










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.17 n.1-4 Lima 2000
COMUNICACIÓN CORTA
Caracterización Molecular de la Secuencia parcialdel Gen de la Glicoproteína NS1 del Virus Dengue 1 proveniente de Máncora, Perú
Carlos Yábar Varas1
1 División de Biología Molecular, Centro Nacional de Laboratorios en Salud Pública, Instituto Nacional de Salud.
RESUMEN
Se caracterizó una región genética que codifica la glicoproteína NS1 del virus dengue 1 proveniente de Máncora, Piura. Comparaciones de secuencias de nucleótidos revelaron un 93,32% de identidad entre el aislamiento peruano y una cepa de Hawai. A nivel de aminoácidos, se observaron cambios de tipo no conservativos en dominios epitópicos de reconocimiento humoral. De otro lado, el perfil hidropático de la región estudiada fue similar al de otros aislamientos referenciales. Los resultados sugieren realizar mayores análisis de identidad genética y mutaciones en dominios epitópicos en el virus dengue 1 peruano.
Palabras claves: Virus del dengue; Proteínas no estructurales virales; Secuencia de bases; Perú (fuente: BIREME).
ABSTRACT
A 419bp-NS1 genetic region corresponding to Dengue virus 1 was characterised from an outbreak in Máncora Piura. The comparison of nucleotide sequences revealed that Peruvian isolates showed high correlation (93.32%) with a Hawaii strain. Amino acids comparisons revealed non-conservative changes into humoral response epitope domains. On the other hand, the hydropathy profile of NS1 was similar to other referential strains. The results suggest that more comparisons are needed regarding genetic identity and mutations into the epitope domain of Peruvian dengue 1 virus.
Key words: Dengue virus; Viral nonstructural proteins; Base sequence; Peru (source: BIREME).
El Dengue es una enfermedad causada por un virus perteneciente a la familia Flaviviridae, el cual es transmitido al hombre a través de la picadura de un mosquito del género Aedes. La enfermedad se manifiesta a través de tres síndromes definidos como el Dengue clásico o benigno (DC), la Fiebre hemorrágica por Dengue (FHD) y el Síndrome de Shock por Dengue (SSD)1.
El dengue en el Perú es actualmente un grave problema de salud pública. Uno de los principales brotes de dengue clásico ocurrió en la ciudad de Máncora, el año de 1997. Todos los casos reportados correspondieron a Dengue 12.
A nivel molecular el virus presenta diez genes, de los cuales siete codifican proteínas no estructurales. La primera proteína no estructural es conocida como NS1. La importancia de NS1 radica en sus propiedades antigénicas, inmunogénicas y su posible relación con manifestaciones hemorrágicas en pacientes infectados 3-6.
En el presente estudio, se analizó una región del gen de la glicoproteína NS1 (del inglés nonstructural 1) del virus Dengue 1 peruano a partir del brote de Máncora. El objetivo fue caracterizar una región de 419 pb que codifica la porción carboxiterminal de NS1 del virus Dengue 1 peruano y compararla con otras cepas referenciales de Dengue del mismo serotipo reportadas en el banco de genes 7-9. La metodología para la amplificación de este fragmento en el Perú ha sido descrita previamente10, y la secuencia correspondiente a NS1 fue reportada en el Banco de Genes11.
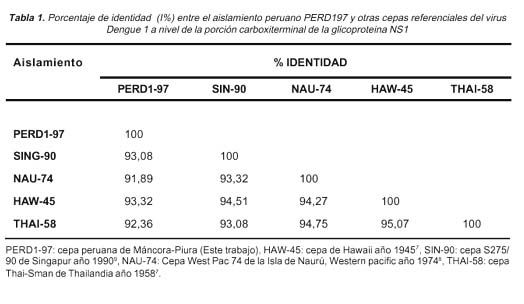
De acuerdo a los datos de comparación de secuencias de nucleótidos, el mayor porcentaje de identidad (% de nucleótidos idénticos) fue hallado entre PERD1-97 y la cepa de Hawai-1974 (Tabla 1) con un índice de 93.32%. Estos datos se relacionan con un parentesco filogenético hallado entre un aislamiento peruano del año 1990 y otro proveniente de Hawaii de 1944 12. Considerando que las secuencias analizadas en este estudio pertenecen a aislamientos de años distintos a los referidos anteriormente, no es posible determinar que PERD1-97 esté relacionado filogenéticamente a Hawai-1945. Sin embargo es probable que el mismo genotipo de dengue 1 peruano de 1990 halla permanecido latente en la naturaleza por lo menos hasta 1997, año en que fue caracterizado PERD1-97. En consecuencia, sería importante realizar un estudio de comparación entre PERD1-97 y el aislamiento peruano caracterizado en 1994, a fin de hallar algún tipo de relación filogenética entre ambos aislamientos.
