










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.11 n.1-2 Lima dic. 1957
Estudio Morfológico de los huevos de algunos Triatominos hallados en el Perú (Reduviidae, Hemiptera)*
Yolanda Lizaraso1
1 División de Estudios Epidemiológicos e Investigaciones Especiales del Instituto Nacional de Salud Pública, Lima, Perú.
* Resumen de la tesis presentada a la Facultad de Ciencias de la Universidad Nacional Mayor de San Marcos para optar el grado de Bachiller en Ciencias Biológicas, 1957.
SUMARIO
Se ha efectuado un estudio morfológico de los huevos de los siquientes triatominos encontrados hasta el presente en el Perú: Triatoma infestans (KLUG, 1834); T. dimidiata (LATREILLE, 1811); T. carrioni LARROUSSE 1926; Panstrongylus chinai (DEL PONTE, 1929); P. herreri WYGODZINSKY, 1948 y Belminus peruvianus HERRER et al., 1954. Para ello se consideran los caracteres macroscópicos y la ornamentación microscópica del corion. El material procede de ejemplares criados en el laboratorio, en todas las especies; en tres de éstas se ha utilizado también muestras procedentes de ejemplares colectados en la naturaleza. Las observaciones efectuadas nos permiten concluir lo siguiente:
1 . Los huevos de las especies procedentes de crías de laboratorio son diferentes entre sí en cuanto a sus dimensiones, demostrado estadísticamente; lo son igualmente en la ornamentación microscópica del corion, siempre que en las observaciones se emplee una misma técnica.
2. Las observaciones comparativas de huevos procedentes de ejemplares criados en el laboratorio y colectados en la naturaleza , (T. infestans, T. dimidiata y P. herreri), indican que existen diferencias estadísticamente significantes en las dimensiones; no así en los caracteres microscópicos del corion, los que permanecen constantes.
3. Por consiguiente, el carácter fundamental cuando se trate de establecer diferencias específicas debe ser la ornamentación microscópica del corion, mientras que las dimensiones constituirían caracteres secundarios o complementarios.
4. Se ha elaborado una clave para la identificación de los huevos de los triatominos estudiados, en la que se considera fundamentalmente la ornamentación microscópica del corion.
SUMMARY
A morphological study was conducted on the following species of peruvian triatomids: Triatoma infestans (KLUG 1834); T. dimidiata (LATREILLE, 1811); T. carrioni, LARROUSSE, 1926; Panstrongy1us chinai, (DEL PONTE, 1929); P. herreri, WYGODZINSKY, 1948 and Belminus peruvianus, HERRER, et al., 1954. Both the macroscopic morphology as well as the microscopic ornamentation of the triatomid eggs were considered in this study. In all species, eggs were collected from the insects reared in the laboratory. In three instances it was also possible to obtain eggs which had been layed in a natural environment. The conclusions of the study were as follows.
1 . While the eggs of each species of triatomid studied appeared to be consistent in dimensions within species, there was a statistically significant difference in the dimensions of eggs between species. The same relationships held also with respect to microscopic ornamentation of the egg shells.
2. Statistically significant differences were also observed within certain species between the gross dimensions of eggs collected in nature as compared to those obtained from insects reared in the laboratory. On the contrary, the microscopic ornamentation of the shells of both groups remained similar.
3. The need for employing the same technique throughout the preparation of all samples in such a correlation study is emphasized.
4. It is concluded that the most important differentiating characteristic in the species identification of triatomid eggs is microscopic ornamentation, while gross dimensions are of only secondary significance.
5. A classification chart or "identification key is presented based mainly on the microscopical ornamentation of the egg.
INTRODUCCIÓN
Es bastante conocida la importancia sistemática que se confiere a los caracteres morfológicos de los huevos de los insectos, En el caso particular de los triatominos, vectores de la trypanosomiasis americana existen cuando menos los trabajos de GALLIARD (1935), USINGER (1944) y ABALOS y WYGODZINSKY ( 1951 ), en los que considerando indistintamente los caracteres macroscópicos así como los detalles microscópicos del corion, se establecen diferencias entre algunas especies. Se sabe también que, en la búsqueda de tales insectos. es frecuente encontrar únicamente los huevos o las envolturas, lo que dificulta o imposibilita la identificación de aquellos.
En el Perú, especialmente en el transcurso de los últimos años y a consecuencia del mayor interés que se concede al conocimiento de la enfermedad de Chagas, las especies de triatominos hallados aumentan cada vez más. Estas circunstancias nos han inducido a realizar un estudio morfológico de los huevos de algunos triatominos encontrados hasta el presente en este país.
