










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
Print version ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.22 no.4 Lima Oct./dic 2005
TRABAJOS ORIGINALES
Estudio Ultraestructural de Leptospira Biflexa Serovar Andamana cepa JNS al Microscopio Electrónico de Transmisión y Barrido
Sonia Macedo A1,2
1 Docente Asociado. Universidad Wiener. Lima, Perú.
2 Ex jefa del Laboratorio de Leptospirosis, Instituto de Medicina Tropical «Daniel A. Carrión», Facultad de Medicina, Universidad Nacional Mayor de San Marcos. Lima, Perú.
RESUMEN
Objetivos: Investigar la ultraestructura de Leptospira biflexa serovar Andamana cepa JNS, por la técnica de réplica metálica, cortes ultrafinos y tinción negativa al microscopio electrónico de transmisión (MET) y barrido (MEB) en alta y ultraalta resolución. Materiales y métodos: Cultivos en fase logarítmica 12-15x106 cel/mL del serovar Andamana cepa JNS, aislada de un paciente. Se aplicaron los métodos de tinción negativa, cortes ultrafinos y réplica metálica para la microscopía electrónica de transmisión. Para la microscopía electrónica de barrido se usaron las técnicas en alta y ultraalta resolución de muestras cubiertas con oro y platino. Resultados: Con el método de réplica metálica se definió la estructura externa en células individuales de Leptospira biflexa serovar Andamana al MET. Mediante la técnica de tinción negativa se observó la membrana externa formada por tres capas, los filamentos axiales, la membrana citoplasmática, el citoplasma granular con inclusiones electronodensas, las formaciones «globosas», la «estructura terminal», «apéndices terminales», además de las inclusiones permeables para los electrones, claramente definidas. La morfología clásica del microorganismo fue observada en cortes finos, y al microscopio electrónico de barrido. Conclusiones: Se reporta por primera vez la técnica de réplica metálica para determinar la morfología externa en células individuales de Leptospira biflexa serovar Andamana JNS, la cual ofrece resultados ventajosos en comparación con otros métodos. La ultraestructura, y las inclusiones permeables para electrones dentro del citoplasma a una magnificación de 120000x 20KV fueron nítidamente observadas por tinción negativa. La estructura helicoidal del microorganismo se comprobó al MEB.
Palabras clave: Microscopía electrónica de transmisión, barrido: Leptospira biflexa serovar Andamana (Fuente: BIREME).
ABSTRACT
Objectives: To investigate Leptospira biflexa serovar Andamana, JNS strain ultrastructure using the metallic replication technique, ultrafine cuts, and negative staining for transmission electron microscope and scanning electron microscope high and ultra-high resolution imaging. Materials and methods: Andamana serovar, JNS strain logarithmic phase cultures, 12–15 x 106 cell/mL isolated from one patient were used. Negative staining, ultrafine slicing and metallic replication methods were used for transmission electron microscopy. For scanning electron microscopy, high and ultra-high resolution samples with gold and platinum coating were used. Results: Using the metallic replication method the external structure of individual cells was defined for Leptospira biflexa, Andamana serovar using transmission electron microscopy. Using the negative staining technique the three-layer external membrane could be seen, as well as axial filaments, cytoplasmic membrane, granular cytoplasm with electron-dense inclusions, «balloon» formations, «terminal structure» «terminal appendages», and also the electron-permeable inclusions, all of these were clearly defined. The typical morphology of the microorganism was observed using fine slices on scanning electron microscopy. Conclusions: For the first time we report the metallic replication technique for determining the external morphology in individual Leptospira biflexa, Andamana serovar, JNS strain, and this technique offers advantageous results compared to other methods. Ultrastructure and electron-permeable inclusions within the cytoplasm at a 120 000 power 20 KV were clearly seen using negative staining. Scanning electron microscopy was also useful for confirming the helical structure of the microorganism.
Key words: Transmission and scanning electron microscopy: Leptospira biflexa serovar Andamana. (Source:
BIREME).
INTRODUCCIÓN
La leptospirosis es una zoonosis reportada mundialmente con una mayor incidencia en países con clima húmedo y tropical1. Se han notificado serovares endémicos en Europa, Norteamérica, Australia, Sudamérica, el Caribe y Asia1-4. Los agentes etiológicos pertenecen al dominio bacteria, phylum Spirochaetes, clase Spirochaetes, orden Spirochaetales, familia Leptospiraceae, género Leptospira con trece especies y género Leptonema5-8 . Según la clasificación por genomospecies se han notificado nueve patógenas y una no patógena Leptospira biflexa9.
