










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica v.20 n.4 Lima oct./dic. 2003
TRABAJOS ORIGINALES
Evaluación serológica de una proteína recombinante a partir de una cepa aislada del virus de la fiebre amarilla en el Perú: un estudio piloto
Carlos Yábar V 1; Juana Choque P 1; Ysabel Montoya P 2.
1 División de Biología Molecular. Centro Nacional de Salud Pública. Instituto Nacional de Salud. Lima, Perú.
2 Instituto Peruano de Energía Nuclear. Lima, Perú.
RESUMEN
Objetivo: Evaluar serológicamente una proteína recombinante de 66 kDa (Er66) de una cepa del virus de la fiebre amarilla aislada en el Perú usando anticuerpos inmunoreactivos. Material y métodos: La proteína Er66 fue expresada in vitro en la bacteria Escherichia coli y enfrentada a títulos de 1/100, 1/50 y 1/25 de anticuerpos monoclonales, y a sueros con anticuerpos IgM e IgG inmunoreactivos contra el virus de la fiebre amarilla mediante ensayos de Western blot (WB). Asimismo, se evaluaron proteínas totales de las cepas de referencia del virus dengue (DEN) y fiebre amarilla como controles de antigenicidad. Resultados: La proteína recombinante Er66 presentó antigenicidad contra títulos de 1/50 y 1/25 de anticuerpo monoclonal (MAB8701); sin embargo, no se observó inmunoreactividad en sueros positivos para fiebre amarilla y dengue. Los ensayos con extractos crudos de la cepa de fiebre amarilla Asibi 17D (PTFA) revelaron que la antigenicidad de las proteínas virales comprendidas entre 60 y 80 kDa fue afectada negativamente a temperaturas desnaturalizantes de 100°C. En cambio, tratamientos con ß-mercaptoetanol sin calor generó un aumento de la antigenicidad de dichas proteínas. Siguiendo este mismo principio, la proteína Er66 fue sometida a tratamientos desnaturalizantes con ß-mercaptoetanol sin incluir calor y fue enfrentada nuevamente a los sueros reactivos. El resultado final reveló que la proteína Er66 generó inmunoreactividad frente a sueros con altos títulos de anticuerpos IgM e IgG para fiebre amarilla mientras que en otras muestras se observó una débil señal. Conclusiones: Los datos obtenidos en este trabajo demuestran que la Er66 pudo ser reconocida por anticuerpos IgM e IgG específicos para fiebre amarilla presentes en sueros de pacientes infectados. Los datos presentados en este artículo sugieren el uso de proteínas recombinantes como posibles candidatas de valor diagnóstico para la FA.
Palabras clave: Virus de la fiebre amarilla; Serología; Proteínas Recombinantes; Anticuerpos Monoclonales; Perú (fuente: BIREME)
ABSTRACT
Objective: To assess a 66kDa (Er66) recombinant protein of yellow fever (YF) virus using immunoreactive antibodies. Material and methods: Er66 was expressed in Escherichia coli and then it was challenged against 1/100, 1/50 and 1/25 monoclonal antibodies containing dilutions, and against clinical sera containing specific IgM and IgG antibodies against YF virus using western blot assay. Likewise, total protein content from yellow fever virus and Dengue (DEN) virus were also tested as antigenicity control. Results: Er66 recombinant protein showed antigenicity against 1/50 and 1/25 titers of monoclonal antibodies (MAB8701); however, no immunoreactivity was observed in sera positive for yellow fever and dengue infections. Serological assays with crude extracts of 17D Asibi yellow fever virus strain revealed that antigenicity of 60 – 80 kDa viral proteins was abrogated at 100 °C (denaturizing temperature). In contrast, when total protein was subjected to b-mercaptoethanol treatments without heat, the antigenicity increased. Using this principle, ER66 protein was reduced under b-mercaptoethanol treatment and it was newly challenged to reactive sera. The final result revealed that Er66 generated immunoreactivity in sera with high titers of IgM and IgG antibodies for yellow fever, while other samples showed a weak immunoreactivity signal. Conclusion: This study shows that Er66 was recognized by specific IgM and IgG yellow fever antibodies from clinical samples of infected patients. The data presented in this study suggest the use of recombinant proteins as possible candidates with a diagnostic value for yellow fever.
Key words: Yellow fever virus; Serology; Recombinant Proteins;Ê Monoclonal Antibodies; Peru (source: BIREME)
INTRODUCCIÓN
La fiebre amarilla (FA) es una enfermedad febril hemorrágica de origen viral transmitida al hombre a través de la picadura de mosquitos vectores de los géneros Aedes, Sabethes o Haemagogus1.
El agente causal es un virus de ARN que presenta un genoma de aproximadamente 11 Kb, el cual codifica 10 proteínas diferentes2. Las tres primeras proteínas conocidas como C, M y E son de tipo estructural y forman parte de la envoltura y la cápside del virus. Las siete últimas, son de tipo no estructural y están involucradas en procesos de ensamblaje, maduración y replicación del virus 3, 4,5.
