










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
Print version ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.25 no.2 Lima Apr./jun. 2008
Efecto del antiveneno botrópico sobre las actividades de fosfolipasa a2, l-aminoácido oxidasa y hialuronidasa de los venenos de serpientes peruanas
Effect of polivalent bothropic antivenom on phospholipase a2, l- amino acid oxidase and hyaluronidase from peruvian snake venom
Julio Cesar Mendoza1,a, Fanny Lazo1,a, Liliana Yarlequé1,b, Nora Cecilia Ruiz1,a, Armando Yarlequé1,a, Silvia Pessah2,c, Vicky Flores2,d, César Bonilla2,a
1 Laboratorio de Biología Molecular, Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Lima, Perú.
2 Centro Nacional de Productos Biológicos, Instituto Nacional de Salud. Lima, Perú.
a Biólogo; b Obstetriz; c Médico; d Químico farmaceútico.
Estudio financiado por el Instituto Nacional de Salud del Perú.
RESUMEN
Las serpientes Bothrops sp. causan el mayor número de casos de ofidismo en el Perú, su veneno contiene enzimas que participan en la difusión de la ponzoña, así como en sus efectos miotóxicos, edemáticos y de alteración en la agregación plaquetaria. Objetivos. Evaluar el efecto del antiveneno botrópico polivalente al estado líquido producido por el Instituto Nacional de Salud (INS) sobre la fosfolipasa A2 (PLA2), L-aminoácido oxidasa (LAO) y hialuronidasa (HA) de los venenos de B. atrox, B. barnetti, B. brazili y B. pictus. Materiales y métodos. La PLA2 fue determinada por el retardo en el tiempo de coagulación de una emulsión lipoproteica al 45%, LAO usando Lleucina como substrato en presencia de O-dianisidina y HA empleando ácido hialurónico y el reactivo turbidimétrico BCTA, se usó para cada enzima ½, 1 y 2 dosis del antiveneno al estado natural o calentado a 37 °C durante cinco días ensayados por triplicado. Resultados. HA fue la enzima más neutralizada por el antiveneno, todos los venenos con excepción de B. brazili fueron totalmente inhibidos a cualquier dosis. Para LAO se tuvieron valores de inhibición de 68 a 100% usando dos dosis del antiveneno, mientras que PLA2 fue la menos inhibida (70 a 80%) a dos dosis. Con el antiveneno calentado se registró una disminución del efecto inhibitorio encontrado inicialmente. Conclusiones. La medición de la HA podría servir como indicador in vitro de la potencia del antiveneno, el antiveneno producido por el INS guarda las condiciones in vitro de inhibición de tres de las principales actividades de los venenos de serpientes peruanas.
Palabras clave: Venenos de serpiente; Hialuronoglucosaminidasa; L-aminoácido oxidasa; Fosfolipasas A2; Antivenenos (fuente: DeCS BIREME).
ABSTRACT
Bothrops sp. snakes causing the largest number of cases of ophidism in Peru, their venom contain several enzymes related to poison spreading, miotoxic and platelet aggregation disturbances. Objectives. The inhibiting capacity of liquid polivalent bothropic antivenom from Instituto Nacional de Salud (INS) has been evaluated on phospholipase A2 (PLA2), L amino acid oxidase (LAO) and hyaluronidase activities using B. atrox, B. barnetti, B. brazili and B. pictus venoms. Material and methods. In each case on 45% egg yolk lipoprotein, Lleucina and O-dianisidine, as well as hyaluronic acid as substrates respectively, using for each enzyme ½, 1 and 2 doses of either natural and heated (37 °C during five days) antivenom, assayed in triplicate. Results. HA was more neutralized enzyme for antivenin, all venoms with the exception of B. brazili were totally inhibited at any dose. For LAO had values of inhibition of 68 to 100% using two doses of the antivenin, PLA2 was the least inhibited (70 to 80%) to two doses. With the heated antivenin was a decline of the inhibitory effect initially found. Conclusions. The measurement of the HA might serve as an indicator of the in vitro potency of antivenins, the bothropic antivenin produced by INS keeping in vitro conditions for inhibition of three major activities of the Peruvian snake venoms.
