










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.30 no.2 Lima abr. 2013
Original Breve
Metalo-ß-lactamasas en aislamientos clínicos de pseudomonas aeruginosa en Lima, Perú
Metalo-ß-lactamases in clinical isolates of pseudomonas aeruginosa in Lima, Peru
Edgar Gonzales-Escalante1,a, William Vicente-Taboada2,b, Roky Champi-Merino3,a,Javier Soto-Pastrana4,a, Wilfredo Flores-Paredes5,b, Margarita Lovera-García5,a, Nancy Chuquiray-Valverde6,a, Carlos Bejarano-Cristobal6,a, Maritza Puray-Chávez 7,a, Segundo León-Sandoval7,a,c
1 Instituto Nacional de Salud del Niño. Lima, Perú.
2 Instituto Nacional de Enfermedades Neoplásicas. Lima, Perú.
3 Hospital Nacional Hipólito Unanue. Lima, Perú.
4 Hospital Nacional Docente Madre Niño San Bartolomé. Lima, Perú.
5 Hospital Nacional Guillermo Almenara Irigoyen. Lima, Perú.
6 Hospital Alberto Sabogal Sologuren. Lima, Perú.
7 Laboratorio de Epidemiología Molecular y Genética, Instituto de Medicina Tropical, Universidad Nacional Mayor de San Marcos. Lima, Perú.
a Tecnólogo médico; b médico patólogo clínico; c magíster en Enfermedades Infecciosas y Tropicales
RESUMEN
Con el objetivo de detectar y caracterizar molecularmente las metalo-ß-lactamasas (MßL) en aislamientos clínicos de Pseudomonas aeruginosa, se realizó un estudio trasversal en seis hospitales de referencia de Lima (Perú) en agosto de 2011. Se evaluó 51 aislamientos de P. aeruginosa, resistentes a ceftazidima y con sensibilidad reducida a carbapenémicos. El ensayo fenotípico se realizó con el método de aproximación de discos con sustratos (ceftazidima, imipenem y meropenem) y con ácido etilendiaminotetraacético (EDTA). La detección de genes MßL se realizó mediante la técnica de reacción en cadena de polimerasa multiplex. A través del método fenotípico se detectaron MßL en el 15,7% de los aislamientos, en todos ellos la detección de genes mostró la presencia del gen blaIMP. La descripción del primer reporte de MßL en aislamientos de P. aeruginosa en el Perú debería alertar a los equipos de vigilancia epidemiológica intrahospitalaria para promover su control y prevenir su diseminación.
Palabras clave: Farmacorresistencia bacteriana; Penicilinasa; Genes MDR; Proteínas asociadas a resistencia a múltiples medicamentos; Pseudomonas aeruginosa (fuente: DeCS BIREME).
ABSTRACT
The aim of this study was to detect and characterize molecularly metallo-ß-lactamase (MßL) in clinical isolates of Pseudomonas aeruginosa. We carry out a cross sectional study in six publics hospital in Lima on August 2011. 51 isolates of P. aeruginosa resistant to ceftazidime and reduced susceptibility to carbapenemes were evaluated.
The phenotypic assay was performed using the approximation method with substrate disks (ceftazidime, imipenem and meropenem) and ethylenediaminetetraacetic acid (EDTA). MßL gene detection was performed using the technique of polymerase chain reaction (PCR) multiplex. Through MßL detected phenotypic method in 15.7% of isolates. Detection of genes revealed the presence of the gene in the 8 isolates blaIMP. The first report of MßL in P. aeruginosa in Peru was described, this should alert the monitoring equipment in the institutions to promote control their spread.
Key words: Drug resistance, bacterial; Penicillinase; Genes, MDR; ltidrug resistance-associated proteins; Pseudomonas aeruginosa (source: MeSH NLM).
INTRODUCCIÓN
La Pseudomonas aeruginosa, es el patógeno humano más importante del género Pseudomonas, caracterizado por su alta virulencia y morbimortalidad. Tradicionalmente considerado como un patógeno oportunista asociado a múltiples infecciones intrahospitalarias, ahora se sabe que la P. aeruginosa se encuentra asociada a infecciones adquiridas tanto en la comunidad como en ámbitos nosocomiales.
Existen pocas opciones terapéuticas efectivas para las infecciones causadas por este microorganismo, ello debido a la elevada resistencia intrínseca que este presenta ante diferentes familias de antimicrobianos (1, 2).
