras observaciones están, en consecuencia, en formal oposición a las concepciones de SAGER 7 y de VAN HARREVELD & KOK,8 según los cuales la catalepsia es una parálisis del sistema nervioso. Empero, la posibilidad de que la excitación se combine con la depresión de ciertos centros no debe descartarse, pues se ha demostrado que durante la catalepsia producida por el óxido de deuterium, disminuye considerablemente la actividad cortical bíoeléctrica.9
No nos inclinamos a formular, como por lo general se acostumbra en estos casos, una explicación de carácter anatómico, a base de un antagonismo de ciertos centros con respecto a otros; y nos parece mucho más conveniente, para aproximarnos a la comprensión de estos complicados fenómenos, tener en cuenta las diferencias funcionales entre los estados de excitación motriz y de catalepsia. En el primer caso, predominan las reacciones fásicas, es decir, la sucesión rítmica de extensiones y flexiones; en el segundo, las reacciones tónicas. La reacción fásica se caracteriza por salvas intermitentes e irregulares de impulsos nerviosos, mientras que en la reacción tónica la emisión de impulsos es continua y regular, de suerte que la reacción muscular a que da origen es una invariable contracción tetánica o semitetánica.
Hay otros ejemplos en que un proceso de excitación creciente del sistema nervioso se traduce primero por reacciones fásicas y en segundo lugar por reacciones tónicas. Así, la estricnina, a dosis progresivas, da lugar a un estado de hipercinesia, y cuando la excitación aumenta, a una rigidez generalizada. Lo mismo ocurre con la cocaína, que a dosis muy altas, y casi cercanas al umbral convulsivante, puede originar estados de rigidez general muy intensa. En la catalepsia por bulbocapnina hay con frecuencia un estado de hipercinesia precataléptico y otro postcataléptíco, y también en este caso la catalepsia se acompaña de ostensibles hipertonías de los miembros y del cuello. Aun más típico es el efecto de la hípoglicemia insulínica, según las observaciones que se han realizado en el tratamiento de SAKEL,10 en el que la excitación nerviosa se inicia casi siempre con reacciones fásicas, presentándose como culminación del proceso de excitación un estado de hipertonía generalizada análogo a la rigidez de descerebración. La administración de glucosa, al disminuir la intensidad de la excitación central, permite en ciertos casos una marcha regresiva, en la que la hipercinesia reaparece al suprimirse la rigidez. En este caso, como la catalepsia que hemos estudiado, los fenómenos de hipertonía son precedidos y seguidos por un estado de agitación motriz.
Nos preguntamos si en todos estos casos los fenómenos de hipertonía son reacciones de protección del organismo, y si en realidad representan una reacción económica. Pero no sería una reacción localizada en el romboencéfalo e idéntica a la rigidez de descerebración, con la que sólo se relaciona por simple analogía, sino de una reacción nerviosa general del sistema funcional tónico, en la que incluso participan los centros subcorticales y la misma corteza, que se opone a las reacciones del sistema funcional fásico. El hecho de que los barbitúricos supriman esta primera reacción sin afectar la última, está en favor de nuestra interpretación funcional mas que de cualquiera otra explicación localizacionista. En otros términos, la acción depresiva del barbitúrico no se ejerce sobre un determinado centro inhibiendo su reacción, sino difusamente sobre el sistema nervioso, paralizando un estado funcional y liberando el tipo de reacción inhibido por aquél.
AGRADECIMIENTOS
Expresamos nuestro agradecimiento al Sr. GERMÁN PAREDES por su colaboración técnica en este trabajo.
NOTAS DE PIE
1. C. Gutiérrez-Noriega & V. Zapata Ortiz : Revista de Medicina Experimental, v. 4, p 59. 1945.
2. H. De Jong & H. Baruk: La catatonia experimentale par la bulbocapnine, París, 1930.
3. C. Gutiérrez-Noriega & H. Rotondo : Revista de Neuro-Psiquiatria, v. 2. p. 73, 1939.
4. H. Baruk Psychiatrie médicale, Psysiologique et etperimentale, Paris, 1938.
5. C. Gutiérrez-Noriega : Act. Med. Per., v. 4, p. 29, 1938; y Actas de la Segunda Reunión de las jornadas Neuro - Psiquiátricas Panamericanas, v. 2, p. 289, 1939.
6. J. Stephen Horsley : Narco-analysis, Londres, 1943.
7. O. Sager : Zeitschrift f. d. ges Experimentelle Medizin, v. 81, 1932.
8. Van Harrevled & D. J. Kok: Arch. Néerlandaises de Physiologie de I'Homme et des Animaux, v. 20, p. 411. 1935.
9. J. H. Hermann : Journal of Pharmacology and Experimental Therapeutics, v. 66, p. 265, 1939.
10. H. Delgado, F Valega & C. Gutiérrez-Norieqa : Revista de Neuro-Psiquiatria, v, 1, p. 463, 1938.
*Los análisis químicos fueron practicados por el Dr. Julio Pons, de la Sección de Bioquímica del Departamento de Farmacología.
**Este estudio fué efectuado por el Dr. Alberto Cuba del Departamento de Anatomía Patológica de la Facultad de Medicina.

