










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.16 n.1-2 Lima 1999
ARTÍCULOS ORIGINALES
Clonación y Transcripción del Gen EBER-1 del Virus Epatein-Barr: estudio de su expresión genética en linfomas por hibridación in situ
Guerrero I1; Velazco R1; Mejía R1; Zaharia M2; Sarria G2; Misad O3; Martínez M3; Wachtel A3; Cordero R4; Casanova L4.
1 Unidad de Biología Molecular - Centro de Investigación en Cáncer Maes–Heller.
2 Dpto. de Radioterapia - Dpto. de Patología, Instituto de Enfermedades Neoplásicas Dr. Eduardo Cáceres Graziani.
3 Dpto. de Pediatría, Instituto de Enfermedades Neoplásicas Dr. Eduardo Cáceres Graziani.
4 Dpto. de Medicina, Instituto de Enfermedades Neoplásicas Dr. Eduardo Cáceres Graziani.
RESUMEN
La capacidad del virus de Epstein Barr (VEB) de inducir transformación e inmortalización de linfocitos y su alta prevalencia en algunas neoplasias linfoides ha motivado intensa investigación de su biología y comportamiento. Describimos la hibridación in situ (HIS), para la identificación de transcriptos del virus de Epstein-Barr, desarrollado en la Unidad de Biología Molecular del Centro de Investigación en Cáncer Maes - Heller, en biopsias de linfoma incluidas en parafina.
El gen EBER-1(Epstein Barr-encoded RNA) fue clonado y transcrito, el ARN transcrito fue evaluado con óptimos resultados en las líneas celulares Raji y Daudi que expresan EBER. Este transcrito marcado fue usado como sonda para determinar la expresión genética del virus en 159 especímenes biológicos en estudio y en controles positivos y negativos conocidos empleando la HIS. La actividad transcripcional del virus fue observada en 44% de los casos estudiados. Concluimos que la HIS en tejido incluido en parafina es una buena alternativa para detectar actividad del VEB y la presencia o ausencia de EBER puede ser considerada como marcador biológico en la etiopatología de estos linfomas.
Palabras Clave: Epstein Barr, hibridación, Linfomas.
ABSTRACT
The capacity of Epstein-Barr virus (EBV) to induce transformation and immortalization of lymphocytes and its high prevalence in some lymphoid neoplasias has encouraged research on its biology an behavior. We describe the in situ hybridization (ISH) for Epstein-Barr virus transcript identification, developed in the Molecular Biology Unit of the Centro de Investigación en Cáncer Maes-Helller, in biopsies of paraffin-embedded lymphoma. The EBER-1 gen (Epstein Barr-encoded RNA) was cloned and transcribed, the transcript-RNA was evaluated with good results in Raji and Daudi cellular lines. This labeled transcript was used as a probe to determine the genetic viral expression in 159 biological specimens studied in positive and negative controls applying ISH. The virus transcriptional activity was observed in 44% of the cases studied. We conclude that ISH in paraffin-embedded tissue is a good alternative to detect EBV activity, and the presence or absence of EBER could be considered as a biological marker in the etiopathology of these lymphomas.
Key Words: Epstein Barr, hybridization, Lymphomas.
INTRODUCCIÓN
El virus de Epstein-Barr (VEB), primer virus tumoral humano, fue descubierto hace más de 30 años en líneas celulares de un tumor agresivo prevalente en niños africanos, conocido como Linfoma de Burkitt1. Desde entonces el virus ha sido ampliamente considerado como agente etiológico de enfermedades linfoproliferativas como los linfomas de Hodgkin y no Hodgkin, linfomas post-transplante, linfomas asociados al síndrome de inmunodeficiencia adquirida-SIDA, carcinoma nasofaríngeo y, más recientemente, cáncer de mama2,3.
La prevalencia del virus en estos tumores ha motivado intensa investigación de su biología, y se ha podido inducir transformación de linfocitos B (LB) por el virus y, con menos éxito, en linfocitos T (LT). Estos hallazgos han permitido establecer que el virus puede producir infección lítica e infección latente en las células que infecta4.