Con respecto a la proteína de la envoltura (E) del virus se han reportado distintos pesos moleculares que bordean entre 49 KDa 6, 7, 54 KDa 8, 9 y 65 KDa10. Estos tamaños podrían deberse al número variable de glicosilaciones entre aislamientos de diferentes regiones, y al grado de maduración de la proteína durante el ensamblaje del virus8.
De otro lado, es importante señalar que la proteína E es la primera proteína viral reconocida por los anticuerpos del hospedero durante la primera etapa de la infección viral 11 .
Así, diversos trabajos han demostrado que esta proteína tiene características antigénicas e inmunogénicas muy importantes 6 . Estas propiedades guardan relación con la presencia de los tres dominios epitópicos A, B y C que fueron encontrados en diferentes flavivirus 12 . Adicionalmente, dos epítopes discretos fueron localizados en el dominio I (posiciones 155 y 158) de la proteína E de la cepa 17D 13 .
Si bien es cierto que las características antigénicas de la proteína E anteriormente mencionadas corresponden netamemente a la proteína nativa, se ha visto que la forma recombinante de la proteína expresada mediante ingeniería genética, conserva la misma naturaleza antigénica e inmunogénica 14 . De hecho, algunos autores han realizado la expresión de esta proteína en combinación con otras proteínas del virus con el fin de realizar ensayos de protección inmunológica en animales de experimentación. Dichos estudios se llevaron a cabo en sistemas de vaccinia y baculovirus demostrando que los niveles de protección generados principalmente por la proteína E recombinante fueron muy similares a los obtenidos con el virus 17D atenuado 15,16. Estas mismas propiedades fueron detectadas a partir de proteínas recombinantes no estructurales de la cepa 17D 17, 18 .
La antigenicidad de la proteína E recombinante de diferentes flavivirus también fue evaluada frente a sueros de pacientes mediante ensayos serológicos. Al respecto se demostró que la proteína truncada de la envoltura (E) del serotipo dengue 4 carente de la porción carboxiterminal, había presentado mayor especificidad para inducir reactividad frente a sueros de pacientes con dengue que con pacientes con encefalitis japonesa 19 . Otro trabajo demostró que antígenos recombinantes de prM y E del virus de la encefalitis japonesa habían generado inmunoreactividad por anticuerpos de pacientes voluntarios en ensayos de ELISA de captura de IgM (MAC-ELISA), de manera tal, que estos antígenos podían reemplazar a las proteínas nativas normalmente usadas en este tipo de ensayo serológico 20 . Posteriormente, se encontró un dominio B de la E a partir del serotipo dengue 2, el cual había sido fuertemente reconocido por anticuerpos de sueros de pacientes peruanos e indonesios 21.
Se debe considerar que actualmente se encuentra disponible un reactivo de diagnóstico para dengue basado en ensayos de inmunocromatografía. Esta prueba usa la porción N-terminal de la proteína recombinante de la envoltura de los cuatro serotipos de dengue y presenta una sensibilidad y especificidad de 90% y 86%, respectivamente22.
En síntesis, estos trabajos demuestran que la proteína recombinante E de dengue y encefalitis japonesa presenta características antigénicas con valor diagnóstico serológico específico para dichas enfermedades.
Es importante señalar que para el caso del virus de la fiebre amarilla, no se han reportado trabajos similares en los cuales se haya evaluado las propiedades antigénicas de la proteína recombinante de la envoltura como potencial herramienta de diagnóstico. Asimismo, no existen informes en los que se haya expresado y evaluado serológicamente alguna proteína recombinante a partir de aislamientos virales realizados en el Perú.
En ese sentido, el objetivo de este trabajo es realizar la evaluación serológica de una proteína recombinante denominada Er66 utilizando un panel de sueros inmunológicamente reactivos a fiebre amarilla. Este trabajo es el primer reporte en el que se evalúa una proteína recombinante expresada in vitro a partir de aislamientos de cepas del virus de la fiebre amarilla realizados en el Perú.
MATERIAL Y MÉTODOS
PROTEÍNA RECOMBINANTE
El presente trabajo es un estudio piloto, de diseño descriptivo en el que se evaluó una proteína recombinante correspondiente al antígeno de la envoltura (Er66) del virus de la fiebre amarilla proveniente del departamento de San Martín en el año 1999. De acuerdo con los análisis previos realizados en la secuencia de aminoácidos, la proteína Er66 correspondió a la porción carboxiterminal de la proteína de la envoltura abarcando aproximadamente 337 aminoácidos 23 . La cepa viral de la cual se sintetizó la proteína fue proporcionada por la División de Virología, Centro Nacional de Salud Pública del Instituto Nacional de Salud.
ANTÍGENOS TOTALES
Se evaluaron proteínas totales de la cepa Asibi 17D del virus de la fiebre amarilla. También fueron incluidos en el estudio, lisados de cultivo viral de la cepa viral aislada en Ayacucho en el año 1978. Por otro lado, proteínas totales del serotipo dengue 1 de referencia (cepa Singapur) y serotipo dengue 1 aislado en Tumbes fueron evaluados como controles de especificidad.