Key words: Snake venoms; Hyaluronoglucosaminidase; Phospholidases A2; Hialuronidase; L-amino acid oxidase; Antivenins (source: MeSH NLM)
INTRODUCCIÓN
El Perú, por ser un país amazónico, debe afrontar los retos de una fauna rica y variada, en la que se encuentran los animales ponzoñosos; siendo las serpientes las que generan la máxima atención de los especialistas en salud (1,2). Las dos grandes familias de serpientes venenosas que habitan en nuestro país son las Familias Viperidae y Elapidae. Dentro de la familia Viperidae están la mayoría de las serpientes llamadas comúnmente víboras cuyo veneno posee acción hemorrágica proteolítica y necrozante; mientras que en la familia Elapidae se encuentran las llamadas corales, serpientes con venenos neurotóxicos (3,4).
No existiendo una estadística prolija sobre los accidentes ofídicos ya que muchos de ellos no son reportados (5,6), es claro que las poblaciones más expuestas son las comunidades nativas, los trabajadores eventuales en agricultura y minería y los turistas. Por tanto el requerimiento de antivenenos o sueros antiofídicos se hace cada vez más apremiante. El Instituto Nacional de Salud (INS) es el único centro especializado que produce este tipo de compuesto biológico obtenido por hiperinmunización de caballos con el o los venenos que deben ser neutralizados (7).
Teniendo en cuenta que junto con las enzimas proteolíticas, la hialuronidasa (HA), la L aminoácido oxidasa (LAO) y la fosfolipasa A2 (PLA2), son enzimas relacionadas con el envenenamiento (8-11). la inhibición que puede ejercer el antiveneno sobre alguna de ellas puede servir como indicador de la capacidad neutralizante del antiveneno utilizado.
Con este propósito, en la presente investigación colaborativa se han examinado los cambios en la actividad enzimática de HA, LAO y PLA, enzimas relacionadas con la difusión del veneno, los procesos inflamatorios y la miotoxicidad respectivamente, al interactuar in vitro con el antiveneno botrópico polivalente
MATERIALES Y MÉTODOS
INSUMOS
Venenos
Los venenos en estudio procedieron del Serpentario del INS y correspondieron a las siguientes especies: Bothrops atrox Jergón, Bothrops brazili Jergón shushupe, Bothrops pictus Jergón de costa y Bothrops barnetti Macanche.
Antivenenos
El antiveneno botrópico polivalente al estado líquido, fue proporcionado por el INS en ampollas de 10 mL y contiene anticuerpos equinos contra los venenos mencionados (Lote 300143). En cuanto al antiveneno botrópico polivalente sometido a calentamiento por cinco días a 37 ºC correspondió al lote: 01000376.
DETERMINACIÓN DE PROTEÍNAS
El contenido proteico de cada veneno fue determinado tanto por la absorbancia en el rango de luz ultravioleta a 280 nm, en un espectrofotómetro UV/visible Shimadzu (12). Así mismo se determinó también el contenido proteico por el método de Lowry et al. (13), modificado en este laboratorio (14). Para ello se utilizó una solución alcalina, el reactivo de Folin Ciocalteus y la albúmina sérica bovina como proteína estándar, midiéndose la absorbancia a 660 nm. Para el caso de los antivenenos, la cantidad de proteína fue determinada por el método cuantitativo de Biuret, utilizándose solución de sulfato de cobre 0,15%, tartrato de sodio y potasio 0,6% e hidróxido de sodio 3%, midiéndose la densidad óptica a 450 nm (15).
ACTIVIDADES ENZIMÁTICAS
Actividad de fosfolipasa A2. Se empleó como sustrato 1,5 mL de una emulsión lipoproteica de yema de huevo al 45% en buffer Tris-HCl 10 mM con CaCl2 10 mM a pH 7,4. Luego de adicionar 100 μL de muestra, es decir, veneno o inmunocomplejo (veneno + antiveneno), se incubó por 10 minutos a 37 ºC. Después las mezclas fueron sometidas a un baño de temperatura a 100 ºC, midiéndose el retardo del tiempo de coagulación de la yema de huevo (10,11).
Actividad de L-aminoácido oxidasa. Se empleó como sustrato L-leucina 0,1 M en buffer Tris-imidazol 0,2 M pH 7,6 que contenía 0,0065% de O-dianisidina. La mezcla de reacción conteniendo 2,9 mL de sustrato y 0,1 mL de peroxidasa comercial al 0,001% fue preincubada a 37 ºC por cinco minutos, luego se añadió 0,1 mL de muestra registrándose el incremento en la absorbancia a 436 nm (9,16).