La primera cepa de Leptospira biflexa serovar Andamana cepa CH 11 fue aislada de un paciente por Taylor y Goyle en 1930 en las Islas Andamán10, décadas después se notificaron casos humanos en el Brasil10,11 y Perú12.
La apariencia morfológica de las espiroquetas es básicamente la misma para el género Leptospira, la observación de las cuales generalmente se realizan al microscopio de campo oscuro13. Las investigaciones con el microscopio electrónico de transmisión permitieron demostrar la presencia de tres principales estructuras: la membrana externa o cubierta; el cilindro citoplasmático y los dos flagelos o filamentos axiales uno en cada extremo de la célula14-16.
La configuración de forma helicoidal y su gran longitud (3-20 mm) comparada con su diámetro de 0,1mm (15:1 a 50:1), hacen que las leptospiras sean difíciles de examinar en su totalidad en secciones ultrafinas15, sin embargo, las técnicas de tinción negativa, y recubrimiento con oro y platino, han permitido describir las estructuras de Leptospira interrogans serovar Icterohaemorragiae17,21, cepa B1620, serovar Pomona15,18,21, serovar Canicola19, serovar Hardjo21 y de Leptospira biflexa serovar Patoc21, y cepa B.SH17.
El objetivo del presente trabajo fue investigar la ultraestructura de Leptospira biflexa serovar Andamana cepa JNS, aislada de un paciente12, por los métodos de réplica metálica de superficie, tinción negativa y cortes ultrafinos al microscopio electrónico de transmisión. La morfología externa se estudió al microscopio electrónico de barrido en alta y ultraalta resolución.
MATERIALES Y MÉTODOS
MATERIALES
Microorganismo
Leptospira biflexa serovar Andamana cepa JNS, aislada de un paciente por hemocultivo. El cultivo desarrollado a 28 °C se procesó en fase logarítmica 12- 15x106 cel/mL en el medio líquido de Korthof: peptona 1.0 g, NaCl 1,4 g, carbonato ácido de sodio (NaHCO3) 0,02 g, cloruro cálcico (CaCl2) 0.04 g, cloruro potásico (KCl) 0,04 g, fosfato diácido de potasio (KHPO4) 0,18 g, fosfato ácido de sodio (Na2HPO4H2O) 0.96 g, agua bidestilada 1000 mL pH 7,3, con adición final de suero de conejo al 10% inactivado a 56 ºC durante 30 minutos.
Microscopía electrónica de transmisión (MET)
Cortes ultrafinos. Se usó glutaraldehido 1,5%, fijador de Caulfield pH 7,4, solución tampón de Caulfield, alcoholes en concentraciones crecientes del 30 al 100%, óxido de propileno y resina Spurr. Cortes ultrafinos seriados de color plateado colectadas en rejillas con membranas de soporte de colodión.
Tinción negativa. Se empleó el ácido fosfotúngstico H3[P(W3O10]4) x H20 al 2% con la adición de sacarosa al 0,04%. Las rejillas se cubrieron con membranas de soporte de formar-carbón.
Réplica metálica. Se aplicó glutaraldehido 1% en solución amortiguadora de cacodilato de sodio 1%, pH 7,4, solución tampón de cacodilato de sodio 1% pH 7,4 más sacarosa 3,75%, alcoholes 50, 70, 90 y 100%, recubrimiento de la muestra con platino-carbón, cemento plástico, hipoclorito de sodio (NaOCl) 10% como oxidante, agua bidestilada y acetona. Las rejillas fueron recubiertas con carbón-platino-carbón.
Microscopía electrónica de barrido (MEB
Se usó glutaraldehido 1% en buffer cacodilato de sodio 1% pH 7,4 más sacarosa 3,75%, solución tampón de cacodilato de sodio 1% pH 7,4, alcoholes en concentraciones de 50, 70, 90 y 100%. Las bases para ultraalta resolución con el material fueron recubiertas con platino 150 Å y las bases para alta resolución se recubrieron con oro 200 Å.
