










Servicios Personalizados
Revista

Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
versión impresa ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.37 no.1 Lima ene./mar 2020
http://dx.doi.org/10.17843/rpmesp.2020.371.4745
Brief report
Nitrofuran resistance in Salmonella enterica isolated from meat for human consumption
1 Instituto de Salud Global, Hospital Clinic - Universitat de Barcelona, Barcelona, España.
2 Laboratorio de Microbiología Molecular y Genómica Bacteriana, Universidad Científica del Sur, Lima, Perú.
4 Instituto de Medicina Tropical Alexander von Humboldt, Universidad Peruana Cayetano Heredia, Lima, Perú.
5 Department of Epidemiology, School of Public Health, University of Texas Health Science Center at Houston, Estados Unidos.
INTRODUCTION
Nitrofurans are a group of synthetic antimicrobials which are used against Gram-negative and Gram-positive parasites and bacteria 1. These compounds have been widely used in both human and veterinary medicine, as well as in growth promoters for animals for human consumption (1. However, their use currently in veterinary medicine regarding animals for human consumption is prohibited in many countries because of the risk that these antimicrobials or their metabolites remain in the food 1. Its veterinary use is prohibited in Peru since 2013 (2.
The levels of nitrofuran resistance in Enterobacteriaceae (Shigella spp.), cause of diarrhea, are low 3, and the highest levels of resistance to these antimicrobials were reported in Salmonella spp.4,5. Currently, the mechanisms of action of nitrofurans are poorly studied, they have been described mainly in mutants obtained in vitro.
It has been described that nitrofurans need to be activated by nitroreductases to exert their action; the reduction reaction consists of several sequential steps, some of the products generated in the intermediate steps could possess antimicrobial activity 6,7. The nitroreductases involved in these activation processes are NfsA and NfsB 6. Thus, in studies with mutants obtained in vitro and in studies developed with clinical isolates 8-10, in Escherichia coli the presence of alterations capable of reducing or eliminating the functionality of NfsA and NfsB has been associated with the development of resistance to nitrofurans. The role of efflux pumps in resistance to nitrofurans has also been described. In addition, the OqxAB efflex pumps, which belongs to the RND (Resistan-ce-Nodulation-Division) family, confers resistance to nitrofurantoin 11.
Despite the high levels of resistance in Salmonella spp. the number of studies focusing on the mechanisms of resistance to nitrofurans is low 5,12. Therefore, the objective of this study was to determine the levels and mechanisms of resistance to nitrofuran in isolates of Salmonella enterica from meat samples acquired in traditional markets in Lima.
KEY MESSAGES
Motivation for the study: Increasing levels of antibiotic resistance in Peru is a serious concern. However, there is a scarcity of data regarding nitrofuran resistance in Salmonella spp.
Main findings: There are high levels of nitrofuran resistance in Salmonella spp. related with the presence of chromosomal mutations in the cnr and snrA genes.
Implications: Results show the need for systematic investigations of resistance levels to nitrofurans in Salmonella spp. mainly aimed at the causes of natural selection of resistant isolates.
THE STUDY
18 samples with Salmonella enterica isolated in 2012 from a previous study were used to determine the presence of Enterobacteriaceae and levels of E. coli antimicrobial resistance in meat samples (pork, chicken or calf) acquired in traditional markets in the north (Comas, San Martin), center (La Victoria, Cercado de Lima) and south (Villa El Salvador) of Lima 13. The study was conducted at ISGlobal, Hospital Clinic - Universitat de Barcelona (Spain), Nutreco (Spain) and Instituto de Medicina Tropical Alexander von Humboldt (Peru). In all cases the isolates were previously identified by biochemical methods and confirmed by amplification of the invA gene13. S. enterica isolates were recovered from -80 ºC and reconfirmed prior to use by amplification and sequencing of the 16S rRNA gene14.