Actividad de hialuronidasa. La actividad de esta enzima fue determinada usando como sustrato ácido hialurónico 0,5 mg/mL en buffer acetato de amonio 0,05 M, NaCl 0,15 M, pH 5,0. Luego de agregar la muestra, se determinó la reducción de la turbidez del sustrato por acción enzimática al adicionarse el reactivo de turbidez bromuro cetil trimetil amonio (BCTA), realizándose la medición espectrofotométrica a 400 nm (17,18).
Preparación del antiveneno calentado. Los viales del antiveneno botrópico polivalente (Lote 01000376) fueron colocados a temperatura ambiente por 60 minutos y luego sometidos a calentamiento de 37 °C por cinco días en una estufa incubadora (Gallenkamp). Posteriormente se verificó la homogeneidad del producto antes de utilizarlo.
Ensayos de neutralización. Se prepararon mezclas que contenían cantidades equivalentes a ½, 1 y 2 dosis neutralizantes del antiveneno con cada uno de los venenos en estudio. Para ello se usó como referencia la valoración de la etiqueta del antiveneno que indica que una dosis neutralizante corresponde a 10 mL del antiveneno contra 25 mg del veneno de B. atrox. Luego de 30 minutos de incubación a 37 °C se ensayaron los valores de actividad enzimática por triplicado con respecto a una muestra control de veneno. Se consideró que un antiveneno fue eficaz si una dosis neutralizante logró reducir a menos del 50% la actividad enzimática respectiva.
RESULTADOS
CONTENIDO DE PROTEÍNA EN VENENOS Y ANTIVENENOS
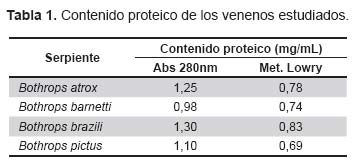
La Tabla 1 muestra los valores de proteína de los venenos en estudio tanto por absorbancia a 280 nm como por el método de Lowry. Se puede apreciar que los valores UV 280, considerando 1 mg de veneno, son mayores en la mayoría de los casos a una unidad óptica; en cambio, por el método de Lowry los valores corresponden a cantidades de proteínas menores al peso del veneno y pueden fácilmente extrapolarse a porcentaje de proteína en cada muestra. Así tenemos que para B. atrox es de 78%, B pictus 69%, B. brazili 83 %, y para B. barnetti es de 74%.
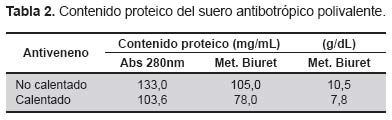
La Tabla 2 muestra los valores de proteína tanto por absorbancia a 280 nm como por el método de Biuret de los antivenenos en estudio. Al igual que los venenos, los antivenenos muestran una mayor concentración relativa por luz UV que por el método de Biuret.
INHIBICIÓN DE LAS ENZIMAS EN ESTUDIO
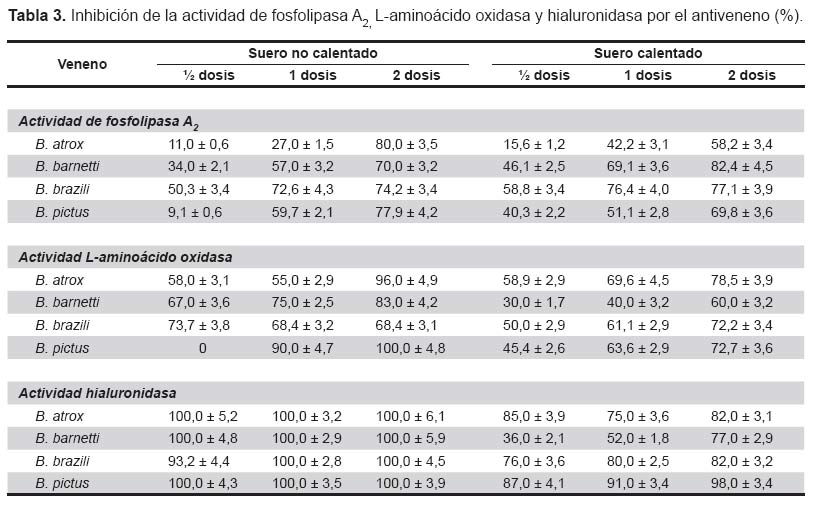
Acción del antiveneno sobre PLA2. La tabla 3 muestra que usando dos dosis del antiveneno, se obtuvo una inhibición entre 70 a 80% de la actividad de PLA2, mientras que a dosis menores, también se reducen los valores de inhibición. Con respecto al suero calentado a 37 °C por cinco días, se observa una reducción del efecto inhibitorio de este antiveneno sobre la actividad de la enzima ya que, a dos dosis los valores de inhibición estuvieron entre 58,2 y 82,4%. Nótese que el veneno más afectado por la inhibición fue el de B. atrox (dos dosis) con un valor de 80% y el de B. barnetti (dos dosis) con un valor de 82,4%.