Para llevar a cabo este trabajo se ha contado con seis especies de triatominos que se mantienen en cría en el Instituto Nacional de Salud Pública, y además con ejemplares de algunas de estas especies colectadas recientemente en la naturaleza. Como resultado de tales estudios se dan a conocer tanto los caracteres morfológicos macroscópicos así como la ornamentación microscópica del corion o cáscara del huevo.
MATERIAL Y MÉTODOS
Los huevos estudiados pertenecen a las siguientes especies de triatominos: Triatona intestans (KLUG, 1834); Triatoma dimidiata (LATREILLE, 1811); Triatoma carrioni LARROUSSE, 1926; Panstrongylus chinai (DEL PONTE, 1929); Panstrongylus herreri WYGODZINSKY, 1948, y Belminus peruvianus HERRER et al., 1954. No obstante que Panstrongylus geniculatus (LATREILLE, 1811); Panstrongylus rufotuberculatus (CHAMPION, 1898); Rhodnius pictipes STAL, 1872 y Rhodnius robustus LARROUSSE, 1927, han sido encontrados también en el Perú, no fué posible incluirlos en estos estudios por falta de especímenes vivos que proporcionaran los huevos.
1. Procedencia de los triatominos.
a) Criados en el laboratorio. Dentro de este grupo en encuentran todas las especies que hemos estudiado. La procedencia de los insectos con los que se establecieron las crías así como el tiempo transcurrido hasta que se utilizaron, son los siguientes: T. infestans, con ejemplares de Arequipa, hace cuatro años; T. dimidiata, de Tumbes, hace un año y seis meses; T. carrioni, de Montero, departamento de Piura, hace un año; P. chinai, con especímenes enviados por el Dr. Francisco Campos del Instituto Nacional de Salud Pública "Leopoldo Izquieta Pérez" del Ecuador, hace seis meses; P. herreri, de Moyobamba, departamento de San Martín, hace un año; y B. peruvianus, de Yamón, departamento de Amazonas, hace seis meses. En el laboratorio se han mantenido a una temperatura fluctuante entre los 15 y 28º C. y a una humedad relativa de 70-95%. Estas especies se alimentaron regularmente en gallinas, salvo T. carrioni que en ciertas oportunidades lo hizo también en conejos debido a las dificultades que ofrece mantenerlos alimentándolos sólo en aves.
b) Colectados en la naturaleza. Se considera en este grupo a los especímenes adultos capturados recientemente en su ambiente natural. Las únicas especies que obtuvimos fueron: T. infestans, procedente de Arequipa; T. dimidiata, de Tumbes y P. herreri, de Moyobamba, departamento de San Martín.
2. Obtención de huevos.
De las especies criadas en el laboratorio, se emplearon 10 hembras y 5 machos en cada caso, tomadas al azar, con excepción de B. peruvianus en la que sólo se contó con 6 ejemplares (4 hembras y 2 machos). Los grupos así formados se mantuvieron en cajas adecuadas a 28º C. y 70 - 75% de humedad relativa, alimentándolos periódicamente en aves. En esta forma fué posible obtener un gran número de huevos de cada especie.
De los ejemplares colectados en la naturaleza, el número de hembras empleadas fué alrededor de 10 por especie. Los huevos examinados corresponden, exclusivamente, a los puestos durante el trayecto hacia el laboratorio.
3. Mediciones y cálculos estadísticos.
Se utilizaron 100 huevos recién puestos y cogidos al azar para obtener los datos biométricos, realizándose las mediciones mediante un ocular micrométrico. Las dimensiones consideradas son las siguientes: largo (eje mayor), ancho (eje menor) y diámetro opercular.
Por sugerencia del señor Raúl Vargas, estadígrafo del Servicio Cooperativo Interamericano de Salud Pública, se utilizó el Análisis de Varianza para comparar estadísticamente los huevos de las especies procedentes de crías de laboratorio; y, el Método del significado estadístico de la diferencia entre dos promedios para comparar las poblaciones de laboratorio y la naturaleza de una misma especie. La probabilidad de los valores hallados se calculó mediante las tablas respectivas.
4. Observaciones microscópicas del corion.
Se considera sólo la ornamentación microscópica del corion. El sistema micropilar, por tratarse de una estructura compleja, creemos que requiere de un estudio aparte.
Para las observaciones de esta índole se empleó fundamentalmente la técnica de Doetschman. Además de ésta se utilizó: a) un aclarante a base de ácido fénico, (6 partes) y alcohol etílico (4 partes); y b) glicerina pura para preparaciones momentáneas. En el caso particular de P. herreri se usó además nitrato de plata amoniacal, desde que con los procedimiento anteriores no se lograba visualizar detalles de ornamentación. Los cortes del corion se hicieron con una cuchilla fina a lo largo del eje mayor del huevo. Los dibujos se han efectuado mediante una cámara lúcida.