ANTICUERPOS MONOCLONALES Y FLUÍDO ASCÍTICO DE RATÓN HIPERINMUNIZADO
El fluído ascítico de ratón hiperinmunizado (FARH) y dos anticuerpos monoclonales: MAB984 (Chemicon) y 2D12, proporcionado por el Naval Medical Research Institute Detachment (NAMRID), fueron usados para la determinación de la antigenicidad de las proteínas totales del virus de la cepa vacunal Asibi 17D y Er66. Asimismo, se usó como control el anticuerpo monoclonal MAB8701 (Chemichon) específico para el serotipo dengue 1.
SUEROS CONTROLES
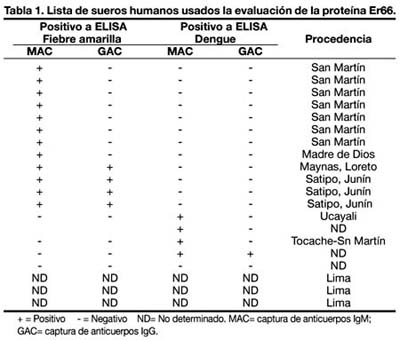
Un panel de veinte (n = 20) muestras serológicas humanas fueron empleados para la evaluación de la proteína Er66 (Tabla 1B). Ocho (n=8) de los sueros correspondieron a pacientes con altos títulos de anticuerpos IgM contra fiebre amarilla según ensayos previos de MAC-ELISA, mientras que cuatro (n=4) tuvieron títulos altos tanto para IgM e IgG. Es importante resaltar que una de las muestras positivas perteneció a un paciente con antecedente de vacunación antiamarílica con la cepa 17D. Los sueros controles de especificidad correspondieron a muestras con títulos altos de anticuerpos IgM (n=3) e IgG (n= 1) para dengue.
Del mismo modo, se consideraron dos muestras serológicas (n=2) de pacientes inmunizados con la cepa 17D por un período menor de 5 años después de haber recibido la vacuna con el fin de evaluar su inmunoreactividad frente a la proteína Er66.
Por último, se incluyeron dos muestras serológicas de individuos sanos no inmunizados contra la fiebre amarilla como controles negativos. Una de ellas fue una muestra previamente evaluada por MAC y GAG ELISA como negativo contra la FA y la otra correspondió a un individuo voluntario que nunca había recibido vacuna y tampoco había tenido contacto con el virus o viajado a zonas endémicas (Tabla 1 y 2).

AMPLIFICACIÓN, CLONACIÓN, ANÁLISIS DE SECUENCIA Y EXPRESIÓN DE ER66
Todos los procedimientos para la extracción del ARN, RT-PCR, clonación del gen que codifica la proteína E y expresión de la proteína recombinante Er66 han sido descritos previamente 23 . Brevemente, se amplificó el gen que codifica la Er66 por RT-PCR usando los primers YF7 y YF2 24 . El producto denominado FA99 fue clonado en el plásmido pGEM - T easy dando lugar a pGEM-FA99, el cual fue usado para determinar la secuencia de nucleótidos y aminoácidos del inserto de interés. A través de este análisis se pudo identificar secuencias específicas que incluían epítopes importantes de reconocimiento humoral previamente reportados en fiebre amarilla y otros flavivirus 12,13,21,25 . Dichos datos despertaron nuestro interés de expresar Er66 y evaluarla serológicamente. En ese sentido, pGEM-FA99 fue usado como ADN molde para la reamplificación de FA99 usando primers con sitios de corte para las enzimas de restricción Bgl II y Hind III. El producto fue purificado, digerido con Bgl II y Hind III y subclonado en el vector de expresión pQE40 previamente digerido con HindIII y Bgl II. El vector recombinante denominado pQE40- FA99 fue introducido en la cepa Escherichia coli XL1-Blue por electroporación induciendo la expresión de la proteína con IPTG. Posteriormente, la proteína fue visualizada por electroforesis en SDS-PAGE.
PREPARACIÓN A MEDIANA ESCALA DE LA PROTEÍNA RECOMBINANTE ER66
Para la preparación a mediana escala de la proteína Er66, se realizó un inóculo a partir del clon bacteriano XL1FA conteniendo el plásmido pQE40-FA99 en 50 mL de caldo Lurian Bertani. El cultivo fue mantenido en movimiento constante en baño maría a 37° C por toda la noche. Posteriormente, se añadió una concentración de 1mM de isopropilgalactopiranocida al cultivo para inducir la expresión de Er66 por 3 horas en movimiento de rotación constante. Las bacterias fueron precipitadas por centrifugación a 4 400 rpm durante 15 minutos. Seguidamente, 1,5 mL de buffer de lisis A (6M de Guanidina hidroclorhidra, 0,1 M de NaH 2 PO 4 , 0,01 M de Trizma-HCl, pH 8,0) permitieron la homogeneización del precipitado y subsecuente liberación del contenido celular durante 10 minutos. El lisado fue sometido a centrifugación a 4 400 rpm durante 15 minutos a temperatura ambiente y posteriormente, se recuperó el sobrenadante en viales de 1,5 mL. Se añadió un volumen de 250 mL de Ni-NTA slurry conteniendo perlas de níquel en complejo con agarosa 26 a todo el volumen del sobrenadante y se homogeneizó la mezcla mediante movimiento rotario constante en un girador orbital a temperatura ambiente por espacio de 1 hora. La agarosa fue centrifugada a 14 000 rpm durante 1 minuto y se eliminó el sobrenadante. Seguidamente, se lavó la muestra tres veces con 1 mL de buffer de lavado C (8M de urea, 0,1 M NaH 2 PO 4 , 0,01 M de Trizma-HCl, pH 6,3) y se recuperó la proteína ocho veces mediante el uso del solución amortiguadora de elusión E (8M de urea, 0,1 M NaH 2 PO 4 , 0,01 M de Trizma-HCl, pH 4,5) Se añadió glicerol a la solución conteniendo las proteínas (concentración final de glicerol 10%) y finalmente se almacenó la proteína eluída a -20° C para su uso posterior.
