










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.20 n.2 Lima abr./jun. 2003
TEMA DE REVISIÓN
Eventos moleculares, genéticos e inmunológicos durante la interacción VIH-Hombre.
Carlos Yábar V.1
1 División de Biología Molecular. Centro Nacional de Salud Pública. Instituto Nacional de Salud. Lima, Perú.
RESUMEN
En el presente trabajo se hace una revisión de los principales estudios realizados en el aspecto genético, molecular e inmunológico de la interacción VIH-Hombre. Del mismo modo, se citan algunos alcances actuales sobre los progresos en el tratamiento contra el SIDA. Finalmente, se plantean estrategias experimentales que podrían ser aplicadas a la realidad peruana y que permitirían responder algunos vacíos sobre los factores genéticos humanos y virales que influyen sobre la transmisión y progresión de la enfermedad.
Palabras clave: VIH; Genes; Inmunología (Fuente: BIREME)
ABSTRACT
This is a review about the major studies related to HIV-Human interaction in genetic, molecular and immunological. In the same way, some highlights of the current progress in AIDS therapy are mentioned. Finally, some experimental strategies to be applied in the Peruvian setting that might allow to answer some questions about human and viral factors implicated on transmission and progression of the disease are suggested.
Key words: HIV; Genes; Immunology (Source: BIREME)
INTRODUCCIÓN
Los mecanismos biológicos que rigen la interacción del hombre y el virus de la inmunodeficiencia humana (VIH) son muy complejos. Hasta la actualidad, se han logrado dilucidar una serie de eventos que ocurren desde la introducción del virus en el sistema biológico humano hasta la generación de millones de partículas virales como resultado final de la infección. Estos eventos están relacionados con mecanismos genéticos y moleculares que han permitido al virus variar, es decir cambiar a diferentes especies - hoy denominadas cuasi especies - y al mismo tiempo, desencadenar una maquinaria de sorprendente complejidad, que en su conjunto han permitido al virus modular a su libre albedrío el sistema inmunológico humano. Todos estos mecanismos han sido tan exitosos que han permitido al VIH ir ganando con increíble sobriedad esta guerra por la supervivencia. El hombre por su parte, también ha sabido ser un buen protagonista en este repertorio de armas biológicas. Además de los fármacos y moléculas sintéticas diseñadas como fruto de su habilidad y que han contribuido a mantener en temporal equidad su interacción biológica con el virus y con cierto éxito prolongar la vida, se encuentra el hecho de que ciertas poblaciones humanas han logrado evitar el ingreso e infección del virus al presentar un nivel de polimorfismo en genes que codifican los Antígenos del Leucocito Humano o HLA. Del mismo modo, han aparecido individuos homocigotes y heterocigotes de genes mutados que codifican receptores de moléculas quimioatrayentes y que han demostrado ser útiles en evitar el ingreso del virus en la célula hospedera. Este singular repertorio genético en el hombre conjuntamente con los antirretrovirales y otras moléculas antivirales, están desempeñando un papel muy importante en contrarrestar el inminente progreso del Síndrome de Inmunodeficiencia Humana o SIDA. A continuación se detallan los aspectos más importantes que rigen esta interacción VIH-hombre.
ANTECEDENTES Y SITUACIÓN ACTUAL DE LA ENFERMEDAD
En el período de octubre de 1980 y mayo de 1981, cinco hombres homosexuales fueron diagnosticados con neumonía por Pneumocystis carinii y sarcoma de Kaposi en tres diferentes hospitales de Los Angeles, California. Estos casos, fueron reportados como inusuales desde que se había detectado inmunodeficiencia e infección oportunista con citomegalovirus (CMV) y candidiasis bucal en pacientes aparentemente sanos. Los científicos de entonces presumían que esta rara enfermedad podría estar relacionada a comportamientos homosexuales e incluso a una probable predisposición genética 1 . Hoy en día, esta enfermedad conocida como Síndrome de Inmunodeficiencia Adquirida (SIDA) no sólo afecta a homosexuales sino a cualquier individuo que tenga contacto directo con el virus, entre los cuales se incluye a heterosexuales en general y determinados grupos de personas con conducta de riesgo que conlleve a una eventual infección por VIH como trabajadoras sexuales (TS), usuarios de drogas intravenosas (IDUs) y gestantes. Este amplio rango de conductas de riesgo relacionados con factores sociales y culturales han permitido la rápida diseminación de la enfermedad. Según los últimos informes, se afirma que aproximadamente 18,8 millones de personas alrededor del mundo han muerto de SIDA, de los cuales 3,8 millones son niños. Sin embargo, casi el doble de dicha población, unos 36 millones de personas entre adultos y niños, están siendo infectados con el VIH, los cuales morirían en los próximos diez años. De acuerdo con datos estadísticos de 1999, se calcula que un total de 5,4 millones de personas ya han sido nuevamente infectadas con el VIH 2 En el Perú los casos de SIDA han ido en aumento de manera preocupante desde que en 1983 se detectara la enfermedad. Hasta mediados del 2000 se reportaron un total de 11 694 nuevos casos de SIDA (Ver Sala de Situación de Salud. Situación de SIDA en el Perú de la Organización Panamericana de la Salud, http://www.per.ops-oms.org/sala-sit/SIDA2000.ppt). La mayoría de dichos casos han sido detectados en Lima y Callao, mientras que otros lugares del país como Piura, Ancash, La Libertad, Loreto, Ica y Arequipa muestran cifras relativamente significativas. Recientes datos publicados en el boletín epidemiológico de la Oficina General de Epidemiología mencionan que los casos de VIH hasta febrero de 2003 ascienden a 13 427, siendo la vía de transmisión más frecuente el contacto sexual con un índice de 96,0%3 . El síndrome de la inmunodeficiencia adquirida (SIDA) es una enfermedad causada por el virus de la inmunodeficiencia humana (VIH) y transmitida al hombre a través del contacto sexual, vía parenteral (de madre a hijo) y vía sanguínea (por transfusión sanguínea o contacto accidental con sangre infectada). La definición clínica de la enfermedad dada por el CDC en 1993 es compleja y se aplica con fines de vigilancia, según los cuadros clínicos y el recuento de linfocitos T CD4+. De acuerdo con ello, cualquier paciente con infección con VIH con un recuento de linfocitos T CD4+ menor de 200/uL sufre por definición el SIDA independientemente de las enfermedades oportunistas que aparezcan como la tuberculosis, neumonía y cáncer cervical 4 . La infección sin intervención terapéutica generalmente se expresa a través de tres fases durante un período de ocho a diez años5 :
-Infección aguda, que es la fase de viremia y donde se genera síndrome de tipo mononucleosis severa
- Latencia clínica, donde ocurre un decrecimiento lento pero consistente de linfocitos CD4 y de todo el sistema inmunológico integral. Asimismo, hay una baja en la concentración de partículas virales en sangre periférica y decrecimiento del título de anticuerpos antivirales
-Enfermedad clínicamente aparente, es caracterizada por una reaparición de partículas virales, una pérdida significante de linfocitos T CD4, una declinación del sistema inmunológico y numerosas enfermedades que definen en su totalidad el SIDA.
Otras anomalías muy relacionadas a la infección por VIH-1 es la manifestación de demencia por la alteración del sistema nervioso central conocido como complejo de demencia por SIDA (ADC por sus siglas en inglés) o más recientemente demencia por VIH-1 (HIVD por sus siglas en inglés)6 Debido a que los aspectos clínicos del SIDA son vastos y no corresponden al objetivo principal de este trabajo, no se dará mayor detalle al respecto. Sin embargo se tomará mayor atención sobre el agente infeccioso y los últimos alcances que se han obtenido con respecto al estudio molecular del VIH y las variantes genéticas aparecidas hasta la actualidad.
El VIRUS: UNA PARADOJA ENTRE SIMPLICIDAD Y COMPLEJIDAD
El virus de la inmunodeficiencia humana (VIH) es un retrovirus que pertenece a la familia de los lentivirus o virus lentos. Se caracteriza por presentar un genoma de ARN de cadena simple el cual requiere ser convertido a ADN complementario (ADNc) para luego ser integrado al genoma del hospedero. Dicho mecanismo es llevado a cabo por intermedio de una transcriptasa reversa del virus, la cual viene incorporada dentro de la cápside 7 . La morfología del VIH de acuerdo con las microfotografías electrónicas es icosaédrica provista de numerosas proyecciones.
Presenta una cápside, la cual está cubierta por una membrana de bicapa fosfolipídica que el virus obtiene a partir de la célula hospedera a través del mecanismo de esporulación o escape hacia el medio extracelular. En la superficie de la membrana se encuentra embebidas proteínas estructurales denominadasproteínas de la envoltura. Entre ellas figuran dos glicoproteínas de 120 y 41 kDa de peso molecular denominadas gp120 y gp41. Estas proteínas son las responsables en reconocer a los receptores CD4 de los linfocitos así como también diversos ligandos de la célula a ser infectada, que en su conjunto permitirán el ingreso e internalización del VIH 4 . (figura Nº 1).