Breve reseña acerca de los caracteres morfológicos de los huevos de los triatominos
Son de forma más o menos oval, simétricos o asimétricos. De acuerdo al eje mayor del huevo se considera un extremo anterior y un extremo posterior. En el extremo anterior la cáscara presenta, según las especies, una constricción leve o profunda denominada cuello, que se expande para formar el reborde de la abertura opercular destinada a recibir el opérculo, formación discoidal de convexidad variable. La coloración de los huevos cambia desde el blanco o rosado en el momento de la puesta, hasta el castaño, rojo o grisáceo cuando culmina el desarrollo embrionario. El color se debe exclusivamente al contenido del huevo, ya que la cáscara es en sí blanca o traslúcida y de aspecto opaco o brillante, lo que permite visualizar estructuras embionarias tales como los puntos oculares.
Se acepta que los huevos en el momento que son puestos poseen dos envolturas: la membrana vitelina y el corion o cáscara. Existe además, en mayor o menor proporción, una cubierta superficial de cemento.
De acuerdo a BEAMENT (1946), quien ha estudiado detalladamente estos aspectos en Rhodnius prolixus STAL, 1859, la membrana vitelina se forma por la condensación de la parte más externa del oocito maduro, mientras que el corión es la envoltura segregada por el folículo ovárico.
En el corion se distingue fundamentalmente dos capas: el endocorion y el exocorion. El endocorion se forma sobre la membrana vitelina en la primera fase de actividad secretora del folículo ovárico, es la parte más flexible de la cáscara y varía en espesor en las diferentes regiones del huevo; es también la que con mayor facilidad toma los colorantes. El exocorion se forma inmediatamente sobre el endocorion en una segunda fase de actividad secretora, es la capa que da rigidez a la cáscara y, fundamentalmente, la que presenta en su superficie externa la ornamentación poligonal específica, huella dejada por el epitelio folicular que lo formó. En la región del cuello, esta ornamentación varía un tanto con respecto al resto siendo unas veces más complicada y otras más sencilla. En el opérculo casi siempre los detalles poligonales están mejor definidos. En la capa más interna del endocorion es posible distinguir huellas de una ornamentación poligonal muy simple, la que es fácilmente destruida por la acción de ciertas sustancias químicas empleadas en estas observaciones; se nota aquí que los detalles poligonales no coinciden en superposición con los del exocorión, debido quizá a que estas capas se forman en dos momentos diferentes de la actividad folicular.
Finalmente, revistiendo al huevo se encuentra la cubierta superficial de cemento, producto de secresión de las glándulas colaterales. Dicha cubierta, unas veces es gruesa y permite al huevo adherirse a las superficies donde es puesto, como ocurre en Psammolestes coreodes BERGROTH, 1911, en unas pocas especies del género Triatoma LAPORTE, 1833 según ABALOS y WYGODZINSKY (1951), y en las especies del género Rhodnius STAL, 1859. En otras la cubierta es tan débil que los huevos quedan libres como sucede en las especies que hemos estudiado.
Los huevos de los triatominos presentan un sistema micropilar característico y complicado. Tal sistema está situado entre el contorno del opérculo y el reborde de la abertura opercular; consta de dos tipos de capilares: unos que atraviezan todas las capas del corion y otros sólo parte de él, denominándoseles micrópilos verdaderos y seudomicrópilos, respectivamente. Este sistema ha sido ampliamente estudiado por BEAMENT (1948) en Rh. prolixus. No conocemos referencias de trabajos semejantes en otros triatominos.
RESULTADOS
1. Triatoma infestans.
a) Huevos de especímenes criados en el laboratorio. Largo: 2,691* (2,452 - 2,970) **; ancho: 1,473 (1,350-1,575) y diámetro opercular: 762 (675 - 855).
b) Huevos de especímenes colectados en la naturaleza. Largo: 2,760 (2,498-2,992); ancho: 1,575 (1,418 - 1,687) y diámetro opercular: 768 (675 - 855) .
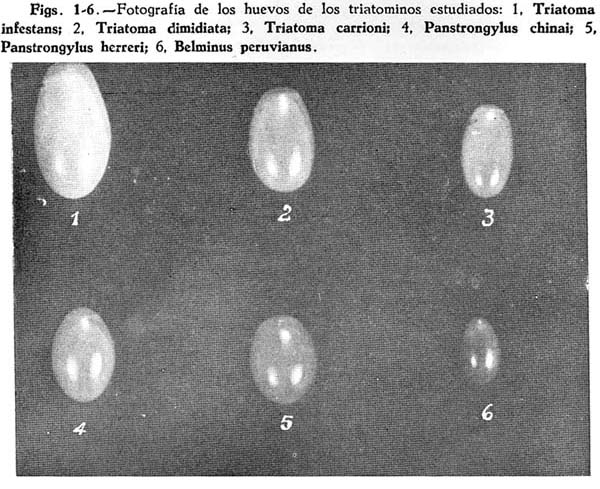
El huevo en ambos grupos de muestras es oval, ligeramente adelgazado en el extremo anterior, simétrico (Fig. 1 ). El cuello está marcado por una ligera constricción de la cáscara. Cuando está recién puesto es de color blanquecino amarillento pero a través del desarrollo embionario va adquiriendo un tono rosado que culmina en un rojo grisáceo cuando va a eclosionar. La cáscara es blanca y opaca, sin embargo permite distinguir algunas estructuras embrionarias.