WESTERN BLOT
Las proteínas fueron sometidas a electroforesis de SDS –PAGE al 10% en geles de poliacrilamida de 10 y 15 pocillos y en pocillo preparativo. Posteriormente, fueron transferidas a membranas de nitrocelulosa (0,05 m de porosidad, GIBCO, BRL) usando una máquina de transferencia (BIORAD) en un sistema semiseco por 2 horas a un voltaje dependiente del área del gel (5,5 mV/cm 2 ).
Las membranas conteniendo las proteínas de interés fueron incubadas en albúmina sérica bovina (BSA) al 3% en TBS (25 mM Tris, 137 mM NaCl y 2,7 mM KCl) por 1 hora. Seguidamente, fueron lavadas tres veces con TBS conteniendo 0,05% de tween-20 (SIGMA), luego, fueron enfrentadas a anticuerpos inmunoreactivos diluidos en TBS a diferentes concentraciones. La reacción fue incubada por espacio de 3 horas o por toda la noche. El TBS fue reemplazado con buffer PBS (137 mM NaCl, 2,7 mM KCl, 0,024% de KH 2 PO 4 y 0,14% de Na 2 HPO 4 ) en el caso de realizar la incubación con el anticuerpo secundario conjugado con peroxidasa.
Culminada la incubación, se añadió anticuerpo secundario anti-IgG humano conjugado con fosfatasa alcalina (dilución 1/1000) o anticuerpo secundario anti-IgM humano o anti-IgM de ratón, conjugado con peroxidasa (dilución 1/250). La incubación en todos los casos se realizó con agitación suave en movimiento rotatorio constante usando un girador orbital durante un período de una hora. Para el revelado de la reacción antígeno - anticuerpo en sistema fosfatasa, las membranas se incubaron en el buffer para la fosfatasa alcalina o AP (100 mM NaCl, MgCl 2 , 100 mM Tris pH = 9,5) con 0,03% de Nitro Blue Tetrazolium, 0,015% de 5-Bromo 4-cloro 3-indolil fosfato y 0,025% de NN-dimetilformamida. En el sistema de peroxidasa se usó una solución de solución amortiguadora PBS conteniendo 0,04% de cloro-1-naftol, 10% de metanol y 0,025% de peróxido de hidrógeno.
DETERMINACIÓN DE LA ANTIGENICIDAD DE LA PROTEÍNA TOTAL DEL VIRUS 17D (PTFA) FRENTE A DIFERENTES TRATAMIENTOS DENATURANTES
Las proteínas totales de la cepa Asibi 17D (PTFA) fueron sometidas a diferentes agentes denaturantes como urea, mercaptoetanol y calor con el fin de dilucidar si la antigenicidad de la proteína nativa de la envoltura (E) del virus de la fiebre amarilla podría sufrir alguna alteración con estos tratamientos. Para detectar la proteína de la envoltura E en el inmunoblot como una banda comprendida entre 49 y 65 kDa, se utilizó fluido ascítico de ratón hiperinmunizado (FARH).
En ese sentido se utilizó una concentración de 5ug de extracto crudo de proteína total la cual fue sometida a tratamientos con calor a 100 °C, mercaptoetanol y urea 3M. Del mismo modo, se realizaron diferentes combinaciones entre los tres tratamientos. La denaturación por calor se realizó en baño maría a 100° C por 5 minutos. Para la denaturación con urea, PTFA fue mezclada con buffer de elusión E (descrito anteriormente) para un concentración final de 3M de urea. Para los procesos de denaturación con b-mercaptoetanol, las proteínas fueron mezcladas con buffer de la muestra (50 mM Tris-HCl ph 6,8, 2% SDS, 0,1% de azul de bromofenol, 10% de glicerol) conteniendo 1% de b-mercaptoetanol. Posteriormente, se realizó el Western blot de acuerdo con las condiciones anteriormente descritas usando FARH para la detección antígeno-anticuerpo.