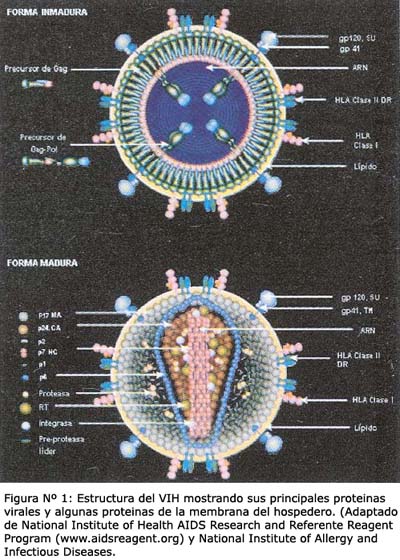
El genoma del virus comprende dos hebras de ARN de un tamaño aproximado de 9,2 y 9,7 Kilobases (Kb). Está compuesto de tres regiones genéticas: gag, que codifica proteínas estructurales del centro viral o core, env que codifica glicoproteínas de la envoltura y pol que contiene secuencias que codifican la enzima transcriptasa reversa, endonucleasa y proteasas virales necesarias para la replicación del VIH. Al mismo tiempo, estas regiones contienen de manera superpuesta seis marcos de lectura correspondiente a genes que codifican proteínas no estructurales de singular importancia para ciertas funciones biológicas que el virus requiere durante su crecimiento, ensamblaje y replicación 8 . La función de cada gen se resume en la Tabla Nº 1. A continuación se detallarán los mecanismos de replicación del virus durante su entrada en la célula hospedera.

PRIMER RECONOCIMIENTO DEL VIRUS CON LA CÉLULA HOSPEDERA: SE DESENCADENA LA REPLICACIÓN DEL VIRUS
El ciclo de replicación del VIH empieza con el primer contacto del virus con la célula hospedera, mecanismo mediado por la interacción de la proteína gp120 y el dominio VI del extremo N terminal del receptor CD4 de la célula hospedera 4 . El receptor CD4 se localiza a nivel de macrófagos, linfocitos T, células dendríticas y de Langerhans. Sin embargo esta unión no es aislada, pues existen otras moléculas de la célula denominadas correceptores que cumplen una función importante para la unión específica del virus a la célula hospedera. Estas moléculas son principalmente receptores de quimiocinas, como el CXCR4, el cual determina el tropismo del VIH-1 a células T. Del mismo modo, existe otro correceptor de tipo B-quimioquina denominado CCR5 localizado principalmente a nivel de macrófagos. Un reciente estudio de corte transversal ha descubierto que niveles altos de las quimiocinas MIP-x y MIP B pero no RANTES, obtenidas in vitro a partir de Linfocitos T CD8+ tiene una alta correlación con la ausencia de síntomas en pacientes infectados con VIH. Este evento probablemente responde a una actividad específica anti-VIH el cual cumple un papel en el control viral 9 . Mutaciones localizadas en estas moléculas son críticas para el éxito de la infección por VIH. Más adelante se explicará con mayor detalle este fenómeno. Después de la unión específica de gp120 con los receptores y correceptores de la célula hospedera, la gp41 facilita la fusión de la membrana del virus con la célula. Este fenómeno ocurre por la inserción de la cabeza amino-terminal de esta glicoproteína en la membrana celular adyacente 10 . El mecanismo de inserción de gp41 permite la introducción de la partícula viral en la célula hospedera, fenómeno que trae como consecuencia la pérdida de la envoltura viral. En subsecuentes eventos alternativos ocurre la migración de la gp120 en la membrana de la célula infectada. Este fenómeno promueve la unión de gp120 de la célula infectada con el receptor CD4 de otra célula no infectada, generando la fusión de membranas entre varias células. Este proceso observado in vitro promueve el paso del virión entre células infectadas de manera directa 7 . En el interior del virus se inicia el proceso de trascripción reversa que facilita la conversión del ARN genómico en ADNc mediante la acción de la transcriptasa reversa. Este proceso genera dos secuencias de repeticiones terminales largas o LTR (del inglés long terminal repeat) en cada extremo del ADN proviral. Una vez formado el ADN viral bicatenario, el virus introduce el material genético en el genoma de la célula hospedera mediante la acción de la enzima integrasa sintetizada por el virus mismo. Los extremos de LTR subsecuentemente participan en la integración del provirus 11 . Una vez integrado el ADN proviral, permanecerá latente por meses y hasta años, hasta que ocurra una serie de interacciones entre el virus y el hospedero que permitan su traducción y posterior ensamblaje y maduración del virus 4 . Algunas moléculas de ADNc que fueron sintetizadas incompletamente no serán integradas eficientemente y permanecerán en el citoplasma celular. Diversos autores piensan que la concentración de ADN viral no integrado probablemente tiene un efecto citopático, y que si no asesina directamente a la célula podría representar en su conjunto, otra forma latente de infección 7 .
EL LTR ES CLAVE EN LA PATOGENICIDAD DEL VIH
El ADN integrado en el genoma del hospedero sufre un proceso de trascripción el cual es regulado por los LTR. Básicamente, los LTR corresponden a secuencias específicas de ADN ubicadas a los extremos del genoma del virus (figura Nº 2), los cuales cumplen funciones reguladoras a nivel molecular, es decir incrementan o disminuyen la tasa de replicación del genoma viral y la expresión de diversos genes. En ese sentido, estas secuencias son el punto de convergencia para factores de transcripción y elementos de la maquinaria transcripcional del hospedero así como también de la proteínas codificadas por el virus que enaltecen o modulan la expresión de proteínas y ARN viral 11 . En consecuencia los LTR cumplen un papel importante en la regulación de la expresión de proteínas virales y del hospedero generando una gran orquesta de moléculas interaccionantes.
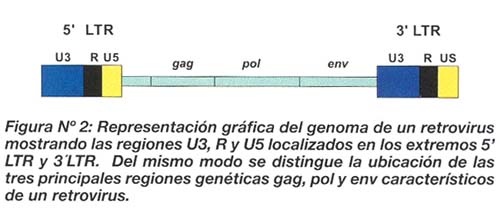
Estructuralmente, los LTR tienen una longitud de 640 pb, segmentado en tres regiones bien diferenciadas conocidas como U3, R y U5 11 . U3 es la región moduladora, y tiene sitios de unión para numerosos factores del hospedero entre los cuales se encuentran secuencias cuesta arriba o upstream de los sitios NF-kB, los factores de la proteína de unión al enhancer CCAAT (C/EBP), los factores nucleares de células T activadas (NF-AT)12 , y también secuencias reguladoras de activación en cis o secuencias cis-activantes de unión a factores de respuesta a AMP cíclico (ATF/CREB)13 . Un elemento conocido como potenciador o enhancer también se localiza dentro de U3 y contiene dos copias de sitios de unión de 10 pb para NF-kB y factores relacionados 14 . También se tiene la región central o core perteneciente a U3, la cual tiene singular importancia por la presencia de la caja TATAA para la unión de la proteína de unión a la caja TATAA (TBP) y regiones ricas en GC para la familia Sp de los factores de trascripción 15 . Como puede apreciarse, este segmento cuenta con secuencias homólogas a sitios estratégicos para factores de replicación del ADN en humanos. El segundo segmento conocido como región R del LTR presenta secuencias de respuesta transactivadora (TAR), a las que se une la proteína viral Tat. Es importante señalar que los miembros de la familia NF-kB, pueden promover la iniciación de la trascripción y la elongación de manera totalmente independientemente de Tat16. Este hecho sugiere que existen mecanismos alternativos del VIH para lograr su activación incluso en casos de que Tat no sea funcional o eficiente. Como es evidente, LTR es crítico para los procesos de activación viral más aún desde que compromete la normal fisiología de la célula infectada por la presencia de secuencias homólogas de unión a factores de trascripción del hospedero. En consecuencia es importante tomar en cuenta el papel fundamental de LTR en la patogénesis de la enfermedad. Estos procesos de patogénesis podrían ocurrir mediante los siguientes fenómenos:
-Regulación de los LTR por factores externos. Aquí los LTR cumplen un papel importante en regular la fase de latencia del virus, especialmente durante la administración de nuevos regímenes de terapia antiretroviral altamente activa (HAART)17 y por la presencia de otros agentes infecciosos oportunistas que activan genes transactivadores de LTR como el citomegalovirus 18 o disminuyen la regulación del LTR por activación del antígeno nuclear asociado a latencia en herpes virus 8 y sarcoma de Kaposi asociado a herpes virus 19 .