The serotypes of the isolates were determined by means of microarrays (Check & Trace Salmonella kit, Check-Points B.V, Wageningen, The Netherlands) following the manufacturer’s instructions. Then sensitivity to furazolidone (100 µg) and nitrofurantoin (300 µg) was determined by the disk diffusion method (BD, St. Augustine del Guadalix, Spain), as well as minimum inhibitory concentration (MIC) values by the agar dilution method, following the CLSI (Clinical and Laboratory Standards Institute) guidelines and using E. coli ATCC 25922 strain as quality control 15. In the CLSI guide there is no specific cut-off point for furazolidone, so in this case data on MIC and/or halo diameter are reported.
For both nitrofurantoin and furazolidone, MIC values were determined in presence of Phenyl-Arginine-ß-Naphthylamide (PAßN), an RND-type efflux pump inhibitor 8. For its use, PAßN was dissolved in dimethyl sulfoxide (DMSO), which is way it determined the effect of this solvent on bacterial growth.
In all the isolations, nfsA and nfsB genes were amplified by the polymerase chain reaction (PCR) technique (initial cycle of 94 ºC for five minutes, followed by 35 cycles of 94 ºC for 40 seconds each, 60 ºC for 30 seconds, 68 ºC for 40 seconds, and a final extension at 72 ºC for five minutes). The PCR was performed with the primers described by Salamanca-Pinzón et al.16) ,and was visualized in 2% agarose gels stained with SYBR Safe (Invitrogen, Carlsbad, USA). The bands obtained were recovered from the gel and purified using the Wizard SV Gel and PCR Clean Up System kit (Promega, Madison, USA). The purified products were sent to Macrogen (Seoul, South Korea) for sequencing.
Finally, the presence of transferable nitrofuran resistance mechanisms was determined by conjugation, following the previously described protocol 17. For this purpose, E. coli J53 (resistant to sodium azide) was used as the receiving strain and Mueller-Hinton agar supplemented with sodium azide (150 µg/ml) and furazolidone (16 µg/ml) as the medium for selecting the transconjugants.
RESULTS
The 18 studied isolates came from four markets, different from the three areas included in the original study (northern, central and southern cone) (Table 1), thus demonstrating a wide dissemination throughout the Lima area.
Table 1 Origin of the meat samples
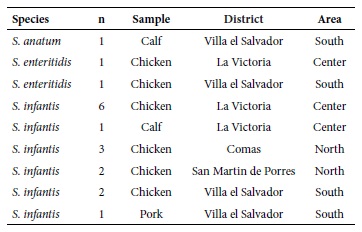
n: number of S. enterica isolates. Only one Salmonella isolation per sample was
considered.
The results of serotyping showed that most of the isolates belonged to the infantis serotype (15 isolates, 83.3%). The remaining three strains were classified as enteritidis (two strains, 11.1%) and anatum (one strain, 5.6%). The isolates of the infantis serotype were recovered from the three types of meat samples, especially from the chicken samples. Thus, 13 of the 15 isolates recovered were from chicken samples, while one of S. infantis was isolated from calf samples, and another from pork samples. The two S. enteritidis isolates were from chicken samples and the one from S. anatum was from calf (Table 1).
All isolates, except for the S. anatum strain, had MIC levels of 32-64 µg/ml for furazolidone, being resistant to nitrofurantoin with MICs of 128-256 µg/ml. The isolation of S. anatum showed MICs of 8 µg/ml for furazolidone and 32 µg/ml for nitrofurantoin (Table 2).
Table 2 Levels and mechanisms of resistance to nitrofurans in Salmonella spp.
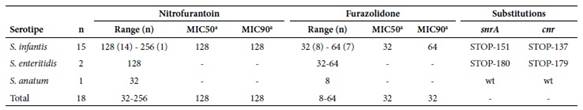
n: number of isolates; MIC: minimum inhibitory concentration; wt: without mutation.
a Calculated only for S. infantis.
Correlation between MIC values and halos observed in studies with antibiotic discs was also observed. Thus, the nitrofurantoin resistant isolates showed 8-11 mm diameter halos and 8-13 mm halos for furazolidone, while sensitive isolates showed 20 mm halos for nitrofurantoin and 24 mm halos for furazolidone.