La farmacorresistencia de este microorganismo esta relacionado a diferentes mecanismos tales como la alteración de las bombas de eflujo o la alteración de la permeabilidad en la membrana celular. La alteración de la MexAB-OprM, que es una bomba de eflujo, compromete de manera simultánea la sensibilidad a penicilinas, cefalosporinas, quinolonas y al meropenem. Otro ejemplo es la pérdida de la porina OprD, que se encuentra en la membrana externa y es utilizada por los carbapenémicos, y no por otros ß-lactámicos, para su penetración en la célula; cuya alteración se asocia a la resistencia a imipenem y a una sensibilidad reducida a meropenem, esta pérdida es mediada por la mutación de los genes que codifican esta porina (3, 4). Sin embargo, es la adquisición de genes codificantes de metalo-ß-lactamasas (MßL) el mecanismo de resistencia bacteriana que tiene una alta implicancia epidemiológica, debido a su capacidad de diseminación horizontal.
Las MßL poseen cuatro características principales: poseen actividad contra los carbapenemes; no hidrolizan los monobáctamicos, como el aztreonam; son inhibidas por quelantes, como el ácido etilendiaminotetraacético (EDTA), o el mercapto acetato de sodio, y requieren cationes divalentes (generalmente Zn +2) como cofactor para su actividad catalítica; además de ello, las MßL no son inhibidas por los antibióticos suicidas (ácido clavulánico, sulbactan y tazobactan) que bloquean la acción de las serino-ß-lacamasas (5, 6). Diversas familias de MßL han sido identificadas hasta el momento y, debido a sus características bioquímicas, han sido divididas en tres subgrupos. El subgrupo 3a incluye enzimas con amplio espectro de hidrólisis para penicilinas, cefalosporinas e imipenem, muchas enzimas de este subgrupo necesitan de Zn+2 para desarrollar su actividad. El subgrupo 3b comprende enzimas que hidrolizan preferentemente carbapenemes siendo consideradas las verdaderas carbapenemasas, todas las enzimas de este subgrupo necesitan Zn+2 para su actividad y son inhibidas por el EDTA. El subgrupo 3c comprende enzimas que hidrolizan rápidamente ampicilina (7, 8).
A pesar de que en otras partes del mundo han sido ampliamente documentadas, en nuestro país los genes productores de MßL no han sido detectados hasta la fecha. La aproximación de un disco que contenga un agente quelante como el EDTA a un disco que contenga un carbapenémico podría resultar una herramienta útil para la detección de MßL; ello debido que el efecto sinérgico entre los carbapenémicos y el agente quelante sería indicativo de la presencia de MßL. Sin embargo, ningún método fenotípico ha sido suficientemente estandarizado o recomendado para la detección de MßL en el laboratorio de microbiología clínica (9, 10). El objetivo de este estudio fue detectar y caracterizar molecularmente las MßL responsables de la resistencia a carbapenémicos en aislamientos clínicos de Pseudomonas aeruginosa.
EL ESTUDIO
Se realizó un estudio trasversal, en el cual se estudiaron 51 aislamientos consecutivos y no repetitivos de P. aeruginosa provenientes de pacientes hospitalizados en seis hospitales públicos de Lima (uno pediátrico, uno materno-infantil, uno oncológico y tres generales) recolectados durante el mes de agosto de 2011. Los criterios de selección de los microorganismos fueron señalados siguiendo las recomendaciones de la Subcomisión de Antimicrobianos de la Sociedad Argentina de Bacteriología (10, 11); es decir, que durante las pruebas de disco difusión la P. aeruginosa tuviera resistencia a ceftazidima (halos menores a 15 mm) y sensibilidad reducida a los carbapenémicos (halos menores a 21 mm de imipenem o meropenem). El procesamiento de los aislamientos se realizó en el Laboratorio de Epidemiologia Molecular del Instituto de Medicina Tropical de la Universidad Nacional Mayor de San Marcos.