La infección lítica se desarrolla sin mayores cambios, en las células de la orofaringe. La infección latente está dirigida por la expresión de 11 de sus genes, EBER-1, EBER-2, EBNA-LP, EBNA-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C y LMP-1, LMP-2A y LMP-2B. Algunos de estos genes, que se expresan en los LB o LT con infección latente, se comportan como oncogenes, el EBNA-2 es un potente transactivador viral, LMP-1 induce proliferación celular, EBNA-LP puede unirse a los productos de los genes supresores de tumores p53 y pRb; un gen que se expresa en ambos tipos de infección, el BHFR-1, es un homólogo de bcl-2, comportándose como un inhibidor de la muerte celular programada o apoptosis. Recientemente se ha descubierto que EBER bloquea la muerte celular programada5, punto importante y de interés nuestro para determinar la respuesta del paciente al tratamiento. Sin embargo, a pesar de las evidencias, el rol del virus aún no es universalmente aceptado como factor causal en algún tipo de cáncer humano. El 95% de la población mundial es portadora del VEB, con mayor frecuencia del tipo 1 o A, sin embargo en África, y sorpresivamente en nuestra población, existe una alta prevalencia del tipo 2 o B, tanto en personas aparentemente sanas como en aquellas portadoras de tumores6; sin embargo, es necesario continuar el estudio en poblaciones mayores.
Se ha descrito métodos muy sensibles para estudiar la prevalencia del VEB, desde screening con anticuerpos monoclonales e hibridación in situ hasta la PCR de altísima sensibilidad para su detección; sin embargo, muchos investigadores postulan que podría tratarse sólo de un ocasional espectador.
Nuestro objetivo ha sido determinar la expresión del virus en un grupo de linfomas, con estudio previo de detección de VEB por PCR, y su posible rol en la linfomagénesis, para lo cual nos propusimos desarrollar un método de hibridación práctico y sensible, basado en la síntesis y marcaje no radioactivo de ARN antisentido, el que usamos como sonda para la hibridación in situ, y que nos permita detectar el transcripto EBER-1 en células neoplásicas, considerando que se expresa en los tres tipos de latencia que presenta el virus.
MATERIALES Y MÉTODOS
Fue seleccionado, para clonar, el fragmento BamHI C de la cepa B95-8 que contiene el gen EBER del VEB en el plásmido pBluescrip II(pBSII) que permite clonar fragmentos de ADN de hasta 10 Kb, para sintetizar in vitro el ARN antisentido marcado, que fue hibridado como sonda marcada empleando la HIS como herramienta de diagnóstico molecular en el grupo de estudio.
El material biológico del estudio estuvo constituido por 159 muestras de linfomas de diferente procedencia, de pacientes admitidos al Instituto Nacional de Enfermedades Neoplásicas Dr. Eduardo Cáceres Graziani, distribuidos de la siguiente manera: 108 linfomas nasales, 23 orbitarios, 16 paranasales y 12 cutáneos. El criterio de inclusión de este grupo de linfomas fue haber tenido estudio molecular previo para la detección del virus Epstein-Barr mediante PCR.
CLONACIÓN DEL GEN EBER-1 DEL VEB
El plásmido recombinante pBR322, conteniendo un fragmento BamHI C del ADN del VEB, fue digerido con la enzima de restricción EcoRI, los fragmentos digeridos fueron resueltos en un gel de agarosa (Perkin Elmer) al 0,8% y recuperados, posteriormente fueron ligados al plásmido-pBS II (Stratagene) con la ligasa del T4 (Sigma), transfectados en la cepa E.coli XL1-Blue MRF (Stratagene); las cepas recombinantes fueron seleccionadas por screening de color (lacZ), hibridación y confirmadas por PCR7.
TRANSCRIPCIÓN DEL GEN EBER-1/VEB
El plásmido recombinante pBSEBER-1, obtenido con el procedimiento anterior, se usó para sintetizar ARN antisentido corriente, arriba del promotor T7 (transcripción dirigida por el promotor T7). Para la síntesis del ARN antisentido se empleó un sistema de transcripción comercial (Roche ® ), el ARN (-) fue marcado con digoxigenina, el marcaje fue determinado y cuantificado por un blotting usando un sistema de detección no radiactiva (Roche® ).