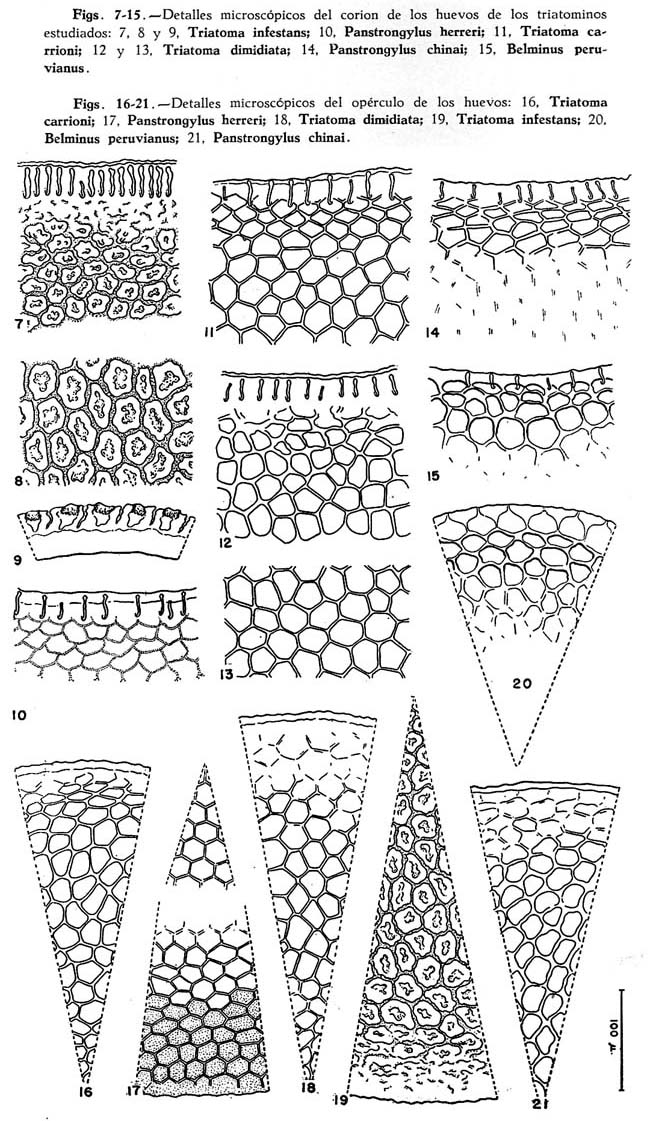
La ornamentación microscópica del corion es idéntica en ambos grupos de muestras. Las celdas poligonales que abarcan toda la superficie, son muy particulares y perceptibles a diferentes aumentos. Los límites de estas celdas son sinuosos y están dados por incisiones profundas a modo de surcos; en el centro de cada celda existe una fosa folicular amplia (Fig. 8). En cortes transversales de la pared del corion se diferencia el endocorion y el exocorion; el primero aparece coloreado con la fucsina, mientras que el segundo permanece incoloro y en él se aprecia la profundidad de las fosas foliculáres y de las incisiones que delimitan las celdas; tales excavaciones atraviezan el exocorion llegando hasta los límites del endocorion (Fig., 9). A nivel del cuello las celdas están incompletas logrando su forma definitiva a medida que se alejan de esta zona (Fig. 7). En el opérculo la ornamentación abarca toda la superficie, es semejante a la descrita para la parte general del coríon pero, en la periferie las celdas están incompletas (Fig. 19).
El huevo de T. infestans puede ser identificado porque el corion en su totalidad, presenta una ornamentación microscópica con celdas de límites sinuosos marcados por incisiones profundas; en el centro de cada celda existe una fosa folicular amplia. Por el aspecto general de la cáscara, blanca y opaca y por su gran tamaño en relación con las demás especies estudiadas.
2. Triatoma carrioni.
a) Huevos de especímenes criados en el laboratorio. Largo: 1,801 (1,689 - 1,960); ancho: 1,008 (945 - 1,085) y diámetro opercular: 557 (508 - 595).
El huevo es de forma oval alargado con extremos suavemente adelgazados, simétrico (Fig. 3). El cuello es casi imperceptible. Recién puesto es de color blanco, posteriormente cuando está por eclosionar adquiere un tono anaranjado rojizo. La cáscara blanco tornasolada, transparente y brillante, permite distinguir al embrión.