MÉTODOS
Microscopía electrónica de transmisión (MET)
Cultivo bacteriano. La cepa de la especie Leptospira biflexa serovar Andamana JNS se subcultivó en el medio líquido de Korthof cada 48-72 horas en fase logarítmica de crecimiento a 28 °C, determinándose la concentración de células 12-15x106 /mL en la cámara de Petroff-Hausser. La pureza del cultivo se controló por observación directa al microscopio de campo oscuro y siembra por inoculación en caldo tripticase y agar sangre. Este cultivo fue uso para todas las técnicas descritas.
Réplica metálica de superficie. Se usó el método descrito por Ureña et al.22 con algunas modificaciones. La muestra fue centrifugada a 5000 g durante 20 minutos a 4 °C, el sedimento fue fijado con glutaraldehido 1% en buffer cacodilato de sodio 1% y sacarosa 3,75% durante una hora a 20 °C. Luego se deshidrató en una serie de alcoholes de 50, 70, 90 y 100 %. Inmediatamente, el sedimento fue centrifugado 10 minutos a 1500 r.p.m., y 1-2 gotas del sedimento fueron transferidas a un cubreobjeto previamente descargado eléctricamente en el cobertor iónico (Marca Eico IB-3®), dejando secar la muestra a temperatura ambiente. Enseguida se sombreó por rotación con platino-carbón, evaporándolo a 10-5 torr con un ángulo aproximado de 10°, seguidamente fue cubierta con cemento plástico. Una vez solidificado se separó y fue sumergido en hipoclorito de sodio al 10% para eliminar todas las sustancias orgánicas (proceso de oxidación). Luego fue lavada con agua destilada y se recubrió de nuevo por rotación con carbón. El corte se hizo con una aguja, se eliminó el plástico con acetona y se obtuvo la preparación final de réplica carbón-platino- carbón y fue lavada repetidamente con acetona. Finalmente se recogió la réplica en rejillas y se observaron al microscopio electrónico modelo H-300 a 80Kv (Hitachi®, Japón).
Tinción negativa. El cultivo se pasó a través de un filtro Millipore 0,45 µm para eliminar los residuos del medio de cultivo, luego fue centrifugada a 3000 r.p.m por 15 minutos a temperatura ambiente y se separó el sedimento en dos tubos, uno de ellos con ácido fosfotúngstico al 2%, y el otro con amortiguador de fosfatos 0,1 M pH 7.4, al cual se añadió ácido fosfotúngstico al 2%. Inmediatamente se colocaron dos a tres gotas de ambas suspensiones sobre una placa Petri cubierta con parafilm, y las rejillas recubiertas con membrana de soporte de formar-carbón fueron impregnadas con cada una de las suspensiones dejándose durante 30 segundos, un minuto y tres minutos. Luego, se dejaron secar durante 24 horas, y se observaron al microscopio electrónico modelo H-300 y HU-13 (Hitachi®, Japón).
Cortes ultrafinos. La metodología usada fue similar a las descritas previamente23,24 con algunas modificaciones. Al tubo con 5 mL de cultivo se añadió 5 mL de glutaraldehido 1,5% de concentración final, luego fueron centrifugadas a 5000 g por 20 minutos a 4 °C y se eliminó el sobrenadante. Al sedimento se añadió el fijador de Caulfeld durante dos horas a 4 °C , luego, se centrifugó y el sedimento fue transferido a las cápsulas y se deshidrataron en una serie de alcoholes de 30 a 100%. Enseguida, se infiltró e incluyó en resina Spurr durante 72 horas para la polimerización. Los cortes se realizaron en el ultramicrótomo Sorvall MT-2B, se colorearon con acetato de uranilo y de Reynold. La observación se realizó después de 24 horas al microscopio electrónico modelo HU-12 A (Hitachi®, Japón) a 80Kv.
Microscopía electrónica de barrido (MEB)
El proceso de fijación fue similar al de la réplica metálica. El sedimento obtenido en la última deshidratación, fue colocado directamente sobre un cubreobjeto adherido con plata metálica a la base y se recubrió con oro durante cuatro minutos con un grosor de 200 Å en el cobertor iónico (marca Eico IB-3®), para ser observada con alta resolución. Paralelamente otra muestra se puso en una base inclinada con un ángulo aproximado de 45° y se recubrió con platino por tres minutos con un grosor de 150 Å. Ambas preparaciones fueron observadas al microscopio electrónico de barrido S-570 (Hitachi®, Japón).