-Los LTR facilitan la infección de monocitos y macrófagospor VIH-1, donde ocurre una serie de interacciones moleculares entre el LTR viral y diversos factores que modulan la trascripción de estas células. Dentro de los principales se encuentran el factor C/EBP cuya estructura molecular a manera de cierre de leucinas (Leucine zipper) facilita la unión con sitios específicos del ADN necesario para la trascripción. En el LTR de VIH-1 se han caracterizado sitios específicos para la unión de este factor 20 . Adyacente a este sitio se localiza una secuencia de unión para factores activadores de la trascripción y de unión a elementos de respuesta a AMP cíclico (ATF/CREB)13 . La combinación resultante de todos estos factores que comparten casi el mismo sitio de unión en LTR sugieren la existencia de una amplia gama de mecanismos que pueden servir para regular la actividad de LTR dentro de macrófagos y monocitos.
-Los LTR ganan acceso al interior del ADN, que involucra mecanismos complejos por el cual el LTR logra ingresar a regiones del ADN del hospedero aún en presencia de nucleosomas, por remodelación de la cromatina en la vecindad de los LTR. Este proceso requiere la unión específica del LTR sobre una región flanqueada por dos nucleosomas denominados nuc-0 y nuc-1 y separados por una región libre de nucleosomas de aproximadamente 265 nucleótidos. Para llevar a cabo el inicio de la trascripción se requiere la disrupción de nuc-1, permitiendo la unión de los factores de la trascripción en la región LTR viral 21 .
-Infección del VIH-1 en células nerviosas, al respecto se ha demostrado que las primeras células sensibles a la infección corresponden a las líneas celulares de monocitos y macrófagos incluyendo células microgliales del cerebro 22 .Otras líneas también son afectadas como es el caso de los astrocitos, los cuales son susceptibles a la infección por VIH in vitro y pueden mantener en su contenido citoplasmático material genético y proteínas de VIH-1 in vivo 23 . De otro lado, algunos estudios realizados en el Sistema Nervioso Central (SNC) indican que la activación del LTR puede expresarse diferencialmente durante los distintos estados de desarrollo embrional de la célula nerviosa. Estos estudios fueron llevados a cabo usando LTR transgénico derivado del cerebro 24. Asimismo, existe una variación secuencial de LTR por tipo de compartamentalización celular, de tal manera que el LTR proveniente de sangre periférica difiere del que proviene del SNC en un mismo paciente. Estos resultados fueron demostrados por la presencia de secuencias que son reconocidas por factores del hospedero de sangre periférica distintos a los de LTR de SNC 25 .
-Papel de las proteínas virales Tat y Vpr en la infección, las cuales son importantes dado que actúan incrementando la expresión viral ya que inducen la actividad del LTR. En ese sentido, la proteína Tat se une a la estructura en stem-loop del elemento TAR consistente de 26 nucleótidos. Este proceso permite la interacción de Tat con la maquinaria transcripcional basal del hospedero 26 . Además de estos procesos Tat es capaz de modular la expresión de genes celulares como el que codifica la superóxido dismutasa dependiente de manganeso. De manera interesante Tat también puede actuar como una neurotoxina desencadenando efectos asociados a neuropatogénesis y demencia 23 . Vpr por su parte también es capaz de aumentar la actividad LTR teniendo un efecto indirecto sobre el arresto del ciclo celular 27 . Asimismo, se ha observado que proteínas de fusión de Vpr presentan interacción directa con los sitios I y II de C/ EBP, sugiriendo que Vpr puede modular la expresión de VIH-1 en células monocíticas a través de la interacción con factores C/EBP y sus sitios cis-activadores 28 .
-El polimorfismo de los LTR, que esta dado principalmente por la baja fidelidad de la transcriptasa reversa, falta de actividad correctora de lectura de la polimerasa viral (proofreading), tasas de producción viral y por la misma presión de selección in vivo. También son importantes los procesos de recombinación que ocurren entre especies de VIH que van apareciendo por cada generación durante todo el curso de la enfermedad. Son muy conocidos los procesos en los cuales existe infección dual entre diferentes subtipos de VIH en un mismo individuo que por recombinación da lugar al establecimiento de nuevas generaciones de cuasi especies recombinantes de HIV-1 29 .
Es importante señalar que la presencia de los diferentes subtipos de VIH no sólo se diferencian por las distancias genéticas producto de la divergencia en su secuencia de nucleótidos o aminoácidos, sino también por la presencia o ausencia de secuencias reguladoras de la trascripción en sus LTRs. Este es el caso del subtipo recombinante CRF01_AE producto de la recombinación de los subtipos A y E reportado en África Central, el cual contiene un sitio NF-Kb (AGGACTTCC) que difiere de la secuencia consenso GGGRNNYYCC, debido a una transición y una delesión en el extremo 5 30 . En total, hacia 1999 ya se habían reportado alrededor de catorce subtipos recombinantes dispersados por todo el planeta 31 y probablemente, la cifra esté experimentado un aumento en los últimos años. De otro lado, el subtipo C de VIH-1 también fue reportado por presentar tres sitios NF- kB más que otros subtipos. De manera interesante este subtipo demostró una mayor trascripción de la proteína p24 y una alta tasa de replicación y expansión que otros subtipos 32 . Ello sugiere que la presencia de un alto número de copias en secuencias de unión a factores de la trascripción producto de la variación del LTR podría contribuir a marcar diferencias de virulencia entre subtipos. Sin embargo, algunos estudios de cohorte han demostrado que estas diferencias no guardan ninguna correlación estadística con la progresión de la enfermedad 33 . En consecuencia la contribución de la variabilidad del LTR en la patogénesis de la enfermedad podría darse bajo ciertas circunstancias y por lo tanto deberá ser tomada con cuidado durante la interpretación de los resultados. · El LTR del virus también modula factores del sistema inmune, es decir algunos factores del sistema inmunológico del hospedero también influyen en la activación de los LTR virales para la transcripción del ADN proviral integrado. Entre estos factores tenemos a las lectinas de unión al TCR, factor de necrosis tumoral (TNF) e interluquina 6 (IL-6). La estimulación de estas citoquinas probablemente induce factores nucleares que se unen a secuencias virales del LTR que son homólogas a la secuencias de unión NFk-B. Estos procesos están relacionados con los mecanismos que promueven la activación o crecimiento de la célula T infectada y en consecuencia con la patogénesis de la enfermedad a través de dos vías 7 :
-Activación fisiológica de la célula T infectada que está en un estado de latencia, y que al finalizar da lugar a la producción de nuevas partículas virales.
-Las múltiples infecciones en pacientes con SIDA que lleva a una elevada producción de TNF y que estimula a la producción del VIH y a la infección de células adicionales.
En síntesis, los datos mencionados anteriormente sugieren que los procesos que gobiernan la interacción VIH-hombre se realizan en complicidad con los factores de la célula hospedera. Este fenómeno por tanto, es consecuencia de las estrategias moleculares del agente infeccioso de disfrazar secuencias genéticas específicas de su LTR para ser reconocidas por las proteínas celulares reguladoras, moduladoras o enaltecedoras de la trascripción. Muchas de las proteínas virales incluso pueden interactuar con proteínas celulares alterando los procesos fisiológicos normales de la célula y en algunos casos pueden desencadenar efectos tóxicos en ella. En combinación con estos fenómenos también se debe considerar la habilidad del agente infeccioso para mutar y desarrollar variantes genéticas con propiedades infectivas diferentes entre subtipos genéticos. A continuación se hará una breve revisión de los acontecimientos de variabilidad genética y la aparición de mutantes resistentes a antirretrovirales. El primer suceso ha permitido al VIH superar la barrera inmunológica del hospedero y generar un mosaico de subespecies genéticas que hasta el momento ya se han establecido como nuevas poblaciones de VIH descritas en estudios de epidemiología molecular en todo el mundo. El segundo acontecimiento ha generado especies mutantes que han demostrado habilidad para evadir exitosamente el poder terapéutico de las nuevas drogas antirretrovirales.
EL PODER DE VARIABILIDAD DEL VIH
Los subtipos de VIH: reflejo de la variabilidad genética y recombinación Existen dos tipos de VIH: VIH-1 y VIH-2. El primero de ellos el VIH-1, es el más prevalente mientras que el segundo probablemente haya tenido un origen zoonótico (transmisión de animal a humano) debido a su alta relación genética con el virus de la inmunodeficiencia del simio (SIV por sus siglas en inglés)11 .