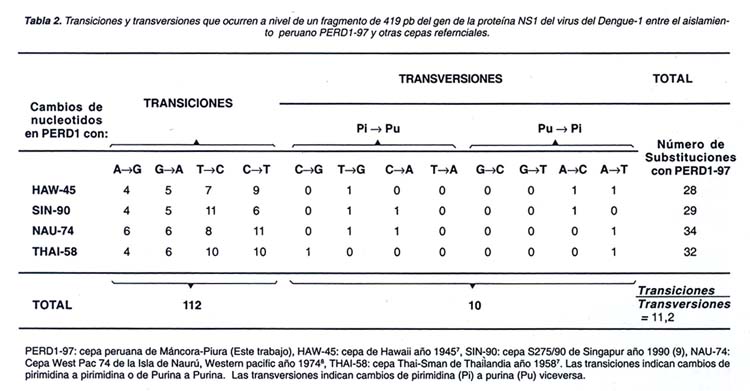
De otro lado,111 transiciones fueron observadas frente a 10 transversiones entre PERD1-97 y los cuatro aislamientos de referencia (Ver Tabla 2). La proporción de 11:1 hallado en este estudio concuerda con trabajos previamente publicados 13 donde se menciona que este tipo de substituciones nucleotídicas probablemente responda a una tasa de error generada por la propia ARN polimerasa del virus durante los procesos de incorporación de purinas y pirimidinas para la replicación de ARN genómico. Eventualmente este proceso tiene una tasa de error relativamente pequeña y asegura la supervivencia del virus a la alta presión de la selección natural.
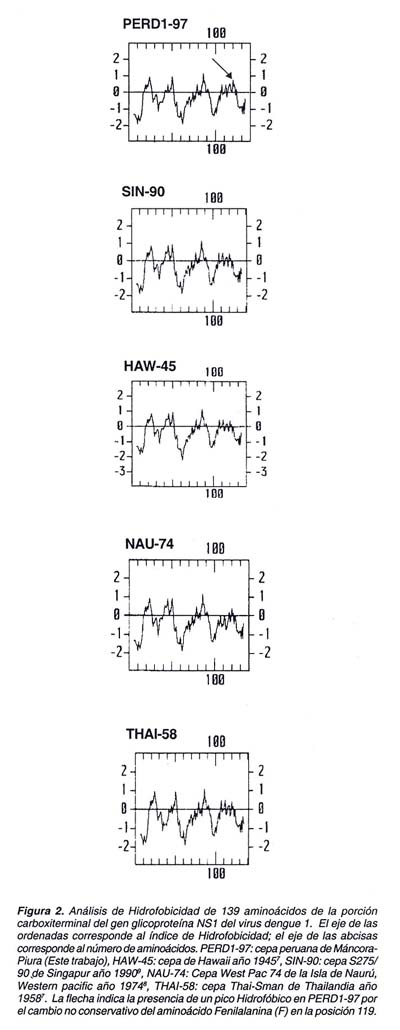
De otro lado, el número de cambios aminoacídicos de tipo no conservativos entre PERD1-97 y los demás aislamientos referenciales fue relativamente bajo. Cabe resaltar que la substitución de Serina(S) por Fenilalanina(F) (posición 119) sólo estuvo presente en el aislamiento peruano mientras que en los demás aislamientos S fue conservado (Figura 1). Adicionales comparaciones usando la misma región de NS1 entre Flavivirus (datos no mostrados en este trabajo) revelaron que S es altamente conservado con excepción del virus de la encefalitis asociado a ácaros (TBEV) donde solo hubo un cambio conservativo por adenina.
Por último, el análisis de hidrofobicidad mostró un perfil hidropático similar entre PERD1-97 y las cuatro cepas referenciales de dengue. Sin embargo un pequeño pico hidrofóbico apareció a nivel del aminoácido F de la posición 119 de PERD1-97 (Figura 2).
De manera interesante el pico hidrofóbico alterado se halló dentro de un dominio epitópico que fue reportado por Putnak 14 en otras cepas de dengue. Estos cambios en regiones o dominios importantes de NS1 han sido relacionados con procesos de evasión de la respuesta inmune y virulencia del agente infeccioso tal como se halló en el virus de la fiebre amarilla15. También se ha demostrado que algunos dominios epitópicos de NS1 son altamente homólogos a adhesinas e integrinas y que podrían generar procesos hemorrágicos6. En consecuencia la aparición de cambios no conservativos y/o la alteración de dominios epitópicos deben ser ampliamente estudiados en cepas peruanas de dengue 1, con el fin de analizar con mayor detalle algún tipo de evento molecular en NS1 que este produciendo atenuación o exacerbación de la patogenicidad del virus.
Pese a estos pequeños cambios, los perfiles de hidrofobicidad entre PERD1 -97 y las cepas de referencia fueron en general superponibles. Esta característica de conservación es propia de proteínas que cumplen importantes roles biológicos. En el caso de NS1, algunos autores proponen que la proteína podría estar implicada en procesos de replicación viral, específicamente como parte de la maquinaria de replicación del ARN 16.