DIÁLISIS DE LA PROTEÍNA ER66
Con el fin de eliminar el exceso de urea contenida en la solución de resuspensión de la proteína Er66 y conseguir la renaturación de la estructura nativa de la proteína recombinante, la diálisis se realizó usando membranas con porosidad máxima de 200 kDa (SIGMA). En tal sentido, 1 mL de buffer E conteniendo la proteína Er66 fue colocado en una pequeña bolsa de diálisis y la abertura fue sellada con ganchos a presión. La bolsa fue sumergida en 1 Lt de buffer de diálisis (25 mM Tris, 137 mM NaCl y 2,7 mM KCl) y sometida a movimiento de rotación constante usando un agitador magnético a 4° C por toda la noche. Después el buffer fue cambiado por solución fresca y se dejó incubando por 2 horas más. La solución conteniendo la Er66 de las membranas de diálisis fue transferida a tubos de 1,5 mL de capacidad y las muestras fueron visualizadas por electroforesis en SDS-PAGE.
RESULTADOS
EVALUACIÓN DE LA INMUNOREACTIVIDAD DE LA PROTEÍNA ER66 DENATURADA
Los primeros resultados usando la proteína Er66 desnaturalizada en urea (8M) y con tratamientos usados de manera estándar para electroforesis de proteínas 31 , revelaron que sólo el anticuerpo MAB8701 presentó inmunoreactividad frente a la Er66 en los títulos de 1/50 y 1/25 (Figura 1, carril 2 y 3), en tanto, que ninguna señal de intensidad significante pudo ser visualizada a partir de una mezcla de sueros IgM (+) para fiebre amarilla (dilución 1/ 100). Estos resultados sugirieron que algunos o todos los agentes denaturantes empleados durante la purificación de la Er66 o la electroforesis podrían estar afectando de manera adversa la antigenicidad de la proteína recombinante. En ese sentido se utilizaron proteínas totales de la cepa estándar 17D Asibi (PT17D) tratadas o no con estos agentes desnaturalizantes. La proteína E nativa podría ser identificada por su peso molecular de aproximadamente 60 kDa.

En tal sentido, los resultados de Western Blot revelaron la presencia de dos bandas de aproximadamente 84 kDa y 60 kDa en las proteínas tratadas con b-mercaptoetanol, de 84 kDa en las proteínas sin ningún tratamiento, y de 84 kDa y 60 kDa sólo en las proteínas tratadas con urea (Figura 2). En contraste, cuando se combinó mercaptoetanol y urea o se usaron ambas condiciones conjuntamente con calor a 100 °C, la antigenicidad PT17D se perdió completamente.
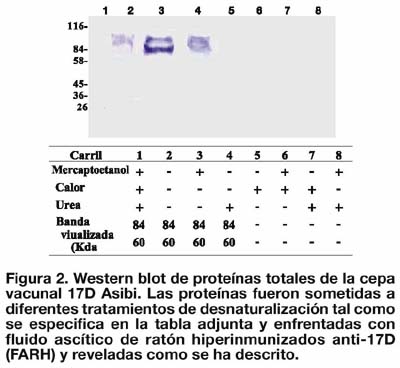
Estos resultados permitieron demostrar que la antigenicidad de la proteína E del virus FA 17D fue afectada negativamente mediante calor y con denaturación conjunta con mercaptoetanol y urea. En ese sentido, se decidió realizar la diálisis de Er66 para remover completamente las altas concentraciones de urea y excluir los tratamientos denaturantes con calor.
EVALUACIÓN DE LA INMUNOREACTIVIDAD DE LA PROTEÍNA ER66 RENATURADA
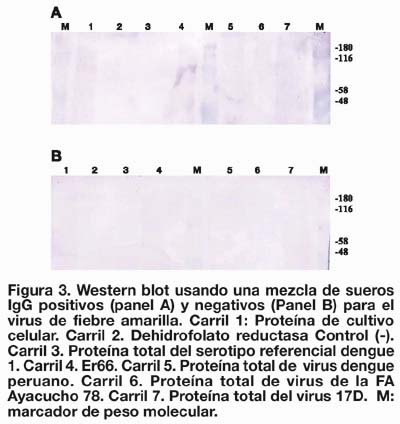
La proteína recombinante renaturada por díálisis fue enfrentada a una mezcla de sueros altamente reactivos para fiebre amarilla y dengue de acuerdo con los títulos de MAC y GAC ELISA (datos no mostrados). Conjuntamente a Er66 se consideró a PT17D, proteínas totales de dengue serotipo 1 de referencia de Hawai (PTDENR1) y proteínas totales de fiebre amarilla cepa peruana (PTFA-78) (Ayacucho 1978).
De acuerdo con los resultados obtenidos, se pudo visualizar una banda tenue correspondiente al peso molecular de la Er66. En contraste, no se observaron bandas bien definidas en los carriles correspondientes a PT17D, PTFA-78 y PTDENR1 (Figura 3).
INMUNOREACTIVIDAD DE LA PROTEÍNA ER66 FRENTE A SUEROS INDIVIDUALES DE PACIENTES IGM E IGG POSITIVOS PARA FIEBRE AMARILLA
Con el fin de evaluar individualmente la inmureactividad de cada suero positivo para fiebre amarilla, se utilizaron tiras de nitrocelulosa conteniendo la proteína Er66. De acuerdo con los resultados, se observó una fuerte señal a partir de una muestra clínica de paciente conteniendo IgG inmunoreactivo contra la fiebre amarilla, en tanto, que en las demás muestras IgG positivas incluyendo sueros positivos a dengue y controles negativos, no se pudo observar con nitidez la banda de interés.