RESULTADOS
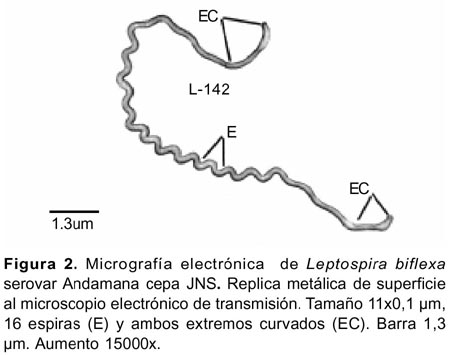
La técnica de réplica metálica de superficie22 se aplicó por primera vez para el estudio de Leptospira al microscopio electrónico de transmisión. El método permitió observar nítidamente la morfología externa de células individuales de Leptospira biflexa serovar Andamana cepa JNS en fase logarítmica de crecimiento. En las preparaciones las células presentaron forma helicoidal de 11 x 0,1µm, con 16 espiras estrechamente unidas, con uno o ambos extremos curvados (Figuras 2 y 3).
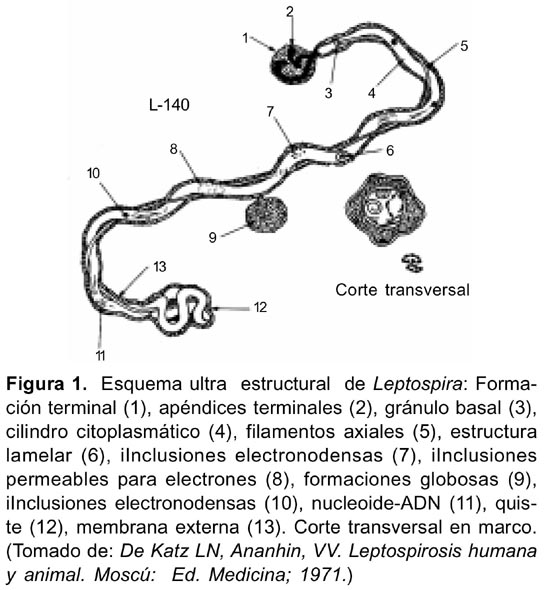

En células sometidas a la tinción negativa con ácido fosfotúngstico al 2%, se observaron las ultraestructuras del género Leptospira referidas por Katz y Ananhin (Figura 1)16. La membrana externa que cubre al microorganismo fue observada en ciertas áreas tanto en células completas (Figuras 4 y 5) como fraccionadas (Figuras 6-8), Leptospira biflexa serovar Andamana cepa JNS presentó dos flagelos o filamentos axiales localizados entre la membrana externa y el cilindro citoplasmático, cada uno nacía de un solo gránulo basal de localización subterminal (Figura 6), y se enrollaba alrededor del cilindro citoplasmático. En la parte media de las células no se observó esta estructura (Figura 4,5).