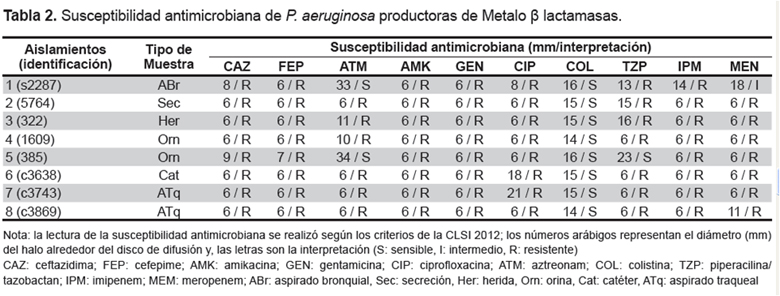
Se realizaron pruebas de sensibilidad a los antimicrobianos en los aislamientos mediante la técnica de disco difusión, de acuerdo con las recomendaciones del Clinical and Laboratory Standards Institute (CLSI). Se incluyeron discos conteniendo aztreonam 30 µg (ATM); ceftazidima 30 µg (CAZ); cefepima 30 µg (FEP); imipenem 10 µg (IPM); meropenem 10 µg (MEN); piperacilina/tazobactam 100/10 µg (TZP); amikacina 5 µg (AMK); gentamicina 10 µg (GEN); ciprofloxacina 5 µg (CIP), y colistin 10 µg (COL, Oxoid®). Inmediatamente después del tiempo de incubación, los diámetros de los halos de inhibición para cada uno de los antimicrobianos se registraron e interpretaron de acuerdo con las normas CLSI 2012 (12).
Para la determinación fenotípica de las MßL se utilizó la la técnica de aproximación del disco modificada. Para ello se inocularon las placas según las recomendaciones del CLSI para la prueba de disco difusión, y se colocaron discos conteniendo EDTA (Laboratorios Britania-Argentina: EDTA 372 µg / mercapto acetato de sodio 900 µg) y discos comerciales conteniendo imipenem 10 µg, meropenem 10 µg y ceftazidima 30 µg (Oxoid®), respetando la distancia de 1,5 cm de centro a centro entre los discos con antibióticos y el inhibidor. Las placas se incubaron durante 18 horas a 35 ºC. Un agrandamiento en el halo de inhibición del disco con antibiótico en la zona adyacente al disco con EDTA fue interpretado como posible presencia de MßL (9, 10).
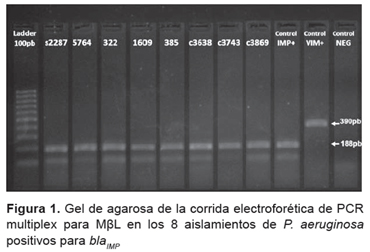
La detección genotípica de la MßL se realizó mediante un ensayo de Multiplex-PCR empleando como molde ADN total (10). Los pares de cebadores empleados se detallan en la Tabla 1.

Los reactivos empleados fueron de Invitrogen (EE. UU.). Se empleó un termociclador VERITI Applied Byosistem (EE. UU.) y la siguiente reacción de amplificación: desnaturalización 94 °C, 5 min; seguida de 36 ciclos de: desnaturalización 94 °C, 30 s; hibridación 52 °C, 40 s; amplificación 72 °C, 50 s; y un período final de elongación 72 °C, 5 min. Los fragmentos esperados fueron IMP (188pb), VIM (390pb), SPM (271pb), GIM (477pb) y SIM (570pb).
HALLAZGOS
A través del método fenotípico se detectaron aislamientos sospechosos de producir MßL en el 15,7% (8/51) de los aislamientos, en ellos se observó sinergia entre los carbapenémicos y el disco que contenía EDTA. El 25% (2/8) de los aislamientos sospechosos de producir MßL fueron sensibles al aztreonam, a pesar de ser este un indicador de la presencia de las MßL. En todos los casos los aislamientos fueron sensibles a colistin. Los datos de susceptibilidad se muestran en la Tabla 2. La detección de los genes codificantes de MßL por Multiplex-PCR reveló la presencia del gen bla IMP en los ocho aislamientos sospechosos de producir MßL que fueran detectados por el ensayo fenotípico empleando EDTA (Figura 1), lo que indica que los ensayos de detección fenotípica resultaron adecuados.
DISCUSIÓN
Si bien los resultados obtenidos pertenecen a un periodo corto y la situación epidemiológica es particular para las distintas instituciones del país, es importante notar que la prevalencia de MßL en aislamientos de P. aeruginosa resistentes ceftazidima y con sensibilidad reducida a carbapenémicos resultó ser similar a la de otros estudios realizados en América Latina. En Argentina, dos estudios han señalado frecuencias de 11 y 14%, en ambos casos se aisló la enzima VIM (13, 14); en Chile se encontró una frecuencia de 18,6% de MßL también del tipo VIM (15). En tanto que en Brasil se ha señalado que el gen prevalente es SPM-1 con frecuencias que varían entre 5 a 62%, en diferentes regiones del país, en el caso del gen IMP la prevalencia va de 2 a 8% y, finalmente, el gen VIM fue detectado con una prevalencia del 30% (16).