Acción del antiveneno sobra la actividad de LAO. Los venenos de B. pictus y B. atrox fueron los más severamente inhibidos por el antiveneno no calentado con valores de inhibición de 100 y 96% respectivamente usando dos dosis del suero. Siendo el mínimo valor de inhibición (68%) el encontrado para B. brazili (Tabla 3).
Observando los datos obtenidos con el antiveneno calentado a 37 °C, nuevamente encontramos una reducción en la capacidad inhibitoria del antiveneno ya que, por ejemplo a dos dosis el valor registrado con el veneno de B. atrox fue de 78.5% en tanto que, para el veneno de B. barnetti se obtuvo un porcentaje de 60%.
Acción del antiveneno sobre la actividad de HA. Los datos registrados en la tabla 3, señalan claramente que el antiveneno botrópico polivalente no calentado a cualquier dosis utilizada produjo una inhibición total de la actividad hialuronidasa, con excepción del veneno de B. brazili que registró un valor de 93,2% usando ½ dosis del suero. En cuanto al antiveneno calentado a 37 °C por cinco días, se obtuvieron valores de inhibición menores, aún cuando a dos dosis del suero todos los venenos fueron inhibidos en la actividad de HA.
DISCUSIÓN
Como se ha demostrado en numerosos reportes, los venenos ofídicos son complejos proteicos en los que predominan las enzimas (Familia Viperidae) o las toxinas (Familia Elapidae). Siendo la mayoría de serpientes peruanas especies de la familia Viperidae y teniendo en cuenta que B. atrox es la causante del mayor número de accidentes registrados (6), es conveniente considerar que el envenenamiento por esta serpiente es un proceso sumamente complicado en el que participan las enzimas contenidas en la ponzoña, las cuales afectan en diferente grado los distintos componentes biológicos de la víctima (19,20). En este sentido, es preciso conocer la actividad enzimática de las proteínas implicadas en cada veneno y deducir a base de los efectos biológicos, la acción que ellas ejercen en los diferentes órganos, tejidos y fluido sanguíneo.
Así tenemos que HA es la enzima responsable de la difusión del veneno tal como lo han demostrado numerosos autores entre otros, un reciente reporte para el veneno de la serpiente peruana Lachesis muta (8) en el que se demuestra la purificación y acción biológica de esta enzima. Esta enzima es una glicoproteína alcalina de alto peso molecular que ataca exclusivamente el ácido hialurónico y que permite la difusión continua y sostenida del veneno a través de los tejidos (21,22).
Así mismo, LAO es una enzima que si bien es cierto in vitro sólo produce la desaminación oxidativa de los L- aminoácidos generando con ello peróxido de hidrógeno, amoníaco y α- cetoácido, in vivo muestra diferentes acciones dependiendo del tipo de veneno, así tenemos que puede estar implicada en el proceso edemático (23), en la inducción (24) o inhibición de la agregación plaquetaria (25), así como en el fenómeno de apoptosis o muerte celular (26).
Por otro lado, PLA2 es una enzima mucho más variada que LAO en su acción biológica, ya que en algunos venenos se comporta como una neurotoxina tal como la crotoxina de Crotalus durissus terrificus (27), y en otros como cardiotoxina, miotoxina (28) o una proteína anticoagulante (29). Empero, su actividad sobre fosfatidilcolina y otros sustratos análogos en los ensayos in vitro es relativamente semejante.