Para determinar si la Er66 podría ser reconocida por anticuerpos IgM específicos para fiebre amarilla se seleccionó un grupo de muestras serológicas correspondientes al último brote de San Martín en el año 2003. De acuerdo con los resultados obtenidos, dos muestras positivas para FA (n=2) presentaron una fuerte inmunoreactividad frente a la Er66 (Figura 4, carril 5 y 10), mientras que en el resto de muestras, incluyendo los controles de especificidad para dengue, se observó una señal muy débil.
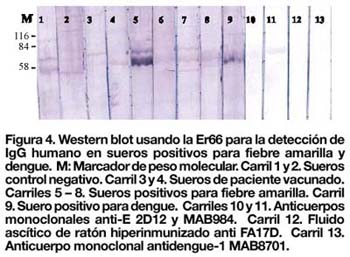
DISCUSIÓN
En el presente trabajo hemos realizado la evaluación serológica de una proteína recombinante denominada Er66, sintetizada a partir de una cepa aislada del virus de la fiebre amarilla en el Perú.
Los primeros resultados observados en el inmunoblot, después de enfrentar la Er66 con algunas muestras positivas para dengue y fiebre amarilla, no fueron satisfactorios debido a una posible falta de antigenicidad de esta proteína. Sin embargo, los datos obtenidos con anticuerpos monoclonales demostraron que la Er66 sí guardaba propiedades antigénicas por lo menos frente al anticuerpo monoclonal MAB984 y que la pérdida de su antigenicidad estaba relacionada con la desnaturalización por calor a 100 °C tal como fue demostrado en nuestro estudio.
Estos datos sugieren que la Er66 podría presentar epítopes de tipo estructural que al cambiar su conformación tridimensional por desnaturalización ya no es reconocida por los anticuerpos específicos. Esta afirmación se apoya en el hecho de que la mayoría de los epítopes de la proteína E del virus de la encefalitis asociada a los ácaros (TBEV), un flavivirus de la familia del virus de la fiebre amarilla, depende de sus características hidropáticas, es decir de su estructura tridimensional 27 . Sin embargo, la señal positiva débil que generó la Er66 frente a una mezcla de sueros inmunoreactivos para fiebre amarilla, probablemente, se debió al bajo título de los anticuerpos inmunogénicos producto de la mezcla realizada entre las muestras reactivas.
Nuestros resultados evaluando sueros individuales con anticuerpos IgG reactivos para la fiebre amarilla revelaron que una sola muestra de suero presentó una fuerte reactividad frente a la Er66, mientras que los demás no. Estos resultados fueron incongruentes con los altos valores de positividad obtenidos por MAC y GAC ELISA en los sueros evaluados (datos no mostrados). Las razones de este hecho no son muy claras aunque se podrían plantear diversas hipótesis. Una de ellas es la presencia de nuevos epítopes discretos en la proteína recombinante que sólo son reconocidos por una muestra clínica y no por las otras. De hecho, algunos autores han reportado nuevos epítopes de este tipo que recientemente han sido caracterizados en la proteína de la envoltura del virus de la FA cepa 17D 13 .
Otra hipótesis que podría explicar este hecho es la existencia de una afinidad inmunogénica por cercanía de espacios geográficos. La única muestra serológica que presentó una fuerte inmureactividad frente a la Er66 fue la de un paciente natural de Iquitos, zona de selva oriental del país. El resto de muestras que presentaron débil inmunoreactividad correspondieron a pacientes de Satipo, en la selva central del Perú. Probablemente, este hecho guarda relación con la procedencia de la Er66, la cual fue sintetizada a partir de un aislamiento viral proveniente de San Martín.
En ese sentido, se evaluaron sueros conteniendo IgM específicos para fiebre amarilla de pacientes provenientes de San Martín. Aunque no se logró determinar una fuerte reactividad en todas las muestras ni tampoco, capturar y registrar, todas las bandas deseadas, 100% de ellas (n= 8) mostraron inmunoreactividad frente a la Er66 en ensayos de western blot. Estos datos sugieren que esta proteínapodría tener mayor especificidad para muestras provenientes de San Martín; sin embargo, es necesario evaluar un mayor número de muestras serológicas de la selva nororiental y de otras áreas endémicas o enzoóticas del país para demostrar la sensibilidad y especificidad de la Er66.
Es importante mencionar que la Er66 no es antigénica frente a sueros de pacientes vacunados, este hecho podría potencial-mente discriminar pacientes vacunados de pacientes infectados durante el diagnóstico serológico de la fiebre amarilla. Todos estos datos en conjunto permiten demostrar que la Er66 es antigénica para anticuerpos monoclonales y muestras serológicas con altos títulos de anticuerpos IgM e IgG.