El corion de todo el huevo muestra una ornamentación poligonal perceptible a diferentes aumentos; las celdas son predominantemente hexagonales y sus lados están marcados por incisiones lineales que alcanzan sólo la superficie del exocorion. A nivel del cuello existe una formación a modo de "collar" constituido por 3 ó 4 filas de celdas típicas cuyos lados son algo más profundo y gruesos, diferenciándose de las celdas que se inician hacia la parte posterior (Fig. 11 ) . El opérculo muestra detalles poligonales en toda su superficie, semejantes a los de la parte general del corion, aunque en la periferie las celdas están incompletas (Fig. 16). La capa más interna del endocorion, con sus detalles poligonales muy simples, puede observarse en este huevo sólo cuando no ha sido tratado con aclarantes, pues ella es destruída rápidamente. Esta capa es de color marrón, constituida por finísimas puntuaciones y las celdas están delimitadas por una mayor acumulación de este material; estas celdas son de igual forma y tamaño que las del exocorion pero no coinciden en superposición.
El huevo de T. carrioni puede ser identificado por la ornamentación poligonal nítida de todo el corion, destacándose en el cuello un "collar" de celdas, carácter principal que lo diferencia de la especie más parecida: T. dimidiata. La forma oval con los extremos suavemente adelgazados, es otra característica macroscópica notable.
3. Triatoma dimidiata.
a) Huevos de especímenes criados en el laboratorio. Largo: 1,877 (1,680-2,048); ancho: 1,124 (1,059 - 1,199) y diámetro opercular: 678 (621 - 761).
b) Huevos de especímenes colectados en la naturaleza. Largo: 1,878 (1,750 - 2,013); ancho: 1,166 (1,085-1,226) y diámetro opercular:682 (639 - 735) .
El huevo en ambos grupos de muestras es oval, con los extremos redondeados, simétrico (Fig. 2). El cuello está marcado por una ligera constricción. Recién puesto es de color blanquecino amarillento, pero se torna rojo vivo cuando está próximo a eclosionar. La cáscara es blanco tornasolada, algo transparente y brillante.
En ambos grupos de muestras, el corion de todo el huevo presenta una ornamentación microscópica típicamente poligonal, perceptible a diferentes aumentos. Las celdas son predominantemente hexagonales y sus límites están dados por incisiones líneales nítidas que alcanzan sólo la superficie del exocorion (Fig. 13). A nivel del cuello dichas celdas están incompletas y mal formadas, faltando por lo tanto el collar (Fig. 12). En el opérculo los detalles poligonales abarcan toda la superficie y son semejantes a los de la parte general del corion, aunque las celdas periféricas están incompletas como en el cuello (Fig. 18). Los detalles poligonales de la capa más interna del endocorion puede observarse también en este huevo, sin ofrecer particularidades.
El huevo de T. dimidiata puede ser identificado, por la ornamentación poligonal nítida de la totalidad del corion; sin embargo, carece de .collar, detalle que lo diferencia de T. carrioni. Macroscópicamente, la forma oval con extremos redondeados es una característica notable.
4. Panstrongylus herreri.
a) Huevos de especímenes criados en el laboratorio. Largo: 1,635 (1,514-1,838); ancho: 1,266 (1,067-1,330) y diámetro opercular: 739 (682-805) .
b) Huevos de especímenes colectados en la naturaleza. Largo1,789 (1,680-1,900); ancho: 1,322 (1,225-1,382) y diámetro opercular: 792 (761 - 837) .
El huevo en ambos grupos de muestras es de forma oval redondeado, simétrico (Fig. 5). El cuello es casi imperceptible. Recién puesto es de color rosado anaranjado, posteriormente se torna rojo vivo cuando está por eclosionar; en estas circunstancias se observa un halo blanco que rodea el cuello y la periferie opercular. La cáscara es blanco tornasolada, transparente y brillante.
En ambas muestras el corion del cuerpo del huevo *** no muestra ornamentación, excepto a nivel del cuello donde existe un esbozo de collar formado por unas pocas filas de celdas poligonales, de bordes difusos (Fig. 10). En el opérculo en cambio, existen celdas poligonales que abarcan toda la superficie, estas celdas que son más perceptibles en el centro y en la periferie, están delimitadas por líneas finas: sobre las celdas periféricas se aprecia una fina puntuación superficial constituida tal vez por restos de cemento (Fig. 17).
Las características microscópicas del corion del "cuerpo del huevo" varían, cuando éste es tratado durante unas horas con nitrato de plata amoniacal después de haber sido aclarado. En estas condiciones se observa una ornamentación poligonal en toda la superficie, con celdas típicamente hexagonales limitadas por líneas muy finas; sin embargo en el cuello se aprecia el mismo esbozo de collar ya anotado.