Los resultados obtenidos en esta investigación muestran en forma diáfana que el antiveneno botrópico polivalente al estado líquido no calentado, tiene una fuerte capacidad inhibitoria de las tres enzimas. En el caso de PLA2 y LAO (Tabla 3), la inhibición alcanzada siendo muy alta no llega al 100% y es dependiente de las dosis del antiveneno usado. En cambio al ensayarse el efecto del antiveneno sobre HA (Tabla 3), la inhibición fue total con excepción de lo observado para el veneno de B. brazili aún cuando el valor obtenido fue muy alto, es decir 93,2%. Nótese también que someter al antiveneno al calentamiento exprofeso por cinco días a 37 °C origino una reducción en la capacidad inhibitoria del suero antibotrópico.
Estos datos permitirían considerar a HA como un posible indicador de la potencia del antiveneno sobre todo teniendo en cuenta que la prueba enzimática es rápida y sencilla. Además, los efectos marcados sobre las actividades de LAO y PLA2 señalan la eficacia de este producto y refuerzan el concepto de calidad expresada en varios reportes (7,30). Por último la exposición del suero a temperaturas mayores a 25 °C como por ejemplo 37 °C rango térmico que es usual en nuestra amazonía señala que la falta de cadena de frío si afecta al antiveneno al estado líquido aún cuando no anula su potencia antitóxica.
Concluimos de esta investigación que un dosaje de las tres enzimas estudiadas, pero principalmente HA, podría servir como indicador in vitro de la potencia del antiveneno mediante un ensayo rápido de laboratorio y además que el antiveneno producido por el INS guarda las condiciones in vitro de inhibición de tres de las principales actividades de los venenos de serpientes peruanas, implicadas en la difusión de la ponzoña así como en sus efectos sistémicos.
REFERENCIAS BIBLIOGRÁFICAS
1. Campbell JA, Lamar WM. The venomous reptiles of Latin America. New York: Comstock Publishing; 1989.
2. Carrillo N. Contribución al conocimiento de las serpientes venenosas del Perú de las familias Viperidae, Elapidae e Hydrophiidae. Publ Mus Hist Nat UNMSM. 1983; (A, Zool) 49: 1-27.
3. Tu AT. Venoms: chemistry and molecular biology. New York: Wiley & Sons; 1977.
4. Meneses O. Los animales venenosos y sus peligros. Lima: Instituto de Salud Pública; 1974.
5. Zavaleta A. Mordedura de serpiente (ofidismo): un problema de salud en el Perú. Rev Med Hered. 2004; 15(2): 61-63.
6. Villanueva M, Maguiña C, Cabada M, De Marini J, Alvarez H, Gotuzzo E. Ofidismo en la provincia de Chanchamayo, Junín: revisión de 170 casos consecutivos en el Hospital de Apoyo de La Merced. Rev Med Hered. 15(2): 82-87.
7. Laing GD, Yarlequé A, Marcelo A, Rodríguez E, Warrell DA, Theakston RGD. Preclinical testing of three South American antivenoms against the venoms of five medically-important Peruvian snake venoms. Toxicon. 2004; 44(1): 103-6.
8. H urtado L, Lerma L, Rodríguez E, Yarlequé A. Aislamiento y algunas propiedades bioquímicas de una hialuronato glucanohidrolasa del veneno de la serpiente Lachesis muta Shushupe. Rev Soc Quim Peru. 2007; 73(4): 226-34.
9. Lazo F, Málaga O, Yarlequé A, Severino R, Gutierrez, S. Actividad antimicrobiana de una flavoproteína aislada del veneno de la serpiente peruana Bothrops atrox (jergón). Rev Soc Quim Peru. 2007; 73(4): 197-207.
10. Mejia J, Inga R, Lazo F, Rodriguez E, Yarleque A, Zavaleta A. Purificación y propiedades bioquímicas de una Fosfolipasa A del veneno de la serpiente Lachesis muta Shusupe. Rev Soc Quim Peru. 2006; 75(2): 86-95.
11. Vidal JC, Stoppani AO. Isolation and purification of two phospholipases A from Bothrops venoms. Arch. Biochem.Biophys. 1971; 145(2): 543-55.
12. Warburg O, Christian W. [Isolation adn crystallization of the glycolytic enzyme enolase]. Biochemische Zeitschrift. 1941; 31: 384-421. [alemán]
13. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the folin phenol reagent. J Biol Chem. 1951; 193: 265-75.