Los resultados mostrados en este reporte dejan pendientes varios aspectos por estudiar, uno de ellos es la probable existencia de anticuerpos que presentan una especificidad por ciertos epítopes localizados en la Er66, hasta el momento son desconocidos. Basándose en estos datos se podrían realizar estudios de mapeo epitópico de la proteína E y caracterizar epítopes discretos presentes en subpoblaciones de aislamientos virales de diferentes partes del país. Del mismo modo, otras proteínas del virus como NS1 y NS3 podrían ser sintetizadas y evaluadas serológicamente desde que se tienen datos actuales de sus propiedades de generar respuesta humoral y celular 28 .
Por último, podría ser interesante evaluar proteínas recombinantes expresadas a partir de otros aislamientos del virus de la fiebre amarilla de diferentes partes del país, y al mismo tiempo, incrementar el número de muestras positivas a evaluar. Todos estos procedimientos ayudarán a demostrar la presencia de epítopes específicos para determinados anticuerpos, y a seleccionar la mejor proteína recombinante que permita ser usada como herramienta de diagnóstico para la fiebre amarilla.
AGRADECIMIENTOS
Agradecemos el apoyo del Consejo Nacional de Ciencia y Tecnología (CONCYTEC) que subvencionó parcialmente el presente trabajo. Asimismo, al personal de la División de Virología, Laboratorio de Arbovirus del INS entre los que cabe mencionar a la TM María Paquita García Mendoza por proporcionarnos la cepa de virus de la fiebre amarilla de San Martín aislada en 1999, a la Blga. Victoria Gutiérrez Peceros, al Blgo Miguel Cobos Zelada, y al técnico Tomás Paredes Astupiña por su apoyo en facilitarnos las muestras serológicas. Asimismo, agradecemos al Blgo. Enrique Mamani por ayudarnos en la obtención del fluido ascítico de ratón hiperinmunizado y las muestras serológicas con anticuerpos IgM para fiebre amarilla del último brote del año 2003. Agradecemos también al NAMRID por proporcionarnos el anticuerpo monoclonal 2D12. A los biólogos Adolfo Marcelo y Vanesa Adawi de la Universidad Peruana Cayetano Heredia por facilitarnos las membranas de diálisis y al Sr. David García Neyra, personal técnico del laboratorio de Biología Molecular del Instituto Nacional de Salud (INS) por apoyar de manera entusiasta en el presente estudio.
REFERENCIAS BIBLIOGRÁFICAS
1. Tomori O. Impact of yellow fever on the developing world. Adv Virus Res 1999; 53: 5-34. [ Links ]
2. Rice CM, Lenches EM, Eddy SR, Shin SJ, Sheets RL, Strauss JH. Nucleotide sequence of yellow fever virus: implications for flavivirus gene expression and evolution. Science 1985; 229(4715): 726-33. [ Links ]
3. Reed KE, Gorbalenya AE, Rice CM. The NS5A/NS5 proteins of viruses from three genera of the family Flaviviridae are phosphorylated by associated serine/threonine kinases. J Virol 1998; 72(7): 6199-206. [ Links ]
4. Muylaert IR, Chambers TJ, Galler R, Rice CM. Mutagenesis of the N-linked glycosylation sites of the yellow fever virus NS1 protein: effects on virus replication and mouse neurovirulence. Virology 1996; 222(1):159-68 [ Links ]
5. Chambers TJ, Nestorowicz A, Rice CM. Mutagenesis of the yellow fever virus NS2B/3 cleavage site: determinants of cleavage site specificity and effects on polyprotein processing and viral replication. J Virol 1995; 69(3): 1600-5 [ Links ]
6. Brandriss MW, Schlesinger JJ, Walsh EE. Immunogenicity of a purified fragment of 17D yellow fever envelope protein. J Infect Dis 1990; 161(6): 1134-9 [ Links ]
7. Gould EA, Buckley A, Cane PA, Higgs S, Cammack N. Use of a monoclonal antibody specific for wild-type yellow fever virus to identify a wild-type antigenic variant in 17D vaccine pools. J Gen Virol 1989; 70 (Pt 7): 1889-94. [ Links ]
8. Desprès P, Girard M, Bouloy M. Characterization of yellow fever virus proteins E and NS1 expressed in Vero and Spodoptera frugiperda cells. J Gen Virol 1991; 72(Pt 6): 1331- 42. [ Links ]
9. Buckley A, Gould EA. Neutralization of yellow fever virus studied using monoclonal and polyclonal antibodies. J Gen Virol 1985; 66 (Pt 12): 2523-31. [ Links ]
10. Ballinger-Crabtree ME, Miller BR. Partial nucleotide sequence of South American yellow fever virus strain 1899/81: structural proteins and NS1. J Gen Virol 1990; 71(Pt 9): 2115-21. [ Links ]
11. Chambers T, Hahn C, Galler R, Rice C. Flavivirus genome organization, expresión, and replication. Annu Rev Microbiol 1990; 44: 649-88. [ Links ]