Debido a la alta variabilidad de su genoma, el VIH ha dado origen a diferentes grupos genéticos. Así por ejemplo el VIH-1 está dividido en tres grupos: el grupo M (del inglés main o principal), el O (del inglés outlier) y el recientemente decubierto N (de new o nuevo). De todos ellos M es el más común y a excepción de los demás se divide en diez subtipos clasificados desde la A-H, J y K. El VIH-2 por su parte sólo se divide en subtipos designados por letras desde la A hasta la F 34 . En términos de prevalencia en todo el mundo, el subtipo B es el más prevalente en América, Japón, Australia, el Caribe y Europa; los subtipos A y D predominan en el Africa Subsahariana, el subtipo C se encuentra más frecuentemente en Sudafrica y la India, mientras que el subtipo E lo es en la República Central Africana. (Ver HIV variability en la página web de UNAIDS http://www.unaids.org/hivaidsinfo/faq/
variability.html#how)
Esta gran diversidad de subtipos se debe, como se mencionó anteriormente, a la baja fidelidad de la transcriptasa reversa, la cual comete errores en la incorporación de nucleótidos. Estos errores traducidos como tasa de mutación han sido calculados in vivo utilizando plásmidos reporteros, dando como resultado un tasa de 3,4 x 10 -5 mutaciones por pares de bases por ciclo de replicación 35 . Estos valores denotan una baja tasa de fidelidad de la enzima, que consecuentemente originaría nuevas variantes genéticas o también denominadas cuasi especies, es decir partículas virales con características genéticas diferentes a la anterior generación.
Como consecuencia de ello los anticuerpos generados durante una eventual respuesta inmune frente a una primera generación son incapaces de neutralizar una segunda generación del virus 36 . En el caso de aparecer este tipo de mutaciones en proteínas que son blancos para drogas las partículas virales serán genética y fenotípicamente resistentes a dichas drogas. (Ver Genotyping: response to the resistance. En: Genotyping Launch Symposium – Responding to Resistance. Geneva, 7-9. Pagina Web: http:// www.roche-hiv.com/roche_movie.html).
Es por estas razones que en la actualidad, la determinación de subtipos genéticos de VIH es importante para el entendimiento de la biología del virus y la inmunología, así como también para el desarrollo de vacunas. Esto se debe a que el estudio de subtipos genéticos constituye una herramienta poderosa para monitorizar cambios en patrones de transmisión local y global 37.
El estudio de subtipos ha sido mayormente centrado a grupos de riesgo como pacientes adictos drogas intravenosas (IDUs por sus siglas en inglés)38, 39 en mujeres gestantes 40- 42 y en homosexuales 43
Recientemente la aparición de nuevas especies recombinantes, es decir la mezcla genética entre dos o más subtipos diferentes, viene teniendo fuerte impacto en salud pública debido a las siguientes razones (NMRCD; presentación oral en la Reunión de la Red Andina de Vigilancia epidemiológica, Lima, 2002):
-Se da lugar a una gran variabilidad del virus lo cual favorece el incremento de la complejidad general de la pandemia del VIH
-En redes sexuales, donde cocirculan los subtipos de VIH-1, siempre surgen los recombinantes
-Los estudios de inmunidad de grupos genéticos-cruzados en primates infectados con SIV (Virus de la inmunodeficiencia del simio) y humanos infectados con VIH han demostrado que solamente existe protección parcial (especialmente inmunidad de tipo celular) en los diferentes subtipos
-Los recombinantes generan una baja inmunidad (no hay protección) lo cual es un problema muy grave en el desarrollo de vacunas.
En general la extrema variabilidad genética del VIH no sólo es un evento que concierne mutaciones aleatorias sino es una estrategia del virus que le permite ser capaz de evadir la respuesta inmunológica durante el transcurso de la infección en un individuo, principalmente a nivel de HLA (antígeno de leucocito humano) y el receptor del linfocito T (TCR), es decir durante la unión del péptido viral antigénico al HLA y el reconocimiento subsiguiente al receptor de células T (TCR)44 .
Los genotipos resistentes de VIH: habilidad para evadir el poder de los antirretrovirales
Los primeros casos de caracterización molecular de cepas resistentes a VIH fueron estudiados mediante el secuenciamiento del gen que codifica la transcriptasa reveresa (TR) en pacientes con fenotipo resistente a zidovudina 45 . En dicho estudio se hallaron aproximadamente cinco mutaciones puntuales relacionados con la resistencia a este antiretroviral. Posteriormente se llegaría a caracterizar genotipos con resistencia a más de una droga o VIH multidrogo resistentes.
El reporte de un caso de multidrogo resistencia demostró la existencia de mutaciones puntuales en la TR y la proteasa, generando resistencia a diferentes fármacos como zidovudina, lamivudina, saquinavir, ritonavir, indinavir, y nelfinavir 46.
Datos recientes del 2001 muestran alrededor de 607 mutaciones diferentes altamente relacionadas con resistencia a drogas distribuidas en diferentes genes del virus: 179 en proteasa, 1 en integrasa, 296 en transcriptasa reversa y 131 en el gen de la envoltura 47 . Sin embargo, estos datos sólo corresponden al subtipo B por lo que es necesario realizar más estudios de caracterización de genotipos en otros subtipos que son más prevalentes en otras áreas del mundo.
Con respecto a las drogas antiretrovirales existen alrededor de 17 drogas clasificadas en tres grupos: los inhibidores nucleosidos de la transcriptasa reversa, (NRTI), los inhibidores no nucleósidos de la transcriptasa reversa (NNRTI) y finalmente los inhibidores de la proteasa (PI). Mayores detalles sobre las principales drogas antirretrovirales pueden ser consultadas en Goals of therapy en: Guidelines for the Use of Antiretroviral Agents in HIV-Infected Adults and Adolescents. Pag. 13. On line:www.hivatis.org).
Al respecto, es importante señalar que cuando no existe tratamiento antiretroviral las especies de VIH de genotipo resistente generalmente tienen una reducida capacidad de replicación en comparación a las de tipo salvaje 48 . En ese sentido la presión de selección ejercida por la administración de antirretrovirales desencadena un cambio en la población de cuasi especies dándoles la habilidad de pasar por el cuello de botella inducida por la droga. Este proceso crea una nueva población dominante con mayor eficiencia en la capacidad de replicación en el individuo.
Una vez que se generan los mutantes resistentes algunos autores han clasificado las presencia de las mutaciones en dos tipos: el primero por la presencia de mutaciones que confieren resistencia antiretroviral propiamente dicha y que permiten al virus sobrevivir ante la presencia de la primera presión de selección. La segunda es la aparición de mutaciones compensatorias, que van a restaurar la funcionalidad de las enzimas donde ocurrieron las primeras mutaciones confiriéndole a la nueva población una mejor capacidad de replicación en el hospedero 49, 50.
En resumen, la presencia de genotipos resistentes de VIH se muestra como una evidencia clara de la capacidad del virus para sobrevivir frente a tratamientos severos de terapia antiretroviral. Como una nueva tentativa para enfrentar estos genotipos resistentes se han venido desarrollando nuevos fármacos que interfieren en la entrada del virus a la célula hospedera y que podrían actuar en dos diferentes niveles:
a)Durante la interacción de la gp120 con el receptor CD4+51 y
b)Durante la interacción con los receptores de quimiocinas CCR5 o CXCR4 y la fusión de la membrana viral y celular 52 . Recientes trabajos presentados en el 2003 Retrovirus Conference Report y compilados por Dobkin 53 , han mostrado la efectividad de nuevos fármacos frente a cepas de VIH de laboratorio altamente resistentes a antirretrovirales como es el caso de el inhibidor de proteasa RO 033- 4649 el cual fue diseñado exclusivamente para VIH con mutaciones de resistencia en los codones 48, 82, 84 y 90. De manera interesante, el desarrollo del anticuerpo monoclonal humanizado TNX-355 ha resultado ser una excelente alternativa para fortalecer la terapia frente al VIH. Este anticuerpo bloquea completamente y de manera específica el receptor CD4 a altas dosis (25 mg/kg) por un lapso de 15 a 34 días. Finalmente cabe resaltar la presentación de la molécula PA-457, un derivado del ácido betulínico, el cual evita la maduración de la proteína viral p25 a p24 generando un cambio en la formación de la típica estructura cónica del VIH 53.
Algunas mutaciones pueden tener un efecto letal para el VIH
Hemos visto anteriormente una serie de estrategias moleculares que emplea VIH a lo largo de su interacción con el hospedero para evadir o modular la respuesta inmunológica y los diferentes fármacos antirretrovirales que se vienen administrando en la actualidad. Sin embargo, y de manera paradójica, han ido apareciendo nuevas variantes de VIH con características genotípicas diferentes, de tal manera que ha dado como consecuencia la aparición de nuevas cepas con infectividad disminuida. Estas variantes que aparecieron en la naturaleza como cepas atenuadas han sido reportadas en diferentes pacientes en los cuales la enfermedad no ha llegado a progresar después de un largo tiempo (long-term non progressor). Los análisis genéticos revelaron que en algunos grupos de este tipo de pacientes el VIH tiene defectos genéticos en el LTR, principalmente hipermutaciones de G a A en regiones promotoras del LTR 54 . Otros reportes demostraron arreglos de inserciones y delesiones a lo largo de toda la región del LTR 55 . A pesar que en dichos estudios se observó alteraciones en LTR, es probable que en mutaciones localizadas en otras partes del genoma estén también jugando un papel importante en alterar la virulencia del VIH. En líneas generales, estos estudios sugieren que la ocurrencia de mutaciones en LTR o en otras partes del genoma viral no sólo desencadena una aumento en la progresión de le enfermedad, sino también genera graves disfunciones en el virus en caso de ocurrir en regiones importantes de unión a factores de trascripción y replicación.