14. Loayza S, Morante Y, Campos S, Yarlequé A. Enzimas proteolíticas en el veneno de las serpientes peruanas Lachesis muta y Bothrops atrox. Bol Soc Quím Peru. 1985; 52(3): 151-63.
15. Gornall AG, Bardawill CJ, David MM. Determination of serum proteins by means of the biuret reaction. J Biol Chem. 1949; 177(2):751-66.
16. Solis Ch, Escobar E, Yarleque A, Gutiérrez S. Purificación y caracterización de la L-aminoácido oxidasa del veneno de la serpiente Bothrops brazili Jergón Shushupe. Rev Peru Biol. 1999; 6(1): 75-84.
17. Di Ferrante N. Turbidimetric measurement of acid mucopolysaccharides and hyaluronidase activity. J Biol Chem. 1956; 220(1): 303-6.
18. H urtado L. Aislamiento y propiedades bioquímicas de una hialuronidasa del veneno de la serpiente Lachesis muta shushupe. [Tesis para optar el título profesional de Biólogo]. Lima: Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos;1997.
19. Yarlequé A. Las serpientes peruanas y sus venenos. Lima: Fondo Editorial, Universidad Nacional Mayor de San Marcos; 2000.
20. Du XY, Clemetson KJ. Snake venom L-amino acid oxidases. Toxicon. 2002; 40(6): 659-65.
21. Girish KS, Shashidharamurthy R, Nagaraju S, Gowda TV, Kemparaju K. Isolation and characterization of hyaluronidase a spreading factor from Indian cobra (Naja naja) venom. Biochimie. 2004; 86(3): 193-202.
22. Kudo K, Tu AT. Characterization of hyaluronidase isolated from Agkistrodon contortrix contortrix (Southern Copperhead) venom. Arch Biochem Biophys. 2001; 386(2): 154-62
23. A li SA, Stoeva S, Abbasi A, Alam JM, Kayes R, Faigle M, et al. Isolation, structural, and functional characterization of an apotosisinducing L-amino acid oxidase from leaf-nosed viper (Eristocophis macmahoni) snake venom. Arch Biochem Biophys. 2000; 384(2): 216-26.
24. Li ZY, Yu TF, Lian E. Purification and characterization of L-amino acid oxidase from king cobra (Ophiophagus hannah) venom and its effects on human platelet aggregation. Toxicon. 1994; 32(11): 1349- 58.
25. Takatsuka H, Sakurai Y, Yoshioka A, Kokubo T, Usami Y, Suzuki M, et al. Molecular characterization of L-amino acid oxidase from Agkistrodon halys blomhoffii with special reference to platelet aggregation. Biochim Biophys Acta. 2001; 1544(1-2): 267-77.
26. Souza DH, Eugenio LM, Fletcher JE, Jiang MS, Garratt RC, Oliva G, et al. Isolation and structural characterization of a cytotoxic L-amino acid oxidase from Agkistrodon contortrix laticinctus snake venom: preliminary crystallografic data. Arch Biochem Biophys. 1999; 368(2): 285-90.
27 B on C, Bouchier C, Choumet V, Faure G, Jiang MS, Lambezat MP, et al. Crotoxin, half-century of investigations on a phospholipase A2 neurotoxin. Acta Physiol Pharmacol Latinoam. 1989; 39(4): 439- 48.
28. Gutiérrez JM, Ponce-Soto LA, Marangoni S, Lomonte B. Systemic and local myotoxicity induced by snake venom group II phospholipases A2: comparison between crotoxin, crotoxin B and a Lys49 PLA2 homologue. Toxicon. 2008; 51(1): 80-92.
29. Carredano E, Westerlund B, Persson B, Saarinen M, Ramaswamy S, Eaker D, et al. The three-dimensional structures of two toxins from snake venom throw light on the anticoagulant and neurotoxic sites of phospholipase A2. Toxicon. 1998; 36(1): 75-92.
30. Rojas E, Quesada L, Arce V, Lomonte B, Rojas G, Gutiérrez JM. Neutralization of four Peruvian Bothrops sp. snake venoms by polyvalent antivenoms produced in Perú and Costa Rica: preclinical assessment. Acta Trop. 2005; 93(1): 85-95.
Recibido: 27-03-08
Aceptado: 18-06-08