12. Mandl CW, Guirakhoo F, Holzmann H, Heinz FX, Kunz C. Antigenic structure of the flavivirus envelope protein E at the molecular level, using tick-borne encephalitis virus as a model.J Virol 1989; 63(2): 564-71 [ Links ]
13. Ryman KD, Ledger TN, Weir RC, Schlesinger JJ, Barrett AD. Yellow fever virus envelope protein has two discrete type-specific neutralizing epitopes. J Gen Virol 1997; 78 (Pt 6): 1353-6 [ Links ]
14. Shiu SY, Morikawa S, Buckley A, Higgs S, Karunakarannair V, Blachere C, et al. 17D yellow fever vaccine virus envelope protein expressed by recombinant baculovirus is antigenically indistinguishable from authentic viral protein. J Gen Virol 1991; 72 (Pt 6): 1451-4. [ Links ]
15. Pincus S, Mason PW, Konishi E, Fonseca BA, Shope RE, Rice CM, et al. Recombinant vaccinia virus producing the prM and E proteins of yellow fever virus protects mice from lethal yellow fever encephalitis. Virology 1992; 187(1): 290-7 [ Links ]
16. Desprès P, Dietrich J, Girard M, Bouloy M. Recombinant baculoviruses expressing yellow fever virus E and NS1 proteins elicit protective immunity in mice. J Gen Virol 1991; 72 (Pt 11): 2811-6. [ Links ]
17. Putnak JR, Schlesinger JJ. Protection of mice against yellow fever virus encephalitis by immunization with a vaccinia virus recombinant encoding the yellow fever virus non-structural proteins, NS1, NS2a and NS2b. J Gen Virol 1990; 71(Pt 8): 1697- 702. [ Links ]
18. Cane PA, Gould EA. Reduction of yellow fever virus mouse neurovirulence by immunization with a bacterially synthesized non-structural protein (NS1) fragment. J Gen Virol 1988; 69(Pt 6): 1241-6. [ Links ]
19. Makino Y, Tadano M, Arakaki S, Fukunaga T. Potential use of a baculovirus-expressed dengue-4 E protein as a diagnostic antigen in regions endemic for dengue and Japanese encephalitis. Am J Trop Med Hyg 1991; 45(5): 636-43. [ Links ]
20. Konishi E, Mason PW, Shope RE. Enzyme-linked immunosorbent assay using recombinant antigens for serodiagnosis of Japanese encephalitis. J Med Virol 1996; 48(1): 76-9. [ Links ]
21. Simmons M, Porter KR, Escamilla J, Graham R, Watts DM, Eckels KH, et al. Evaluation of recombinant dengue viral envelope B domain protein antigens for the detection of dengue complex-specific antibodies. Am J Trop Med Hyg 1998; 58(2): 144-51. [ Links ]
22. Cuzzubbo AJ, Endy TP, Nisalak A, Kalayanarooj S, Vaughn DW, Ogata SA, et al. Use of recombinant envelope proteins for serological diagnosis of Dengue virus infection in an immunochromatographic assay. Clin Diagn Lab Immunol 2001; 8(6): 1150-5. [ Links ]
23. Yábar C, Montoya Y. Síntesis in vitro de la proteína de la envoltura del virus peruano de la fiebre amarilla. Rev med exp 2001; 18(1-2): 9-3 [ Links ]
24. Brown TM, Chang GJ, Croop CB, Robbins KE, Tsai TF. Detection of Yellow fever virus by polymerase chain reaction. Clin Diagn Virol 1994; 2: 41-51 [ Links ]
25. Roehrig JT, Bolin RA, Kelly RG. Monoclonal antibody mapping of the envelope glycoprotein of the dengue 2 virus, Jamaica. Virology 1998; 246(2): 317-28. [ Links ]
26. QIAGEN. The QIAexpressionist. A handbook for high-level expression and purification of 6xHis-tagged proteins. 3th ed. Hilden (GE): Qiagen; 1997. [ Links ]
27. Rey FA, Heinz FX, Mandl C, Kunz C, Harrison SC. The envelope glycoprotein from tick-borne encephalitis virus at 2 A resolution. Nature 1995; 375(6529): 291-8 [ Links ]
28. Monath TP. Yellow fever: an update. Lancet Infect Dis 2001; 1(1): 11-20. [ Links ]
29. Schlesinger JJ, Brandriss MW, Monath TP. Monoclonal antibodies distinguish between wild and vaccine strains of yellow fever virus by neutralization, hemagglutination inhibition, and immune precipitation of the virus envelope protein. Virology 1983; 125(1): 8-17. [ Links ]
30. Henchal EA, Gentry MK, McCown JM, Brandt WE. Dengue virus-specific and Flavivirus group determinants identified with monoclonal antibodies by indirect immunofluorescence. Am J Trop Med Hyg 1982; 31(4): 830-6 [ Links ]
31. Yábar VC. Manual de procedimientos de electroforesis paraproteínas y ADN. Lima: Ministerio de Salud, Instituto Nacional de Salud; 2003. Serie de Normas Técnicas 38. [ Links ]
Correspondencia: Carlos Yábar V.
División de Biología Molecular, Centro Nacional de Salud Pública.
Instituto Nacional de Salud.
Dirección:Av. Cápac Yupanqui 1400. Lima 11, Perú.
Teléfono: (511) 4719920 Anexo: 129
Correo electrónico: cyabar@ins.gob.pe