En el huevo de esta especie se observa también detalles poligonales en la capa más interna del endocorion.
El huevo de P. herreri puede ser identificado por la ausencia de ornamentación en el corion del "cuerpo del huevo" (tratado según las ,técnicas que se ha empleado en los demás huevos), excepto en el cuello donde existe un esbozo de collar. Macroscópicamente la forma oval redondeada, el gran diámetro opercular en relación al tamaño del huevo y el color rosado anaranjado cuando está recién puesto son caracteres notables.
5. Panstrongylus chinai
a) Huevos de especímenes criados en el laboratorio. Largo: 1,785 (1,601-1,934); ancho: 1,130 (1,050-1,199) y diámetro opercular: 578 (534-639) .
El huevo es de forma oval, con extremos bruscamente adelgazados, simétrico (Fig. 4) . El cuello está marcado por una leve constricción. Recién puesto es de color blanquecino, brillante, poco antes de eclosionar adquiere un tono castaño claro típico. La cáscara es blanca tornasolada, transparente y brillante.
El corion del "cuerpo del huevo" presenta una ornamentación poligonal débil, casi imperceptible. Las celdas están delimitadas por líneas finas, visibles sólo a pequeños aumentos (máximo 100 x); a mayores aumentos se percibe únicamente los lados de las celdas, paralelos al eje mayor del huevo. En el cuello, sin embargo se destaca un collar nítido, formado por 3 ó 4 filas de celdas poligonales, cuyos lados están marcados por líneas gruesas y uniformes, contrastando con las celdas difusas de la parte posterior (Fig. 14) . El opérculo muestra una ornamentación poligonal en toda la superficie; las celdas están delimitadas por líneas precisas, aunque en la periferie las celdas están incompletas (Fig. 21). En el huevo de esta especie también se observa detalles poligonales en la capa más interna del endocorion.
El huevo de P. chinai puede ser identificado por la ornamentación poligonal débil del corion del cuerpo del huevo", y la presencia de un collar formado por celdas poligonales. Macroscópicamente la forma oval con extremos afilados y el color castaño claro cuando está por eclosionar, son características importantes.
Si se considera la ornamentación microscópica del corion del cuerpo del huevola especie más parecida sería B. peruvianus, pero los detalles ornamentales de los opérculos y el tipo de celdas que forman el collar, permite diferenciarlos. Así mismo, el tamaño de los huevos es notablemente diferente.
6. Belminus peruvianus.
a) Huevos de especímenes criados en el laboratorio. Largo: 1,169 (1,155-1,339); ancho: 733 (621-805) y diámetro opercular: 379 (306 - 472).
Es el huevo de menor tamaño dentro de las especies estudiadas, de forma oval ligeramente adelgazado en el extremo anterior, simétrico (Fig. 6). El cuello está marcado por una leve constrición de la cáscara. Recién puesto es de color blanquecino brillante, variando hasta un rojo vivo cuando está por eclosionar. La cáscara es blanca tornasolada, transparente y brillante.
La superficie del corion del "cuerpo del huevo" muestra una ornamentación poligonal débil con celdas delimitadas por líneas finas, casi imperceptibles. Sin embargo a nivel del cuello se destaca un "collar" formado por 3 ó 4 filas de celdas típicas de contorno casi circular y cuyos límites no son uniformes, sino que están dados por una línea gruesa hacia el extremo anterior y otra fina hacia el extremo posterior; en conjunto las celdas del "collar" aparecen en relieve (Fig. 15). En el opérculo la ornamentación es parcial circunscrita a la periferie; las celdas son de dos clases: las periféricas están delimitadas por líneas muy finas y uniformes, luego unas 3 6 4 filas de celdas iguales a las que forman el "collar" y nuevamente celdas finas que van desapareciendo hacia el centro (Fig. 20) . En la capa más interna del endocorion se aprecia también la ornamentación poligonal sencilla.
El huevo de B. peruvianus puede ser identificado por la ornamentación poligonal casi imperceptible del corion del "cuerpo del huevo", excepto en el cuello donde se destaca un "collar" de celdas casi circulares y de contorno diferenciado; igualmente por la ornamentación periférica del opérculo. Macroscópicamente el menor tamaño del huevo en relación con las otras especies, es determinante.
Clave para la identificación de los huevos de los triatominos estudiados.
1. Celdas del corion de todo el huevo bien perceptibles, delimitadas por líneas nítidas (Figs. 8, 11 y 13) Celdas del corion del "cuerpo del huevo" casi imperceptibles (Figs. 10, 14 y 15)
2. Celdas del corion sin estructuras particulares en su interior (Figs .11 y 13 ). Celdas del corion con una fosa folicular en su interior; las celdas son de contorno más o menos sinuosos, delimitadas por incisiones profundas a modo de surcos (Figs. 7, 8 y 9). Triatoma infestans (Klug, 1834).