En resumen a partir de los datos obtenidos en este trabajo se sugiere profundizar el estudio de las comparaciones filogenéticas entre diferentes aislamientos peruanos de dengue 1 a través del tiempo. Dichos datos ayudarían a establecer si un mismo genotipo de dengue 1 se encuentra circulando en el país o están apareciendo nuevos genotipos.
Asimismo el análisis de las variaciones genéticas en otros dominios importantes del gen que codifica NS1 debe ser profundizado conjuntamente con estudios inmunológicos que revelen la importancia de estos cambios durante la respuesta humoral frente al virus. Asimismo otros genes virales que codifican proteínas implicadas en la virulencia y procesos de respuesta inmune deberían ser analizados a fin de dilucidar la naturaleza del dengue 1 que circula en el Perú.

REFERENCIAS
1. World Health Organization. Dengue. Wkly Epidem Rec 1981; 56: 398. [ Links ]
2. Nolasco O, Carrillo C, Gutiérrez V, Yábar C, Douglas S, García M, et al. Diagnóstico temprano en un brote epidémico del virus dengue en Piura usando RT-PCR y Nested-PCR. Rev Med Exp INS 1997; 14:13-17. [ Links ]
3. García G, David WV, Del Angel R. Recognition of syntethic oligopeptides from nonstructural proteins NS1 and NS3 of Dengue-4 virus by sera from dengue virus-infected children. Am J Trop Med Hyg 1997; 56(4): 466-70. [ Links ]
4. Huang JH, Wey JJ, Sun YC, Chin C, Chien LJ, Wu YC. Antibody responses to an inmunodominant nonstructural 1 synthetic peptide in patients with dengue fever and dengue heamorrhagic fever. J. Med Virol 1999; 57(1): 1-8. [ Links ]
5. Kuno G, Vorndam AV, Gubler DJ. Study of anti-Dengue NS1 antibody by Western blot. J Med Virol 1999; 32(2): 102-8. [ Links ]
6. Falconar K. The dengue virus nonstructural-1 protein (NS1) generates antibodies to common epitopes on human blood cotting, integrin/adhesin proteins and binds to human endothelial cells: potential implications in haemorrhagic fever pathogenesis. Arch Virol 1997; 142(5): 897-916. [ Links ]
7. Shiu SY, Ip KW, Gould EA, Chang KM. Comparative analisis of dengue-1 viruses prototype Hawaii strains and Thai isolate TH-Sman, and determination of the intratypic variation of NS1 protein among Dengue-1 viruses. Arch Virol 1993; 31(34): 447-54. [ Links ]
8. Mason P, McAda P, Mason T, Fournier M. Sequence of Dengue- 1 virus genome in the region encoding the three structural proteins and the major non- structural protein NS1. Virology 1987; 161(1): 262-7. [ Links ]
9. Fu J, Tan BH, Yap EH, Chan YC, Tan YH. Full-length cDNA sequence of Dengue type 1 virus (Singapore strain S275/ 90). Virology 1992; 188(2): 953-8. [ Links ]
10. Yábar C, Carrillo C, Nolasco O, García M, Montoya Y. Diagnóstico temprano del virus dengue 1 usando RT-PCR y perspectivas para la caracterización molecular de cepas autóctonas. Rev Med Exp 1999; 15(1-2): 31-4. [ Links ]
11. Yabar C, Carrillo C, Nolasco O, García M, Montoya Y. Partial genomic Analysis of the glicoprotein of the dengue virus type 1 from Mancora - Piura. Access Number AF097020.1998. [ Links ]
12. Chungue E, Cassar O, Drouet MT, Guzman MG, Laille M, Rosen L, et al. Molecular epidemiology of Dengue-1 and Dengue-4 viruses. J Gen Virol 1995; 76(Pt.7): 1877-84. [ Links ]
13. Hanh YS, Galler R, Hunkapiller T, Dalrymple JM, Strauss JH, Strauss EG. Nucleotide sequence of Dengue 2 RNA and comparison of the encoded proteins with those of the other flaviviruses. Virology 1988; 162(1): 167-80. [ Links ]
14. Putnak JR, Charles PC, Padmanabhan R, Irie K., Hoke CH, Burke DS. Funtional and antigenic domains of the Dengue-2 virus nonstructural glycoprotein NS1. Virol 1988; 163(1): 93-103. [ Links ]
15. Muylaert I, Galler R, Rice C. Genetic Analysis of the yellow fever virus NS1 protein: identification of a temperaturesensitive mutation wich blocks RNA acumulation. J Gen Virol 1997; 71(1): 291-98. [ Links ]
16. Lindenbach BD, Rice CM. Genetic interaction of flavivirus nonstructural proteins NS1 and NS4A as a determinant of replicase function. J Virol 1999; 73(6): 4611-21. [ Links ]
Correspondencia: Carlos Yábar Varas.Av. Manuel C. de la torre 477, Los Ficus, Santa Anita, Lima, Perú.Telf.:478-0401 Email: cybar@hotmail.com