HIBRIDACIÓN IN SITU
De cada caso estudiado se tomó cortes de tejido incluido en parafina de 5 µ de espesor, obtenidos tomando las precauciones necesarias para evitar la contaminación, aplicando medidas de seguridad después de cada manipulación y depositados sobre láminas pre-tratadas. Los cortes fueron inicialmente desparafinados, tratados con pronasa y condiciones de pre-hibridación, posteriormente se hibridó con la sonda marcada a una temperatura de 55°C, finalmente fueron sometidos a condiciones de post-hibridación y detección del híbrido con anticuerpo monoclonal de marcaje enzimático8. Se consideró un caso positivo a la evidencia de una reacción intranuclear de color púrpura que indicó la presencia del transcripto en estudio. Se utilizó controles positivo, negativo y blanco, por grupo de estudio trabajado. Como control positivo para EBV se consideró la línea celular Raji y Daudi, como control negativo un paciente negativo conocido, y como blanco un caso conocido en el cual se reemplazó la sonda por agua, en la reacción de hibridación.
RESULTADOS
Las cepas recombinantes fueron seleccionadas mediante una prueba de color e hibridación no radioactiva. La talla de los insertos osciló entre 0,6 – 5,0 Kb (Figura1).
Mediante la PCR confirmamos los insertos, presencia del gen EBER en los plásmidos recombinantes correspondientes a una fracción de 150 pb del gen(Figura 2)
La mejor concentración de los pBSEBER fueron determinadas mediante un blotting, tomando como referencia el control positivo comercial (Roche ® ), pudiéndose detectar concentraciones que oscilaron entre 10 ng y 1 pg, observándose en uno de los plásmidos (D) las concentraciones mas bajas de hasta 10 pg (Figura 3).
Por HIS se detectaron transcriptos en las líneas celulares control para VEB Raji y Daudi, aplicando como sonda el pBSEBER-D y usando concentraciones de hasta 1pg (ver tabla 1), para determinar la concentración adecuada a usarse en el estudio de la expresión genética del VEB en los 159 linfomas.
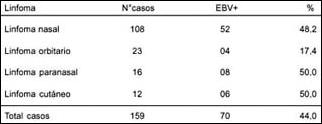
De los linfomas estudiados, correspondieron 108 de ellos a la región nasal, 23 a la región orbitaria, 16 paranasales y 12 cutáneos. Aplicando la hibridación in situ como herramienta de diagnóstico molecular en este grupo de linfomas, y utilizando la sonda preparada, detectamos transcriptos del gen EBER-1 del virus Epstein Barrr en 70/159(44%) casos, los cuales mostraron un patrón de distribución intranuclear típico de las células tumorales(figura4).
Los linfomas que albergaron EBER-1 de acuerdo a su procedencia pueden ser observado en la tabla 2.
Figura 1. Talla de los insertos. Electroforesis de ADN plasmídico, una
alícuota del ADN extraído fue corrido en un gel de agarosa al 0,8% para
evaluar su talla en pares de bases: P1-P4 (plásmidos), M (marcador de
peso molecular)
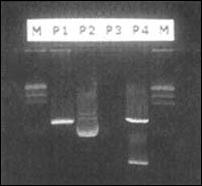
Figura 2. PCR para EBER-1. Una alícuota del producto amplificado del
gen EBER-1 fue corrido en un gel de agarosa. En la foto se muestra la
banda correspondiente de 150 pb. P1-P4 (plásmidos), M (marcador de
peso molecular).

Figura 3. Blotting de concentración de la sonda EBER-1. El ARN antisentido
transcrito fue fijado, como se observa, a una membrana de nylon con luz
UV, el marcaje fue detectado inmunológicamente. En la foto se muestran
las manchas Dot de acuerdo a la concentración de ARN.
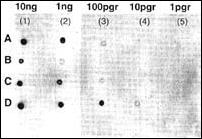
Figura 4. Hibridación in Situ para EBER-1 en linfomas. En la foto se
observa claramente células tumorales con expresión genética viral.
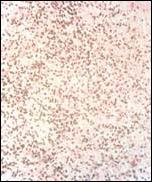
Tabla 1. Concentración de la sonda para HIS vs respuesta por línea celular.