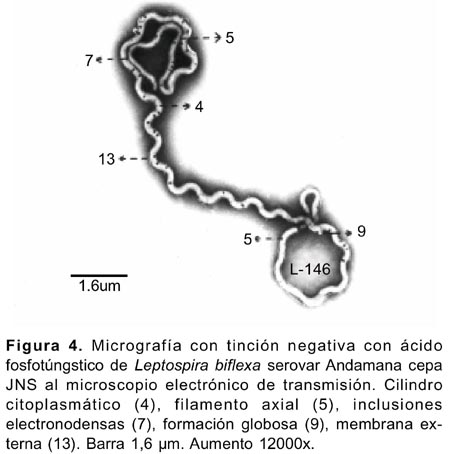

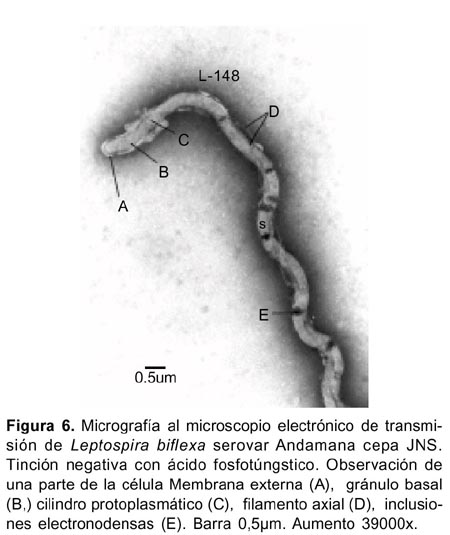
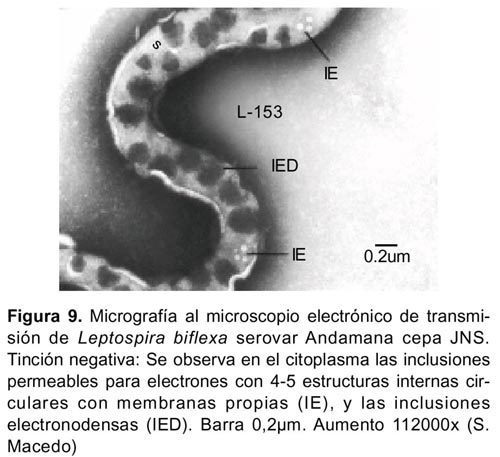
La célula conservó su forma helicoidal aún cuando los flagelos o filamentos axiales se separaron de la célula (Figura10). La membrana citoplasmática se localizó adherida al cilindro citoplasmático (Figura 8). El citoplasma de las células mostró configuración granular con presencia de inclusiones granulares electronodensas de diversos tamaños (Figura 4-6,9) e inclusiones permeables para los electrones observadas a 112000x 20KV, separadas por una membrana, y en interior de estas se advirtió cuatro o cinco estructuras circulares de diversos tamaños delimitadas también por sus propias membranas (Figura 9).
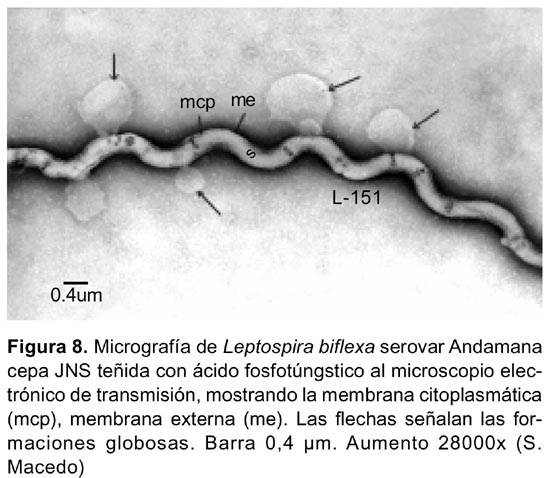
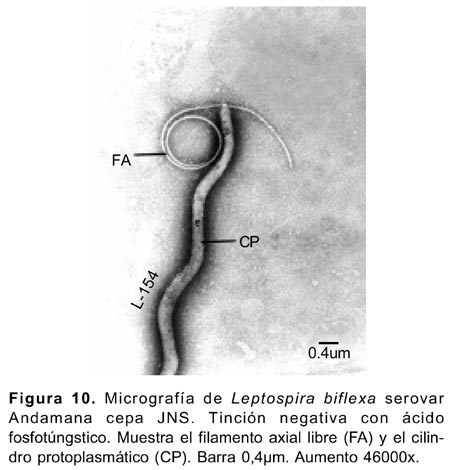
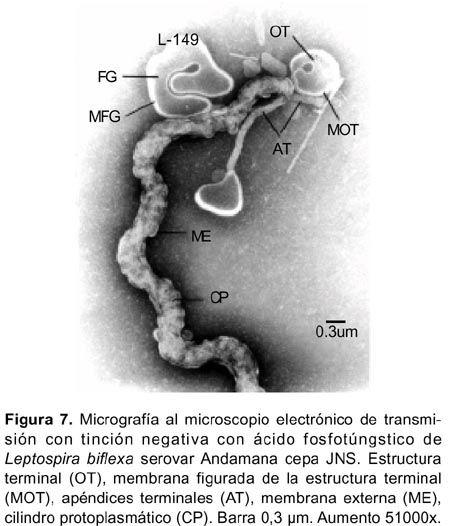
Las formaciones «globosas vacías» con o sin una membrana delimitante, se observaron en algunas células y se originaban a partir de la membrana externa o de la membrana citoplasmática (Figura 4,5,8). Los denominados «estructura terminal» delimitado por una membrana figurada, el «quiste» y los «apéndices terminales », se observaron en la región terminal en algunas células (Figura 5,7).