3. En el cuello, celdas diferenciadas forman un "collar" del que continúa la ornamentación poligonal del resto del coríon (Fig. 11). Triatoma carrioni Larrousse, 1926.
En el cuello las celdas están incompletas y mal formadas faltando por lo tanto el "collar"; hacia la parte posterior va apareciendo la ornamentación poligonal definitiva del resto del corion (Figs. 12 y 13). Triatoma dimidiata (Latreille, 1811)
4. En el cuello se destaca un "collar" formado por 3 ó 4 filas de celdas delimitadas por líneas nítidas (Figs. 14 y 15). Opérculo con celdas en toda la superficie, o sólo en la periferie (Figs. 20 y 21 )
En el cuello se aprecia un esbozo de "collar" formado por celdas de límites difusos (Fig. 10). Opérculo con celdas poligonales y fina puntuación superficial periférica (Fig. 17). Panstrongylus herreri Wyqodzinsky, 1948.
5. "Collar" formado por celdas típicamente poligonales, con lados uniformes (Fig. 4). Opérculo con ornamentación poligonal en toda la superficie (Fig. 21) Largo del huevo de 1.600-1,900 micras. Panstrongylus chinai (Del Ponte, 1929).
"Collar" formado por celdas de contorno casi circular; el límite de cada celda está dado por una línea gruesa hacia la parte anterior y otra fina hacia la posterior (Fig. 15). Opérculo con ornamentación parcial periférica (Fig. 20). Largo del huevo de 1,1501,340 micras. Belminus peruvianus Herrer et al., 1954.
DISCUSION
1. Diferencias morfológicas entre los huevos de las especies criadas en el Laboratorio. Los resultados expuestos indican que en lo referente a las dimensiones, los valores promedios de los huevos de algunas especies son sensiblemente semejantes. A fin de apreciar el significado estadístico de estos datos, se ha creído necesario comparar todas las especies, de acuerdo a cada una de las tres dimensiones, mediante el "Análisis del Varianza".
Según este análisis, los huevos de las especies comparadas son estadísticamente diferentes en el largo, ancho y diámetro opercular, percibiéndose también que la variación que existe entre especies es muy superior a la que hay dentro de ellas, consideradas individualmente.
Aunque los caracteres microscópicos del corion de los huevos de estos triatominos están constituidos por una ornamentación específica de suficiente valor diferencial, es importante seguir una misma técnica ya que como se ha observado en P. herreri los resultados varían al emplear técnicas distintas, hecho éste que podría explicar quizá las diferencias que existen entre esquemas de una misma especie dados por diferentes autores.
2. Observaciones comparativas entre huevos de especímenes criados en el laboratorio y colectados en la naturaleza. Como ya se ha dicho sólo en T. infestans, T. dimidiata y P. herreri se han efectuado observaciones de esta índole. Los resultados indican que en cuanto a las dimensiones, existen diferencias entre los datos obtenidos de ambas muestras de una misma especie. Para apreciar el valor estadístico de estas diferencias, se ha realizado un análisis comparativo en cada caso, empleando el "Método del significado estadístico de la diferencia entre dos promedios".
De acuerdo a este análisis, en T. dimidiata solamente en el ancho la diferencia es significativa ya que la probabilidad hallada de que ambos grupos sean iguales es muy remota (menor de 1 por mil), mientras que en el largo y diámetro opercular las diferencias carecen de valor. En T. infestans, en el largo y en el ancho las diferencias son significativas (menores de 1 por mil), no así en el diámetro opercular. Y finalmente en P. herreri, las diferencias son significativas en las tres dimensiones (menores de 1 por mil). Estos hallazgos indicarían que estadísticamente existen diferencias en las citadas dimensiones entre los huevos de ambas muestras. Sin embargo los caracteres microscópicos del corion permanecen invariables en los huevos de ambos grupos. Tales resultados eran de esperar ya que variaciones de este tipo implicarían modificaciones de la estructura histológica del folículo ovárico, puesto que la ornamentación microscópica del corion no es sino la huella dejada por el epitelio folicular que lo formó.
3. Apreciaciones generales. Lo expuesto permite apreciar que los huevos de todas las especies procedentes de crías de laboratorio, poseen caracteres morfológicos macroscópicos y microscópicos específicos. Sin embargo cuando se trata de poblaciones de una misma especie, procedentes de ambientes distintos y que como en nuestro, caso son el laboratorio y la naturaleza, se observan variaciones considerables que afectan en mayor o menor grado las dimensiones del huevo; pero mientras se producen variaciones de esta índole los caracteres microscópicos del corion permanecen idénticos. Creemos pues, que al efectuar estudios morfológicos de este tipo, con miras taxonómicas se debería tener en cuenta la procedencia del material y considerar la ornamentación microscópica del corion como carácter fundamental, en tanto que las dimensiones constituirían caracteres complementarlos relativos, sujetos a fluctuaciones. No obstante las tres dimensiones en conjunto podrían caracterizar la "forma específica del huevo".