LA VARIABILIDAD GENÉTICA DEL HOMBRE HA PERMITIDO DESARROLLAR MUTACIONES DE SENSIBILIDAD Y RESISTENCIA A LA INFECCIÓN POR VIH
Después de analizar detenidamente los diferentes mecanismos que contempla el ataque del virus hacia el hospedero, en esta parte se estudiará la respuesta frente a este ataque a través del repertorio genético e inmunológico que tiene el hombre para contrarrestar al virus, y que representa un aspecto muy importante en el área de la patogenicidad y progresión del SIDA propiamente dicho. Estos eventos en muchos de los casos suelen ser independientes uno del otro, pero en otros pueden actuar de manera armónica generando individuos humanos VIH positivos pero asintomáticos. En ese sentido se hará una revisión de las variantes genéticas del hospedero reportadas en los últimos años y se hará mención de algunos aspectos inmunológicos que también cumplen un papel importante en acelerar, aletargar y en el mejor de los casos evitar el crecimiento y desarrollo viral.
Existen individuos genéticamente susceptibles a la infección por VIH
De manera interesante, las últimas investigaciones realizadas en el campo genético e inmunológico en el hombre, han permitido determinar la existencia de poblaciones humanas que son genéticamente sensibles, más que otros, a la infección por VIH y que consecuentemente desarrollan SIDA en un tiempo más corto. Sin embargo, para poder discutir con mayor detalle sobre este tema, es preciso recordar algunos aspectos básicos relacionados con los alelos humanos implicados en conferir susceptibilidad o resistencia frente al VIH en ciertas poblaciones humanas. En general, todas las variantes alélicas humanas tienen su base en el HLA del Complejo Principal de Histocompatibilidad (MHC por sus siglas en inglés). Estos loci de HLA residen en un segmento de aproximadamente 3 500 Kb dentro del cromosoma 6p21.31 7 . Gracias al gran polimorfismo contenido en esta región del ADN del hospedero es posible distinguir cientos de diferentes serogrupos clasificados en tres clases principales de HLA: Tipo I, II y III.
HLA tipo I se divide en A, B y C y HLA tipo III se divide en seis loci: DPB1, DPA1, DQB1, DQA1, DRB1 y DRA1. En cada locus existen más de cien alelos distintos, por ejemplo el locus C de HLA I tiene 111 alelos (8 serogrupos), mientras que el B
tiene 446 alelos 56 .
Con respecto a las variantes susceptibles a la enfermedad, muchos estudios interesantes fueron realizados en madres gestantes con VIH, donde se observó que la presencia de un alelo más de HLA I en el niño aumentaba doblemente el riesgo
de transmisión del virus a través de la madre 57 .
Asimismo, se observó una alta incidencia de HLA A*2301, el cual incrementaba sustancialmente la seroconversión por VIH en un estudio de cohorte en Africa 58 . De otro lado, algunos individuos homocigotes en HLA I han demostrado una gran susceptibilidad y rápida progresión de la enfermedad, situación que no ocurrió en individuos heterocigotes 59 . Estos datos sugieren que la escasa variabilidad genética observada en individuos homocigotes es crucial en el progreso de la infección ya que no responde a un avance selectivo del hospedero frente al agente infeccioso invasor. Es importante señalar que hasta la fecha se han identificado cerca de 18 alelos distintos de HLA que tienen una fuerte correlación estadística con la susceptibilidad a la infección. Cada uno de ellos se presenta de manera indistinta en diferentes grupos de riesgo y étnico. Uno de los más frecuentes es el alelo HLA B*35, el cual ha sido fuertemente asociado a una acelerada progresión de la enfermedad en distintos grupos humanos. La forma como se presenta es siempre codominante, en estado homocigote incrementa la susceptibilidad, especialmente cuando existe una simple mutación hacia la forma HLA-B*35-Px 56, 60 . En síntesis, es importante señalar que el estudio de individuos genéticamente sensibles a la infección por VIH es importante principalmente cuando se requiere tomar decisiones en la administración de antirretrovirales donde el acceso a estos fármacos es limitado, como ocurre en países en vías de desarrollo.
Hospederos genéticamente resistentes a la infección por VIH
En este grupo encontramos a los no progresores a largo plazo (NPLP), quienes son aquellos individuos que están infectados con VIH pero no hacen la enfermedad. Estos individuos se caracterizan porque no presentan síntomas, tienen un recuento de linfocitos CD4+ dentro de los valores de referencia, no necesitan de tratamiento antirretroviral y tienen muy poca carga viral en sangre periférica 61 .
Al igual que en los individuos susceptibles, en los NPLP se han observado la presencia de una serie de alelos HLA que tiene una relación en conferir protección y resistencia a la infección por VIH. Un estudio de cohorte realizado en Gambia encontró una relación de la presencia del alelo B*35 en trabajadoras sexuales expuestas al VIH pero no infectadas 62 . De singular importancia fue la presencia de anticuerpos de reactividad cruzada contra VIH-1, en pacientes infectados con VIH-2. Los análisis genéticos revelaron una incidencia del alelo B*5801 con una enaltecida respuesta a epítopes de VIH-1 63 . Estos descubrimientos sugieren que los anticuerpos dirigidos frente a VIH-2 así como también la presencia de un alelo de resistencia B*5801 podrían desempeñar un papel importante en la protección cruzada frente al VIH-1.
Cabe señalar que las bases moleculares de estos eventos por los cuales un alelo determina resistencia o no sobre el VIH aún no están muy claros. Sin embargo es probable que estos acontecimientos encuentren una explicación en las distintas preferencias de unión entre el péptido presentado por el MHC y el receptor del linfocito T. Este reconocimiento desencadena una respuesta celular que puede ser efectiva o no por el tipo de péptido presentado.
En general existen muchos alelos que han sido implicados en la resistencia a la infección por VIH y que han resultado en seroconvesión. Algunos ejemplos de ellos son los alelos DRB1*1501, DRB1*13 que fueron observados en neonatos 64 , los alelos B*44 y B*55 encontrados en amerindios e hispanos 65 . Otros alelos por ejemplo tuvieron incidencia tanto en individuos resistentes como susceptibles como es el caso del alelo B18 56 . Es importante señalar que no sólo los alelos de resistencia van a ser los responsables de proteger al paciente contra la infección del VIH, sino que han de existir otros factores, tanto del hospedero como del virus, así como también del ambiente mismo (como la administración de fármacos y agentes infecciosos oportunistas por ejemplo) que pueden estar desempeñando un papel importante en el progreso de la enfermedad.
Mutaciones en los receptores de quimiocinas también confieren resistencia a la infección por VIH
Desde hace buen tiempo se viene tomando atención de una serie de mediadores químicos de la inflamación y de la maduración de leucocitos que también se han visto relacionados en cumplir un papel muy importante en detener el progreso de la enfermedad. Estas moléculas se conocen como quimiocinas, las que se unen a receptores localizados en el linfocito T y que también van a ser cruciales en evitar la entrada del virus en la célula hospedera. A continuación se abordarán algunos aspectos concernientes al papel de las quimiocinas y sus receptores en la infección por VIH.
Las quimiocinas son proteínas de bajo peso molecular que poseen tanto propiedades quimioatrayentes como también de citoquinas. Basadas en la posición de cuatro residuos de cisteína que forman enlace disulfuro, la quimiocinas se dividen en: CXC o α-quimiocina y CC o ß-quimiocina donde X es cualquier aminoácido 66 . Cada quimiocina puede unirse a diferentes receptores y cada receptor puede albergar diferentes quimiocinas, siendo el receptor CCR5 el principal coreceptor para el reconocimiento del VIH de la cepa que se une específicamente a CD4+ y forma sincitio 61.
CCR5 y otros correceptores de quimicionas.