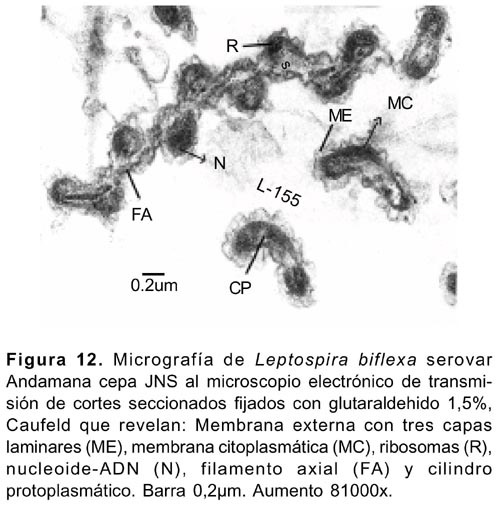
Las micrografías de los cortes ultrafinos al microscopio electrónico de transmisión, mostraron la presencia de la membrana externa con capas triples de apariencia ondulada, la membrana citoplasmática menores a 50 Å, y los flagelos o filamentos axiales entre la última membrana y el cilindro citoplasmático.
El citoplasma mostró naturaleza granular con presencia del nucleoide conteniendo el ADN, y el ARN disperso (Figura 12).
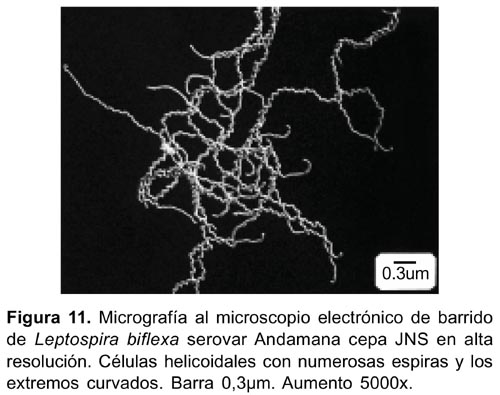
En preparaciones al microscopio electrónico de barrido, se observó la morfología superficial, generalmente de células agrupadas, entrelazadas, con uno o ambos extremos curvados y sumergidas dentro de la capa metálica de oro y platino (Figura11).
DISCUSIÓN
El presente estudio es el primero que se realiza en Perú al microscopio electrónico de transmisión y de barrido del serovar Andamana JNS aislada de un paciente12, y cuyas características fenotípicas corresponden a la genomospecie no patógena Leptospira biflexa serovar Andamana25.
La aplicación de la técnica de réplica metálica de superficie carbón-platino-carbón ha sido reportada para el estudio de espermatozoides de ratas al microscopio electrónico de transmisión22. Este método es aplicado por primera vez para el estudio de Leptospira, el cual permitió definir claramente y con detalle la morfología externa de Leptospira biflexa serovar Andamana cepa JNS, con relación a la forma helicoidal flexible, la longitud, el diámetro, el número de espiras estrechamente unidas, y la disposición curvada de uno o ambos extremos, en cultivos en fase logarítmica 12x106 cel/mL.
La observación de la ultraestructura al microscopio electrónico de barrido en ultraalta resolución mostró la morfología externa y característica de las espiroquetas con gran nitidez por el recubrimiento con las capas metálicas de oro y platino. Esta técnica ha sido aplicada para determinar la condición de giro de las espiras en varios serogrupos de Leptospiras 26 .
El método de tinción negativa con diferentes colorantes ha sido el procedimiento más empleado para la descripción de la ultraestructura de Leptospira interrogans serovar Pomona15,18,21, cepa B1620, serovar Icterohaemorragiae17,21, serovar Hardjo21, serovar Canicola19, y de Leptospira biflexa cepa B SH17 y serovar Patoc21. En este estudio el análisis de la ultraestructura de Leptospira biflexa serovar Andamana cepa JNS, por el método de tinción negativa con ácido fosfotúngstico fue simple y rápida. El colorante confirió a las preparaciones un buen contraste y resolución de detalle de los especimenes permitiendo determinar las estructuras señaladas en investigaciones previas en otros serovares al microscopio electrónico1,15,16,20,21 . La membrana externa puede ser removida total o parcialmente con relativa facilidad, y a menudo inadvertida, durante el lavado y la manipulación de los cultivos1. El serovar en estudio fue filtrado y sometido a centrifugación, lo que habría producido este efecto, ya que en la mayoría de las preparaciones ésta sólo fue observada en determinadas áreas del microorganismo.