La no detección de genes para MßL en 84,3% de los aislamientos de P. aeruginosa resistentes a ceftazidima y con sensibilidad reducida a los carbapenémicos, asociado al resultado de la prueba fenotípica refuerza la hipótesis que la producción de MßL no es el principal mecanismo de resistencia a los carbapenémicos en P. aeruginosa. Este resultado sugiere la ocurrencia de otros mecanismos de resistencia, como mutaciones en el gen OprD, que resultan en porinas con reducida afinidad por el imipenem y es considerado el principal responsable por la resistencia al imipenem en P. aeruginosa (17). La resistencia al meropenem puede ser resultado de la hiperexpresión de sistemas de eflujo (18).
Si bien las MßL no hidrolizan eficientemente al aztreonam, por lo cual no son capaces de conferir resistencia a este antibiótico, en esta investigación se obtuvieron dos aislamientos sensibles, los demás valores se interpretaron como resistentes; por lo que no resulto un buen indicador de la presencia de MßL; sin embargo, la resistencia a dicho agente no descarta su presencia (5, 6). En este caso, la resistencia a este antibiótico estaría mediada por otro mecanismo de resistencia no analizado en el presente estudio, como por ejemplo, la presencia de ß-lactamasas de espectro extendido o sobre expresión de un sistema de eflujo (19, 20). En todos los casos se observo sensibilidad a colistin, siendo este una buena alternativa terapéutica en caso de P. aeruginosa multirresistente.
Algunas limitaciones de nuestro estudio deben ser reconocidas, el tiempo en que se recolectaron las muestras y su cantidad no permiten determinar la magnitud del problema, por lo que hacen faltan estudios mayores para conocer la realidad nacional en torno a la resistencia bacteriana mediada por MßL en P. aeruginosa. Además, es necesaria la realización del secuenciamiento para conocer las variantes alélicas de esta enzima y su codificación genética que nos permita conocer si existen otros genes de resistencia relacionados, como enzimas modificadoras de aminoglucósidos.
En conclusión, se ha descrito el primer reporte de MßL en P. aeruginosa en el Perú, la descripción molecular de estas enzimas ha señalado que el gen blaIMPestuvo presente en todos ellas. El desarrollo de este mecanismo de farmacorresistencia en el Perú podría tener un grave impacto en el ámbito clínico y epidemiológico, en particular de los hospitales incluidos en el presente estudio; por lo cual los equipos de vigilancia de estos hospitales deberían promover medidas de control para evitar su diseminación.
Agradecimientos: al personal del Laboratorio de Epidemiología Molecular y Genética del Instituto de Medicina Tropical de la Universidad Nacional Mayor de San Marcos, por el apoyo brindado para la realización de esta investigación.
Contribuciones de autoría: EGE, WVT, RCM, JSP y WFP han participado en la concepción del artículo, la recolección de datos, material de estudio y redacción del artículo. MPC y SLS participaron en la idea de la investigación, la redacción y la asesoría técnica y administrativa. Además, MLG, NCV Y CBC participaron en la recolección de datos y material de estudio y redacción del artículo. Todos los autores aprobaron la versión final del manuscrito.
Fuentes de financiamiento: autofinanciada
Conflictos de interés: los autores declaran no tener conflicto de interés.