Desde su primera identificación en el ámbito molecular como receptor de quimiocinas 67 , hasta el descubrimiento de una mutación en el gen denominada delta-32 presente en algunos individuos que no desarrollaban SIDA 68 , el receptor CCR5 ha sido ampliamente estudiado por su papel en la transmisión y progresión de la enfermedad. De manera específica, la mutación delta-32, una delesión de un fragmento de 32 pares de bases (pb) ha sido estudiada y considera como un marcador de polimorfismo genético que brinda información sobre los mecanismos de transmisión del VIH y de la patogénesis 69 . Anteriormente se había discutido que en muchos genotipos humanos la heterocigocidad de los alelos HLA era importante para decrecer la progresión de la enfermedad. En este caso, el riesgo de infección por VIH disminuye tanto para individuos homocigotes como heterocigotes que presentan la mutación delta32 70 . Es importante señalar que el porcentaje aproximado que representa la población de individuos heterocigotes para esta mutación es de 20% en poblaciones caucásicas sanas, mientras que los homocigotes representan el 1% de los individuos 71 . En América latina se han venido desarrollando algunos estudios epidemiológicos para investigar la incidencia de esta mutación en poblaciones generales, expuestas y no expuestas al VIH. En Brasil por ejemplo se encontró una frecuencia de 0,035 de la mutación delta32 (7% de individuos heterocigotes y 0% de homocigotes)72 . En Colombia se realizó una búsqueda en 218 individuos encontrándose una frecuencia de 3,8% en individuos expuestos seronegativos, 2,7% en la población en general y 1,7% en individuos seropositivos. De todos estos individuos sólo se encontró un homocigote para delta32 73 . Cabe resaltar que en Perú aún no se han reportado datos sobre la prevalencia de este marcador de resistencia.
Algunos receptores de quimiocinas tipo α han sido también estudiados, como es el caso de la CXCR4, la cual se presenta como el correceptor alternativo del VIH. Ello significa que en la etapa tardía de infección, el virus cambia de fenotipo y selecciona esta molécula en lugar e CCR5 para ingresar a la célula hospedera 61 . Existe un reporte en el que se logró aislar una cepa de VIH que reconocía los receptores CCR2b y CCR3, los cuales también podrían estar implicados en la infección pero con menor frecuencia 74 . Mayores estudios son requeridos para determinar correlación estadística del papel de estas quimiocinas en el progreso de la enfermedad.
Finalmente es importante señalar que las quimiocinas propiamente dichas también cumplen un papel muy importante en el progresión de la enfermedad por VIH. Al respecto se ha escubierto que niveles importantes en plasma de MIP-1α y MIP-1B producidas por células T CD8+) son determinantes en la infección por VIH. Este fenómeno es evidente por la cuantificación de esta quimiocinas en pacientes asintomáticos donde los niveles de MIP-1a y MIP-1b son mucho más altos que en pacientes VIH positivos 9 .
CONCLUSIONES
Hemos realizado una revisión general de los eventos que ocurren durante la interacción del VIH y el hombre. Es importante señalar que todos los mecanismos moleculares, genéticos e inmunológicos no se encuentran aislados sino que se relacionan estrechamente. Cada uno de esos factores en alta o baja incidencia, tanto del virus como del hospedero influenciarán de manera importante en la evolución de la enfermedad.
De otro lado hemos observado que el VIH, como agente infeccioso que interactúa con el hospedero, presenta un grado de evolución importante que le ha permitido manejar la maquinaria de replicación del hombre e incluso modular la respuesta inmune. Muchos de esos eventos responden a la alta variabilidad de sus proteínas de reconocimiento a receptores y que son el primer blanco para los anticuerpos.
Asimismo, ese alto polimorfismo representa un arma de doble filo ya que si bien muchas de las mutaciones puntuales que genera el VIH logran neutralizar exitosamente la carga inmunitaria y los efectos de los antirretrovirales, otras también pueden ocasionar verdaderos desastres en la biología del virus, principalmente a nivel del LTR donde ese encuentran secuencias homólogas de inicio de replicación y de fases de crecimiento celular.
El hombre por su parte, también ha mostrado un repertorio genético singular durante su relación con el VIH, el mismo repertorio que fue producto de un proceso evolutivo de millones de años por innumerables contactos con organismos infecciosos que hoy en la actualidad aún persisten y otros posiblemente ya desaparecieron. Todo este proceso ha permitido al hombre mantener, por selectividad natural, genes o alelos que confieren susceptibilidad o resistencia a muchos agentes etiológicos entre ellos el VIH. En ese sentido, genotipos tales como HLA-B*35, delta32, CXCR4 entre otros no son más que productos finales de un proceso evolutivo causado por múltiples interacciones entre el hombre y diferentes agentes infecciosos a través del tiempo.
De otro lado, otros factores externos que no escapan de este contexto y que pertenecen a la conducta propia del hombre también influyen de manera muy importante en los acontecimientos de infección, de virulencia y resistencia tales como la promiscuidad y las relaciones sexuales en general sin protección, la exposición frecuente con grupos de riesgo como trabajadoras sexuales, usuarios de drogas intravenosas, etc.
Todos estos elementos también favorecen a la rápida diseminación del virus y traen como consecuencia una alta tasa de mutación genética viral juntamente con las distintas respuestas inmunológicas anteriormente mencionadas.
ESTUDIOS A FUTURO
Los datos descritos en esta revisión nos permiten plantear diversos estudios que podrían ser realizados en nuestro país y que podrían contribuir a ampliar nuestro conocimiento del comportamiento del VIH en la población peruana. Por ejemplo, la construcción de un banco de datos de variantes genéticas de VIH ubicadas en distintas áreas geográficas del país, así como también la determinación del desplazamiento de nuevos subtipos en la frontera, que podrían aportar información para los proyectos de vacunación contra el VIH que se vienen ejecutando por organizaciones no gubernamentales en el Perú.
Otro estudio muy importante es la determinación de la prevalencia de la mutación delta32 en CCR5 y otros receptores de quimicionas en pacientes peruanos, dado que en la actualidad no se tienen datos publicados sobre la presencia de esta mutación en nuestra población. La caracterización de nuevos factores del hospedero de resistencia al VIH, podría ser un estudio tentativo que ayudaría a caracterizar nuevos genes implicados en la resistencia o susceptibilidad al VIH. Este estudio se llevaría a cabo mediante el análisis de expresión diferencial de ARN mensajeros de linfocitos que son infectados por VIH.
COMENTARIOS FINALES
Es importante agregar que todos los eventos moleculares que se desencadenan una vez ocurrida la infección por VIH en el hospedero susceptible hasta la aparición de los primeros síntomas clínicos de SIDA son irreversibles y seguirán su curso a través del tiempo. En ese sentido no es preciso ser adivino para presagiar que muchos individuos alrededor del mundo seguirán contrayendo la enfermedad, ya que nadie esta exento de este riesgo incluido el autor de esta nota. Tampoco es irrefutable que pese a nuestros constantes intentos de destruir el virus con todo nuestro armamento tecnológico e imaginativo, el virus literalmente siga ganando la guerra. Dejamos sí, nuestras esperanzas en la comunidad científica a fin de que logre desarrollar un tratamiento 100% efectivo contra el virus; aunque para avizorar el tiempo de dicho logro sea necesario consultar una bola de cristal. Lo que sí es importante tomar en cuenta mientras tengamos este flagelo sobre nuestras espaldas, son todos los factores culturales y sociales de nuestra población y su relación con la enfermedad, ya que ellos, a pesar de estar netamente en nuestras manos aún no se encuentran muy bien encaminados. Estos factores son el fiel reflejo de nuestra humanidad y circunscriben nuestra actitud como seres pensantes y reflexivos frente a la realidad del SIDA en el mundo. Por lo tanto, las campañas de educación sexual, el uso responsable de preservativos, el manejo y control de la prostitución Clandestina , las campañas de educación y control sanitario de personas con conductas de alto riesgo y otros aspectos como estos, deberían ser ampliamente estudiados y tomados siempre en consideración; porque es posible que ellos sean los únicos medios que podamos emplear con mayor accesibilidad contra el VIH, si es que realmente queremos ganar esta guerra sin cuartel.