Las formaciones globosas cuyo diámetro es mucho mayor al del microorganismo, y se originan en la membrana externa en circunstancias no definidas y localizadas indistintamente a lo largo de la célula han sido observadas en otros serovares. Se desconocen su función y se presume que se formen debido a distensiones de la membrana y se desarrollen por presiones internas1,15,20.
Es conocido que las Leptospiras realizan movimientos de «no-translación» en la cual el cuerpo permanece relativamente estacionario, con un extremo ondulatorio en movimiento circular en dirección opuesta. Cuando el movimiento es de translación el extremo hacia el cual se efectúa la translación, realiza ondas helicoidales viajando a corta distancia de la célula hacia el extremo que se arrastra, formando un amplio gancho que se agita en un círculo irregular en la dirección opuesta a la onda28. Los filamentos axiales o flagelos que mostró el serovar Andamana JNS, y que son responsables de la motilidad estaban insertados en los gránulos básales localizados en los extremos del cilindro protoplasmático, se enrollaban alrededor de éste último, y no se cruzaban en la parte media de la célula. Resultados similares han sido reportados en otros serovares1,15,17,20. La separación accidental de los flagelos en algunas células demostró que estos cumplen función locomotora y no de soporte20, ya que la forma helicoidal de la célula se mantiene.
El citoplasma de Leptospira posee las inclusiones características para las bacterias, sin embargo frecuentemente se observan inclusiones densas de diferentes tamaños e inclusiones permeables para los electrones que han sido identificados en diferentes tipos de serovares16,20. Leptospira biflexa serovar Andamana cepa JNS mostró en casi todas las preparaciones con tinción negativa ambos tipos de inclusiones, las primeras se observaron dispuestas regularmente en el cilindro citoplasmático, en cambio las inclusiones permeables para electrones se observaron notoriamente a aumentos de 112000x 20KV separadas por una aparente membrana, y en cuyo interior se advirtió cuatro a cinco estructuras circulares de diversos tamaños delimitadas por sus propias membranas. La compleja configuración y la naturaleza química de estas inclusiones se encuentran en discusión16.
Las ultraestructuras determinadas en los cortes seccionados como son: la membrana externa con tres capas laminares de aspecto ondulante, la membrana citoplasmática que envuelve al cilindro protoplasmático, el nucleoide conteniendo el ADN, granulaciones electronodensas del ARN, y el filamento axial, son similares a los observados en serovares no patógenos16, y patógenos15.
La presencia de los llamados «órgano terminal», los apéndices terminales» y el «quiste» cuyos papeles fisiológicos se mantienen en discusión, coinciden con los reportados para el serovar Patoc17 que también es no patógena, y el serovar Pomona15.
La presente investigación permitió determinar con detalle la morfología externa de Leptospira biflexa serovar Andamana cepa JNS por la técnica de réplica metálica de superficie con carbón-platino-carbón al microscopio electrónico de transmisión, y fue aplicada por primera vez para el estudio estructural del género Leptospira. La técnica de la tinción negativa permitió determinar las ultraestructuras características para el microorganismo, sin embargo en la literatura consultada no se señala un estudio de análisis profundo sobre la compleja configuración y composición química de las inclusiones permeables para los electrones.
AGRADECIMIENTO
Al Sr. Carlos Hurtado de Mendoza S.
REFERENCIAS BIBLIOGRÁFICAS
1.Faine S, Adler B, Bolin C, Perolat P. Leptospira and Leptospirosis. 2nd ed., Melboure: MediSci; 2000. p. 11-28. [ Links ]
2.Macedo S, Cornide R, Cáceres I. Cepas endémicas de Leptospiras patógenas aisladas en América Latina y el Caribe. Rev Cub Med Trop 1983; 35:186-192
3. Alexander A, Wetmore P, Evans L, Jeffries H, Gleiser Ch. Classification of Leptospira isolates from Malaya, Thailand and North-Borneo. Am J Trop Med Hyg1955; 4(5): 492-506.
4. Kmety E, Dikkens H. Classification of the species Leptospira interrogans and history of its serovars. Groningen-Netherlands: University Press Groningen; 1993. p. 17-71.
5. Faine S, Stallman ND. Amended descriptions of the genus Leptospira Noguchi 1917 and the species L. interrogans (Stimson 1907) Wenyon 1926 and L. biflexa (Wolfach and Binger 1914) Noguchi 1918. Int J Syst Bacteriol 1982; 32(4): 461-63.