AGRADECIMIENTOS
Este trabajo se ha realizado bajo la dirección del Dr. Arístides Herrer, Jefe de la División de Estudios Epidemiológicos e Investigaciones Especiales del Instituto Nacional de Salud Pública, a quien expreso mi profundo agradecimiento; así como al Sr. Raúl Vargas, del Departamento de Bioestadística del Servicio Cooperativo Interamericano de Salud Pública por su generosa colaboración en la parte estadística, al Br. Fortunato Blancas por sus sugerencias en la confección de la clave y a todas las personas que me han brindado su gentil ayuda.
*En todas las especies las medidas se dan en micras.
**Las cifras encerradas entre paréntesis constituyen los valores extremos encontrados.
***Se denomina corion del "cuerpodel huevo" a la cáscara sin opérculo.
REFERENCIAS
1. ABALOS, J. W. y WYGODZINSKY, P. Las Triatominae argentinas (Reduviidae, Hemiptera). Inst. Med. Reg. Univ. Nac. Tucumán. Publ. No. 601, Monogr. No. 2, pp. 4042. 1951 [ Links ]
2. BEAMENT, J. W. L. The formation and structure of the chorion of the egg in an Hemipteran, Rhodnius prolixus. Quart. J. Micr. Sci. 87: 393-439. 1946 [ Links ]
3. BEAMENT, J. W. L. The formation and structure of the micropylar complex in the eggshell of, Rhodnius proliuxs Stahl (Heteroptera reduviidae). Jour. Exp. Biol. 23: 213-233. 1947 [ Links ]
4. BEAMENT, J. W. L. The penetration of the insect egg-shell. I. Penetration of the chorion of , Rhodnias prolixus. Bull. Ent. Res. 39; 359-383. 1948. [ Links ]
5. COMSTOCK, J. H. An introduction to Entomology. 2a. ed. Ithaca, N. Y., The Custock Publishing Co. pp. 166-168. 1925 [ Links ]
6. DOETSCHMAN, W. H. Some suggestions in microthecnique particulary useful in microentomology and parasitology. Trans. Am. Micr. Soc. 63: 175-178. 1944 [ Links ]
7. FROST, S. W. General Entomology. New York, McGraw-Hill Book Company, Inc. pp. 117-128. 1942 [ Links ]
8. HERRER, A.; UNT, H. y WYGODZINSKY, P. Contribución al conocimiento del género Belminus Stal, 1859 (Triatomi nae, Reduviidae, Hemiptera). An. Inst. Med. Reg. 4: 85-105. 1954 [ Links ]
9. HERRER, A. Trypanosomiasis americana en el Perú. V. Triatominos del valle interandino del Marañón. Rev. Med. Exp., Lima. 9: 69-81. 1955 [ Links ]
10. IMMS, A. D. A General textbook of Entomology. 7a. ed. New York, E. P. Dutton Co. pp. 161-162.1948. [ Links ]
11. LENT, H. Comentarios sobre o genero ",Rhodnius" Stal, com descricao de uma nova especie do Brasil. Rev. brasila Biol. 14: 237-247. 1954 [ Links ]
12. LIZARASO, YOLANDA. Nota sobre hallazgo de Triatoma dimidiata y Panstrongglus rufotuberculatus en el Perú. Rev. Med. Exp., Lima. 9: 119-121. 1955 [ Links ]
13. RICHARDS, A. G. The Integument of arthropods. Minneapolis, University of Minnesota Press. pp. 279-280. 1951 [ Links ]
14. SNEDECOR, G. W. Statistical Methods. 4a. ed. Iowa, The Iowa State College Press. 486 pp. 1948 [ Links ]
15. TICSE, ABILIA. Observaciones experimentales sobre el ciclo biológico de Triatoma infestans (Klug 1834). Tesis, Fac. Ciencias Univ. Nac. May. San Marcos, Lima, 67 pp. 1951 [ Links ]
16. USINGER, R. L. The triatominae of North and Central America and the West Indies and their public health significance. U. S. Public Health Service. Public Health Bulletin No. 288. 88 pp. 1944 [ Links ]
17. WIGGLESWORTH, V. B. The principles of insect physilology. 5a. ed. London, Methuen & Co. pp. 1-6; 459 - 464. 1953 [ Links ]
18. WYGODZINSKY, P. Elenco sistemático de los Reduviiformes americanos. Inst. Med. Reg. Univ. Nac. Tucumán. Publ. No. 473, Monogr. No. 1. 102 pp. 1949 [ Links ]