REFERENCIAS
1. Durack D. Opportunistic infections and kaposis sarcoma in homosexual men. N Engl J Med 1981. 10; 305(24): 1465-7. [ Links ]
2. Pan American Health Organization. World Health Organization and Joint United Nations Programme on HIV/AIDS (PAHO/ WHO/UNAIDS). HIV and AIDS in the Americas: an epidemic with many faces. 2000. [ Links ]
3. Oficina General de Epidemiología. Situación del VIH/SIDA en el Perú. En: Boletín Epidemiológico Mensual. Ministerio de Salud. Febrero 2003. [ Links ]
4. Fauci A, Clifford L. Enfermedad por el virus de la Inmunodeficiencia humana: SIDA y procesos relacionados. En: Harrison, TR. Principios de Medicina Interna. Vol II. Madrid: Mac Graw-Hill. 1998. [ Links ]
5. Pantaleo G, Graziosi C, Fauci AS. New concepts in the immunopathogenesis of human immunodeficiency virus infection. N Engl J Med 1993; 328(5): 327-35. [ Links ]
6. MacArthur JC, Selnes OA, Glass JD, Hoover DR, Bacellar H. HIV Dementia: incidence and risk factors. En: Price RW and Perry SW eds. HIV, AIDS and Brain. Vol 72 Raven Press Ltd., 1994 New York p. 251-272. [ Links ]
7. Abbas AK, Lichyman AH, Pober JS. Congenital and Acquired Immunodeficiences . En: Cellular and Molecular Immunology. 2 nd Edition. USA : WB Saunders Company; 1991. [ Links ]
8. Korber B, Brander Ch, Haynes B, Koup R, Kuiken C, Moore J, et al. Editores. HIV Molecular Immunology. Los Alamos, New México: LANL; 2001. [ Links ]
9. Cocchi F, DeVico A, Yarchoan R, Redfield R, Cleghorn F, Blattner W et. al. Higher macrophage inflammatory protein (MIP)-1a and MIP-1b levels from CD8+ T cells are associated with asymptomatic HIV-1 infection. Proc Natl Acad Sci USA. 2000; 97(25): 13812-7. [ Links ]
10. Rosenberg ZF, Fauci AS. Immunopathogenic mechanisms of HIV infection: cytokine induction of HIV expression. Immunol Today 1990; 11(5): 176-80. [ Links ]
11. Krebs FC, Hogan TC, Quiterio S, Gartner S, Wigdahl B. (2001) Lentiviral LTR-directed Expression, Sequence Variation, and Pathogenesis. En: Kuiken C, Foley B, Hahn B, Marx P, McCutchan F, Mellors JW, Wolinsky S, Korber B., eds. HIV sequence compendium. Los Alamos, New México. LANL. 2001. p 29-70. [ Links ]
12. Shaw JP, Utz PJ, Durand DB, Toole JJ, Emmel EA, Crabtree GR. Identification of a putative regulator of early T cell activation genes. Science 1988; 241(4862): 202-5. [ Links ]
13. Krebs FC, Goodenow MM, Widahl B. Neuroglial ATF/CREB factors interact with the human immunodeficiency virus type 1 long terminal repeat. J. Neurovirol 1997; 3 Suppl 1, S28-32. [ Links ]
14. Nabel G, Baltimore D. An inducible transcription factor activates expression of human immunodeficiency virus in T cell. Nature 1987; 326(6114): 711-3. [ Links ]
15. Jones KA, Peterlin BM. Control of RNA initiation and elongation at the HIV-1 promoter. Annu Rev Biochem 1994; 63: 717-43. [ Links ]
16. West MJ, Lowe AD, Karn J. Activation of human immunodeficiency virus transcription in T cells revisited: NF-kB p65 stimulates transcriptional elongation. J Virol 2001; 75(18): 8524-37. [ Links ]
17. Wong JK, Hezareh M, Gunthard HF, Havlir DV, Ignacio CC, Spina CA, et al. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 1997; 278(5341): 1291-5. [ Links ]
18. Ghazal P, Young J, Giuletti E, De Mattei C, Garcia J, Gaynor R, et al. A discrete cis element in the human immunodeficiency virus long terminal repeat mediates synergistic trans activation by cytomegalovirus immediate-early proteins. J Virol 1991; 65(12): 6735-42. [ Links ]
19. Renne R, Barry C, Dittmer D, Compitello N, Brown PO, Ganem D. Modulation of cellular and viral gene expression by the latency-associated nuclear antigen of Kaposis sarcoma-associated herpesvirus. J Virol 2001; 75(1): 458-68. [ Links ]
20. Tesmer VM, Rajadhyaksha A, Babin J, Bina M. NF-IL6-mediated transcriptional activation of the long terminal repeat of the human immunodeficiency virus type 1. Proc Natl Acad Sci USA 1993; 90(15): 7298-302. [ Links ]
21. Van Lint C. Role of chromatin in HIV-1 transcriptional regulation. Adv Pharmacol 2000; 48: 121-160. [ Links ]
22. Tillman M, Krebs FC, Wessner R, Pomeroy SM, Goodenow MM, Wigdahl B. Neuroglial-specific factors and the regulation of retrovirus transcription. Adv Neuroimmunol 1994; 4: 305-18. [ Links ]
23. Krebs FC, Ross H, McAllister J, Wigdahl B. HIV-1-associated central nervous system dysfunction. Adv Pharmacol 2000; 49: 315-85. [ Links ]
24. Buzy JM, Lindstrom LM, Zink MC, Clements JE. HIV-1 in developing CNS: developmental differences in gene expression. Virology 1995; 210(2): 361-71. [ Links ]
25. Corboy JR, Garl PJ. HIV-1 LTR DNA sequence variation in brain-derived isolates. J Neurovirol 1997; 3(5): 331-41. [ Links ]
26. Jeang KT, Berkhout B. Kinetics of HIV-1 long terminal repeat transactivation. Use of intragenic ribozyme to assess rate-limiting steps. J Biol Chem 1992; 267(25): 17891-9. [ Links ]
27. Goh WC, Rogel ME, Kinsey CM, Michael SF, Fultz PN, Nowak MA, et al. HIV-1 Vpr increases viral expression by manipulation of the cell cycle: a mechanism for selection of Vpr in vivo. Nat Med 1998; 4, 65-71. [ Links ]
28. Hogan TH, McAllister JJ, Steimer KE, Ross HL, Wigdahl B. Vpr-mediated HIV-1 LTR trans-activation: Implications relevant to CNS infection. J Neurovirol 2000; 6, 430. [ Links ]
29. Malim MH, Emerman M. HIV-1 sequence variation: drift, shift and attenuation. Cell 2001; 104(4): 469-2. [ Links ]
30. Gao F, Robertson DL, Morrison SG, Hui H, Craig S, Decker J et. al. The heterosexual human immunodeficiency virus type 1 epidemic in Thailand is caused by an intersubtype (A/E) recombinant of African origin. J Virol 1996; 70(10): 7013- 29. [ Links ]
31. Quinones-Mateu ME, Arts EJ. Recombination in HIV-1: Update and implications. AIDS Reviews.1999; 1: 89-100. [ Links ]
32. Rodenburg CM, Li Y, Trask SA, Chen Y, Decker J, Robertson DL, et al. Near full-length clones and references sequences for subtype C isolates of HIV type 1 from the three different continents. AIDS Res Hum Retroviruses 2001; 17(2); 161-8. [ Links ]
33. Quinones-Mateu ME, Mas A, Lain de Lera T, Soriano V, Alcami J, Lenderman MM et al. LTR and tat variability of HIV-1 isolates from patients with divergent rates of disease progression. Virus Res 1998; 57(1); 11-20. [ Links ]
34. Kuiken C, Thakallapalli R, Eskild A, de Ronde A. Genetic analysis reveals epidemiologic patterns in the spread of human immunodeficiency virus. Am J Epidemiol 2000; 152(9): 814-22. [ Links ]
35. Mansky LM, Temin HM. Lower in vivo mutation rate of human immunodeficiency virus type 1 than that predicted from the fidelity of purified reverse transcriptase. J Virol; 1995 69(8): 5087-94. [ Links ]
36. Baltimore D, Heilman C. Defeating AIDS: What will it take. Scientific American. 1998; pp 62-87. [ Links ]
37. Alaeus A, Significance of HIV-1 genetic subtypes. Scand J Infect Dis 2000; 32(5): 455-63. [ Links ]
38. Bobkov A, Kazennova E, Khanina T, Bobkova M, Selimova L, Kravchenko A, et al. An HIV type 1 subtype A strain of low genetic diversity continues to spread among injecting drug users in Russia: study of the new local outbreaks in Moscow and Irkuttsk. AIDS Res Hum Retroviruses 2001; 17(3):257-61. [ Links ]
39. Rossini MA, Diaz RS, Caseiro M, Turcato G, Accetturi CA, Sabino EC. HIV- 1 subtypes among intravenous drug users from two neighboring cities in Sao Paulo State, Brazil. Braz J Med Biol Res 2001; 34(1):45-7 [ Links ]
40. Gyuris AA, Segesdi J, Mezei M, Balog K, Jelenik Z, Takacs M, et al. Virological, Neurological and Histological Investigations of a Child Born to a Mother with AIDS. Pathol Oncol Res 1997; 3(4):303-8 [ Links ]
41. Dickover RE, Garratty EM, Plaeger S, Bryson YJ. Perinatal transmission of major, minor, and multiple maternal human immunodeficiency virus type 1 variants in utero and intrapartum. J Virol 2001; 75(5):2194-203 [ Links ]