Referencias Bibliográficas
1. Classics in infectious diseases. On the blue and green coloration that appears on bandages. By Carle Gessard (1850-1925). Rev Infect Dis. 1984;6 Suppl 3:S775-6
2. Gales AC, Jones RN, Turnidge J, Rennie R, Ramphal R. Characterization of Pseudomonas aeruginosa isolates: occurrence rates, antimicrobial susceptibility patterns, and molecular typing in the global SENTRY Antimicrobial Surveillance Program, 1997-1999. Clin Infect Dis. 2001;32(Suppl 2):S146-55. [ Links ]
3. Livermore DM. Of Pseudomonas, porins, pumps and carbapenems . J Antimicrob Chemother. 2001;47(3):247-50. [ Links ]
4. Livermore DM, Woodford N. Carbapenemases: a problem in waiting? Curr Opin Microbiol. 2000;3(5):489-95. [ Links ]
5. Walsh T, Toleman M, Poirel L, Nordmann P. Metallo-ß-lactamases: the quiet before the storm? Clin Microbiol Rev. 2005;18(2):306-25. [ Links ]
6. Queenan A, Bush K. Carbapenemases: the versatile ß-lactamases . Clin Microbiol Rev. 2007;20(3):440-58. [ Links ]
7. Rasmussen BA, Bush K. Carbapenem-hydrolyzing beta-lactamases . Antimicrob Agents Chemother. 1997;41(2):223-32. [ Links ]
8. Bebrone C. Metallo-ß-lactamases (classification, activity, genetic organization, structure, zinc coordination) and their superfamily . Biochem Pharmacol. 2007;74(12):1686-1701. [ Links ]
9. Arakawa Y, Shibata N, Shibayama K, Kurokawa H, Yagi T, Fujiwara H, et al. Convenient test for screening metallo- b-lactamase-producing gram-negative bacteria by using thiol compounds . J Clin Microbiol. 2000;38(1):40-3. [ Links ]
10. Radice M, Marín M, GiovanakisM, Vay C, AlmuzaraM, LimanskyA, et al. Criterios de ensayo, interpretación e informe de las pruebas de sensibilidad a los antibióticos en los bacilos gram negativos no fermentadores de importancia clínica: recomendaciones de la Subcomisión de Antimicrobianos de la Sociedad Argentina de Bacteriología, Micología y Parasitología Clínicas, Asociación Argentina de Microbiología . Rev Argent Microbiol. 2011;43(2):136-53. [ Links ]
11. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing. Twenty-second informational supplement . M100-S22. Wayne PA: Clinical and Laboratory Standards Institute; 2012. [ Links ]
12. Ellington M, Kistler J, Livermore D, Woodford N. Multiplex PCR for rapid detection of genes encoding acquired metallo-beta-lactamases . J Antimicrob Chemother. 2007;59(2):321-2. [ Links ]
13. Pagniez G, Radice M, Cuirolo A, Rodríguez O, Rodríguez H, Vay C, et al. Prevalencia de metalo-ß-lactamasas en Pseudomonas aeruginosa resistentes a carbapenemes en un hospital universitario de Buenos Aires . Rev Argent Microbiol. 2006;38(1):33-7. [ Links ]
14. Cejas D, Almuzara M, Santella G, Tuduri A, Palombarani S, Figueroa S, et al. Caracterización fenotípica y genotípica de la resistencia a imipenem en Pseudomonas aeruginosa aisladas en un hospital de Buenos Aires . Rev Argent Microbiol. 2008;40:238-245
15. Pérez A, García P, Poggi H, Braun S, Castillo C, Román J, et al. Presencia de metalo-ß-lactamasas en Pseudomonas aeruginosa resistente a imipenem . Rev Méd Chile. 2008;136:423-32. [ Links ]
16. Sader HS, Castanheira M, Mendes RE, Toleman M, Walsh TR, Jones RN. Dissemination and diversity of metallo-b-lactamases in Latin America: report from the SENTRY Antimicrobial Surveillance Program . Int J Antimicrob Agents. 2005;25(1):57-61. [ Links ]
17. Farra A, Islam S, Stralfors A, Sorberg M, Wretlind B. Role of outer membrane protein OprD and penicillin-binding proteins in resistance of Pseudomonas aeruginosa to imipenem and meropenem . Int J of Antimicrob Agents. 2008;31(5):427-33. [ Links ]
18. Pages JM, Amaral L. Mechanisms of drug efflux and strategies to combat them: challenging the efflux pump of Gram-negative bacteria . Biochim Biophys Acta. 2009;1794(5):826-33. [ Links ]
19. Schweizer HP. Efflux as a mechanism of resistance to antimicrobials in Pseudomonas aeruginosa and related bacteria: unanswered questions . Genet Mol Res. 2003;2(1):48-62. [ Links ]
20. Bush K, Jacoby GA. Updated functional classification of ß-lactamases . Antimicrob. Agents. Chemother. 2010;54(3):969-76. [ Links ]
Correspondencia: Edgar Gonzales Escalante
Dirección: Av. Brasil 600, Lima 05, Perú.
Teléfono: (511) 330-0066 anexo 3201.
Correo electrónico: egones_5@hotmail.com
Recibido: 19-12-12
Aprobado: 17-04-13