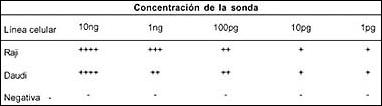
Tabla 2. Frecuencia del transcripto EBER-1 detectado por HIS en linfomas.
DISCUSIÓN
Determinar la presencia de transcriptos virales en las neoplasias y, más aún, conociendo que el virus de Epstein Barr es capaz de inducir transformación, multiplicación e inmortalización de las células blancas9,10, contribuyendo de esta forma a la linfomagénesis en humanos, resulta de gran importancia.
Cuando realizamos el screening de los recombinantes, aprovechamos la característica del plásmido pBluescript cuya zona de inserción se ubica en una secuencia que codifica un péptido para B-galactosidasa, quedando trunca la expresión de éste, de tal forma que las bacterias recombinantes produzcan colonias blancas en un medio que contenga IPTG y xGAL de fácil reconocimiento por esta simple prueba de color. Los transcriptos del plásmido pBSEBER-D nos dieron las mejores concentraciones detectables desde 10ng hasta 10 pg; confirmamos indiscutiblemente la utilidad del método no sólo para clonar fragmentos de ADN con cierta facilidad, sino también nos permitió preparar nuestra propia sonda, sobre todo de ARN cuyo aislamiento y marcaje resulta, muchas veces, laborioso.
Si bien es cierto la frecuencia total detectada del transcripto EBER-1 en el grupo de linfomas que hemos estudiado, proveniente de diferente región, aporta información general de su actividad transcripcional durante la latencia del VEB en linfocitos tumorales, el análisis de estos hallazgos por área de procedencia proporciona información importante del posible rol de observador o etiológico que pueda cumplir el virus en estos linfomas, y hace necesario continuar investigándolo.
La alta frecuencia en la detección de la expresión del gen EBER-1 del virus Epstein Barr, que hemos hallado en el grupo de linfoma nasal, coincide con los descritos en la literatura asiática e incluso con reportes de nuestra población en casuísticas pequeñas11,12, lo cual sustenta el rol de este virus en el proceso de linfomagénesis de esta neoplasia.
Muy por el contrario los reportes a nivel mundial sobre transcriptos del gen EBER-1 por otros investigadores en relación a los linfomas paranasales, cutáneos y orbitarios son muy escasos a nivel mundial, e inexistentes a nivel nacional.
Cuando observamos el grupo de linfomas de procedencia paranasal, encontramos que la mitad de ellos presentó expresión viral, hallazgo similar al detectado por Peh en un grupo de pacientes con linfoma asociado al síndrome de inmunodeficiencia adquirida13,14. Estos resultados nos indican que, en los linfomas del tracto respiratorio superior, el virus de Epstein Barr presenta una prevalente actividad transcripcional que confirma que está fuertemente asociado con los eventos de linfomagénesis de localización nasal y paranasal en nuestra población.
Considerando los escasos reportes para linfoma cutáneo y EBER1-HIS, nuestros resultados revelan una mayor frecuencia de transcriptos del VEB en células tumorales en relación a otros hallazgos15, esto nos permite afirmar que el VEB juega un rol importante en la patogénesis de este linfoma.
La expresión del VEB fue un hallazgo menos frecuente en nuestros casos de linfoma orbitario, no por ello menos importante, porque se puede inferir que el virus no tenga un rol mayor en su desarrollo, hallazgo algo similar detectado por Chan para ADN del VEB en uno de trece linfomas primarios de órbita16, siendo aún necesario su estudio en casuísticas mayores en nuestra población.
Si bien es cierto la RT-PCR es el método molecular más sensible disponible hasta el momento para estudiar actividad del VEB como ha sido reportado por otros investigadores, es todavía un problema extraer ARN de calidad del tejido tumoral incluido en parafina, lo cual hace aún inaccesible su aplicación en estudios retrospectivos. Nuestros hallazgos demuestran que la HIS es una buena alternativa de elección para detectar transcriptos del virus Epstein Barr en estudios retrospectivos de biopsias de tejido incluidos en parafina cuando se dispone de una sonda específica como EBER-1 que, por la frecuencia de su expresión en los tres estados de latencia del VEB17, facilita su detección temprana. Podemos concluir que la presencia y/o ausencia del transcripto del virus de Epstein Barr puede ser considerado como marcador biológico en la etiopatología de estos linfomas y, si complementamos el estudio de la expresión del gen EBER-1 en linfomas con el estudio inmunohistoquímico de ciertas proteínas virales, podríamos determinar el tipo de latencia viral en este tipo de tumores.