42. Guevara H, Johnston E, Zijenah L, Tobaiwa O, Mason P, Contag C, et al. Prenatal transmission of subtype C HIV-1 in Zimbabwe: HIV-1 RNA and DNA in maternal and cord blood. J Acquir Immune Defic Syndr 2000; 25(5):390-7 [ Links ]
43. Morgado MG, Guimaraes ML, Neves Junior I, dos Santos VG, Linhares- de-Carvalho MI, Castello-Branco LR, et al. Molecular epidemiology of HIV in Brazil: polymorphism of the antigenically distinct HIV-1 B subtype strains. The Hospital Evandro Chagas AIDS Clinical Research Group. Mem Inst Oswaldo Cruz.1998;93(3): 383-6. [ Links ]
44. Callahan KM, Fort MM, Obah EA, Reinherz EL, Siliciano RF.Genetic variability in HIV-1 gp120 affects interactions with HLA molecules abd T Cell receptor. J Immunol 1990; 144(9): 3341-6 [ Links ]
45. Larder BA, Kemp SD. Multiple mutations in HIV-1 reverse transcriptase confer high-level resistance to zidovudine (AZT). Science 1989, 246(4934):1155-8 [ Links ]
46. Cohen OJ, Fauci AS. Transmission of multidrug-resistant human immunodeficiency virusthe wake-up call. N Engl J Med 1998; 339(5):341-3. [ Links ]
47. Parikh U, Calef C, Larder B, Schinazi R, Mellors J. Mutations in Retroviral Genes Associated with Drug Resistance. En: Kuiken C, Foley B, Hahn B, Marx P, McCutchan F, Mellors JW, Wolinsky S, Korber B., eds. HIV sequence compendium. Los Alamos, New Mexico 2001; p 29-70. [ Links ]
48. Coffin JM. HIV population dynamics in vivo: implications for genetic variation, pathogenesis, and therapy. Science.1995; 267(5197):483-9. [ Links ]
49. Berkhout B. HIV-1 evolution under pressure of protease inhibitors:climbing the stairs of viral fitness. J Biomed Sci 1999; 6(5):298-305. [ Links ]
50. Clavel F, Race E, Mammano F. HIV drug resistance and viral fitness. Adv Pharmacol 2000; 49:41–66. [ Links ]
51. Jacobson JM, Lowy I, Fletcher CV, ONeill TJ, Tran DN, Ketas TJ. et al. Single-dose safety, pharmacology, and antiviral activity of the human immunodeficiency virus (HIV) type 1 entry inhibitor PRO 542 in HIV-infected adults. J Infect Dis. 2000; 182(1):326-9. [ Links ]
52. Kilby JM, Hopkins S, Venetta TM, DiMassimo B, Cloud GA, Lee JY, et al. Potent suppression of HIV-1 replication in humans by T-20, a peptide inhibitor of gp41-mediated virus entry. Nat Med 1998; 4(11):1302–7. [ Links ]
53. Dobkin JF. 2003 Retrovirus Conference Report. Infect Med 2003; 20(3): 132- 41. [ Links ]
54. Zhang L, Huang Y, Yuan H, Chen BK, Ip J, Ho DD. Genotypic and phenotypic characterization of long terminal repeat sequences from long-term survivors of human immunodeficiency virus type 1 infection. J Virol 1997; 71(7): 5608-13 [ Links ]
55. Rousseau C, Abrams E, Lee M, Urbano R, King MC. Long terminal repeat and Nef gen variants of human immunodeficiency type 1 in perinatally infected long-term survivors and rapid progressors. AIDS Res Hum Retroviruses 1997; 13(18): 1611-23 [ Links ]
56. Trashtemberg EA, Herlich AE. A review of the role of the human leukocyte antigen (HLA) system as a host immunogenetic factor influencing HIV transmission and Progression to AIDS. En: Korber B, Brander Ch, Haynes B, Koup R, Kuiken C, Moore J, Walker B, Watknis D Eds. HIV Molecular Immunology. Los Alamos, New Mexico. USA. 2001 [ Links ]
57. MacDonald S, Embree J, Njenga S, Nagelkerke JD, Ngatia I, Mohamed A, et al. Mother-child class I HLA concordance increases perinatal human immunodeficiency virus type 1 transmission. J Infectious Diseases 1998; 177(3): 551-6 [ Links ]
58. MacDonald KS, Fowke KR, Kimani J, Dunand VA, Nagelkerke NJ, Ball TB, et al. Influence of HLA supertypes on susceptibility and resistance to human immunodeficiency virus type 1 infection. J Infect Disease 2000 181(5): 1581-9 [ Links ]
59. Carrington M, Nelson GW, Martin MP, Kissner T, Vlahov D, Goedert JJ, et al. HLA and HIV-1: Heterozygote advantage and B*35-Cw*04 disadvantage. Science 1999; 283(5408): 1748-52 [ Links ]
60. Gao X, Nelson GW, Karacki P, Martin MP, Phair J, Kaslow R, et al. Effect of a single amino acid change in MHC class I molecules on the rate of progression to AIDS. N Engl J Med. 2001. 344(22):1668-75. [ Links ]
61. Zúñiga J, Vargas-Alarcón G, Salgado N, Flores C, Martínez-Tripp S, Miranda A, et al. Los factores genéticos determinantes de la resistencia a la infección por VIH y del control de la progresión al SIDA: Implicaciones sobre la patogénesis y las estrategias terapéuticas para la erradicación del VIH. Una Revisión. Rev Invest Clin 2000. 52(3): 284-95. [ Links ]
62. Rowland-Jones S, Sutton J, Ariyoshi Y, Dong T, Gotch F, MacAdam S, et al. HIV-specific cytotoxic T-cell in HIV-exposed but uninfected Gambian women. Nat. Med 1995; 1(1): 59-64 [ Links ]
63. Bertoletti A, Cham F, McAdam S, Rostron T, Rowland-Jones S, Sabally S, et al. Cytotoxic T cells from human immunodeficiency virus type 2-infeceted patients frequently cross-react with different human immunodeficiency virus type 1 clades. J Virol 1998; 72(3): 2439-48. [ Links ]
64. Winchester R, Chen Y, Rose S, Selby J, Borkowsky W. Major hiscompatibility complex class II DR alleles DRB1*1501 and those encoding HLA- DR13 are preferentially associated with a diminution in maternally transmitted HIV- 1 infection in different ethnic groups: determination by an automated sequence- based typing method. Proc Natl Acad Sci USA 1995; 92(26): 12374-8. [ Links ]
65. De Sorrentino AH, Marinic K, Motta P, Sorrentino A, Lopez R, Illiovich E. HLA class I alleles associated with susceptibility or resistance to HIV-1 infection among a population in Chaco Province, Argentine. J Infect Dis 2000; 182(5):1523- 6. [ Links ]
66. Murphy PM. The molecular biology of leukocyte chemoattractant receptors. Annu Rev Immunol 1994; 12: 593-633 [ Links ]
67. Raport CJ, Gosling J, Schweickart VL, Gray PW, Charo IF. Molecular cloning and functional characterization of a novel human CC chemokine receptor (CCR5) for RANTES, MIP-1beta, and MIP-1alpha. J Biol Chem. 1996; 271(29):17161-6. [ Links ]
68. Huang Y, Paxton WA, Wolinsky SM, Neumann AU, Zhang L, He T, et al. The role of a mutant CCR5 allele in HIV-1 transmission and disease progression. Nat Med 1996; 2(11):1240-3. [ Links ]
69. Paxton WA, Kang S, Koup RA. The HIV type 1 coreceptor CCR5 and its role in viral transmission and disease progression. AIDS Res Hum Retroviruses 1998; 14 Suppl 1:S89-92. [ Links ]
70. Hoffman TL, MacGregor RR, Burger H, Mick R, Doms RW, Collman RG. CCR5 genotypes in sexually active couples discordant for human immunodeficiency virus type 1 infection status. J Infect Dis 1997; 176(4):1093-6 [ Links ]
71. Martinson JJ, Chapman NH, Rees DC, Liu YT, Cleg JB. Global distribution of the CCR5 gene 32-basepair deletion. Nat Genet 1997; 16(1): 100-3. [ Links ]
72. Passos GA Jr, Picanco VP. Frequency of the delta CCR5 deletion allele in the urban Brazilian population. Immunol Lett 1998; 61(2-3): 205-7 [ Links ]
73. Díaz FJ, Vega JA, Patiño PJ, Bedoya G, Nagles J, Villegas C, et al. Frequency of CCR5 delta-32 mutation in human immunodeficiency virus (HIV)- seropositive and HIV-exposed seronegative individuals and in General Population of Medellin, Colombia. Mem Inst Oswaldo Cruz. 2000; 95(2): 237-42. [ Links ]
74. Doranz BJ, Rucker J, Yi Y, Smyth RJ, Samson M, Peiper SC, et al. A dual- tropic primary HIV-1 isolate that uses fusin and the b chemokine receptor CKR-5, CKR-3 and CKR-2b as fusion cofactors. Cell 1996; 85: 1149-1158 [ Links ]
Correspondencia: Carlos Augusto Yábar Varas. División de Biología
Molecular. Centro Nacional de Salud Pública.
Dirección: Av. Cápac Yupanqui 1400. Lima 11, Perú.
Telf: 4719920. Anexo: 149 Fax: 4719920
Correo electrónico: cyabar@ins.gob.pe-bioyabar@yahoo.es