6. Hovind-Hougen K. Leptospiraceae, a new family to include Leptospira Noguchi 1917 and Leptonema gen. Int J Syst Bacteriol 1979; 29: 245-51.
7. Marshall R. International Subcommittee on Systematic Bacteriology. Subcommitteon the Taxonomy of Leptospira. Minutes of the meetings. 13-15 September 1990. Inter J Syst Bacteriol 1992; 42: 330-34.
8. Garriti GM. Taxonomic outline of the Prokaryotes. In: Bergeys Manual of Systematic Bacteriology, 2nd ed. New York: Springer-Verlag; 2002.
9. Weyant R, Bragg S, Kaufmann H. Leptospira and Leptonema Taxonomy. En: Murray P. Manual of Clinical Microbiology 7th ed. Washington DC: American Society of Microbiology 1999. p. 739-45.
10. Correa MOA, Hyakutake S, Natale V, Galvao PAA, Tiriba AC. Leptospiroses humanas ainda nã assinaladas no Brasil. Rev Inst Med Trop Sao Paulo 1964; 6(1):71-74.
11. Correa M, Hyakutake S, Natale V., Tiriba da Cruz. V, Martirani I, Galvao PAA, et al. Leptospirose humana por Leptospira andamana. Rev Inst Med Trop Sao Paulo 1971; 18(2): 137-43.
12. Macedo S, Isasi CR, Reyes N, Valencia E. Primer aislamiento en humano de Leptospira andamana (L. biflexa) en el Perú. En: Resúmenes VIII Congreso Peruano de Microbiología y Parasitología. Lima: Edit. PROPASEB; 1990. p. 44
13. Wolf JW. The laboratory diagnosis of Leptospirosis. Springfield, Illinois: Thoma; 1954.
14. Babudieri B. The morphology of the genus Leptospira as show by the electron microscope. J Hyg 1949; 47:390-392.
15. Ritchie AE, Ellinghausen HC. Anatomical features of Leptospira pomona. J Bacteriol 1964; 98(1):223-33.
16. Katz LN, Ananhin VV. Morfología y organización submicroscópica de Leptospira. En: Leptospirosis Humana y animal. Moscú: Ed. Medicina; 1971. p. 16-40. [en ruso].
17. Kiktenko VS, Troshin KA. Leptospira morphology studied by electron microscopy. J Microb Epidemiol Immunobiol 1966; 2: 85-90.
18. Miller N, Wilson R. In vivo and in vitro observations of Leptospira pomona by electron microscopy. J Bacteriol 1962; 84(3): 569-76.
19. Swain R. The electron microscopal anatomy of Leptospira canicola. J Bacteriol 1957; 73:155-58.
20. Holt SC. Anatomy and chemistry of Spirochaetes. Microbiol Rev 1978; 42(1): 114-60.
21. Hovind-Hougen K. Morphology of Leptospires. Electron microscopic studies in relation of the classification of Leptospira. In: Ellis WA, Little TWA. The present state of leptospirosis diagnosis and control. Dordrecht: Martinus Nijhoff Publishers; 1984. p. 1-12.
22. Ureña F. An improve method for the surface replica. J Electron Microsc 1985; 34(2):144-45.
23. Kushida H. A new embedding method for ultrathin sectioning using a methacrylate resin with three dimensional polymer structures. J Electron Microsc (Tokyo) 1961; 10:194-97.
24. González-Santander R. Técnicas de microscopía electrónica en biología. Madrid: Edit. Aguilar; 1968. p. 26-36.
25. Cinco M, Dougan R. Factor analysis of saprophytic serogroups semaranga and andamana of Leptospira biflexa. Inter J Syst Bact 1975; 25: 138-41.
26. Carleton O, Charon NW, Allender P, OBrien S. Helix handedness of Leptospira interrogans as determined by scanning electron microscopy. J Bacteriol 1979; 137(3):1413-16.
27. Cox PJ, Twigg GI. Leptospiral motility. Nature 1974; 250(463): 260-26.
Correspondencia: Sonia Macedo Aguirre PhD. Universidad Wiener, Lima, Perú.
Dirección: Av. Arequipa 440, Lima 1, Perú.
Teléfono: 4339119 anexo 114
Correo electrónico: sma1611@yahoo.com, smacedo@wienergroup.com