REFERENCIAS BIBLIOGRÁFICAS
1. Epstein M, Achong B, Barr Y. Virus particles in cultured lympho-blasts from Burkitts lymphoma. Lancet 1964; 1: 702-3. [ Links ]
2. Lyons S, Liebowitz D. The roles of human viruses in the pathogenesis of lymphoma. Sem Oncol 1998; 25: 161-75. [ Links ]
3. Magrath I, Bathia K. Breast cancer : a new Epstein-Barr virus-associated disease?. J Nat Ca Inf 1999; 91: 1349-50. [ Links ]
4. Kieff E. Epstein-Barr Virus and the replication. En Fields Virology 3ra. ed. Fields B, Knipe D, Howley P y col. Lippincott-Raven editores, Filadelfia 1996: 2343-96. [ Links ]
5. Zur Hausen H. Viruses in human cancers. Eup J Ca 1999; 35: 1174-81. [ Links ]
6. Guerrero I. Centro de Investigación en Cáncer Maes Heller. Datos no publicados. [ Links ]
7. Sambrook J, Fritsch E, Manniatis T. Molecular cloning : A laboratory manual. 2da.ed. Col Spring Harbor Laboratory. NY, USA. 1989. [ Links ]
8. Chang K, Chen Y, Shibata D, Weiss L. Description of an in situ hybridization methodology for detection of Epstein-Barr Virus RNA in paraffin-embedded tissues, with a survey of normal and neoplastic tissues. Diag Mol Pathol 1992; 1: 246-55. [ Links ]
9. Radha K, Shanthi P, Madhavan M. A study of association of Epstein-Barr-virus with Hodgkins dosease. Ind J Pathol Microbiol 1997; 40: 351-4. [ Links ]
10. Rickinson A.B. Epstein -Barr Virus in action in vivo. New Engl J Med 1998; 338 (20): 1461-3. [ Links ]
11. Arber DA, Weiss LM, Albujar PF, Chen Y-Y, Jaffe ES. Nasal lymphomas in Peru: high incidence of T-cell immunophenotype and Epstein Barr virus infection. Am J Surg Pathol 1993; 17: 392-9. [ Links ]
12. Guerrero I, Francklin JL, Naresh KN, Cáceres E, Bathia K, OConnor G, et al. P53 expression, bcl-2 expression and EBV association in peruvian nasal lymphomas. Blood 1994; 84 (Suppl 1): 522ª. [ Links ]
13. Peh SC, Sandvej K, Pallesen G. Epstein-Barr virus (EBV) in Malaysian upper-aerodigestive-tract lymphoma: incidence and subtype. Int J Cancer 1995; 61: 327-32. [ Links ]
14. Luzi P, Leoncini L, Funto I, et at. Epstein-Barr virus infection in sinonasal non-Hodgkins lymphomas. Virchows Arch 1994; 425: 121-5.
15. Hirose Y, Masaki Y, SSAki K, et at. Determination of Epstein-Barr virus association with B-cell lymphomas in Japan: study of 72 cases-in situ hybridization, polymerase chain reaction, immunohistochemical studies. Int J Hematol 1998; 67: 165-74.
16. Chan Chi-Chao. Detection of Human Herpes virus-8 and Epstein-Barr virus DNA in primary intraocular lymphoma. Blood 1999; 93(8): 2749-51. [ Links ]
17. Bella C, Santón A, Plaza G. Virus Epstein Barr (VEB) y neoplasia. Hospital Universitario Ramón y Cajal, comunicación personal. 1997. [ Links ]
Correspondencia: Ivonne Guerrero. Instituto Nacional de Salud.
Calle Cápac Yupanqui 1400, Lima 11, Perú. Apartado postal 471.
Telf.: (0511) 4719920 - Fax: (0511)4710179.
Email: iguerrero@inen.sld.pe