The addition of PAßN did not affect the MIC values, which in all cases remained unchanged, showing no involvement from RND-type efflux pumps in the development of resistance to nitrofurans in the isolates studied. Neither PAßN nor DMSO interfered with the normal growth of the bacteria.
All nitrofuran-resistant isolates showed mutations in snrA and cnr. All 15 S. infantis isolates had STOP codons at position 151 of snrA and 137 of cnr, while the two S. enteritidis isolates had STOP codons at positions 180 of snrA and 179 of cnr (Table 2). Finally, no nitrofuran-resistant transconjugants were obtained in the conjugation studies.
DISCUSSION
Although the use of nitrofurans in animals for human consumption is prohibited in many countries, resistance to them has been described in enteropathogens, such as Salmonella spp. isolated from food samples 4,5; in some cases, even traces of nitrofurans have been detected in meat products 18. There are several possible explanations for these facts, which include the stability of resistance to nitrofurans, the use of these antimicrobials, despite being banned products, or the existence of environmental contamination 4,8,18.
The fact that the enteropathogens isolated originated from samples of the three types of meat (chicken, calf and pork) included in the study and their presence in the different areas of Lima, suggests the wide geographical spread in the country of S. enterica resistant to nitrofurans. However, it should be considered that the processed samples were collected in 2012, one year before Peru’s ban on the use of nitrofurans in breeding animals for consumption 2.
It has been observed that the acquisition of resistance to nitrofurans is a sequential event, in which the nitro-reductase NfsA and NfsB accumulate alterations that affect their functionality and have an additive effect on the final levels of resistance to nitrofurans 9. The study results were concordant to previous studies, detecting the presence of mutations leading to the presence of STOP codons in the equivalent genes (snrA and cnr) and the subsequent lack of functional nitroreductases. To date, very few studies have analyzed the mechanisms of resistance to nitrofurans in S. enterica; they have found similar scenarios, with the presence of STOP codons or other alterations in the genes snrA or cnr5,12.
Although no transconjugants resistant to nitrofuran were obtained in this study, transferable mechanisms of resistance to nitrofuran have been described. Among these, the OqxAB efflux pump, which has recently been implicated in the development of resistance to nitrofurantoin, deserves special attention 11. OqxAB is an RND-type efflux pump, specific to Klebsiella spp. which was first detected and coded in plasmid from E. coli of veterinary origin in a study on resistance to olaquindox 19. In Peru, the use of olaquindox in animals intended for human consumption was banned in parallel with the use of nitrofuran (2. Currently, at least 14 oqxA and 28 oqxB alleles are known to be plasmid-coded, but the specific activity of each has not been established 19.
The presence of mutations in acrB, emrD, yajR or macB, chromosomal efflux pump coding genes has been implicated in the development of resistance to nitrofurans 20. The most studied of these pumps is AcrAB which, like OqxAB, is an RND-type efflux pump. The AcrAB pump, like other RND pumps including OqxAB, can be inhibited using substances such as PAßN 19,20. In this study, the effect of PAßN was not observed, so it was ruled out that overexpression of AcrAB pumps was involved in the development of nitrofuran resistance in these isolates, providing further evidence for the absence of OqxAB plasmid pumps. In a previous study in which furazolidone-resistant E. coli mutants were developed, no involvement of PAßN-inhibitable ejection pumps was detected either 8.
Due to the methodology used, the presence of transferable resistance mechanisms which only produced low increases in resistance levels could have gone unnoticed, which is a limitation of the present study. Also, the isolates included in the study are from 2012, which highlights the need for carrying out further studies to assess the current situation.
The present study describes the presence of high levels of resistance to nitrofurans in isolates of Salmonella enterica from samples of meat foods marketed in Lima. These levels of resistance were directly related to the presence of chromosomal mutations in the snrA and cnr genes. It is necessary to keep track of the levels of resistance to nitrofurans in S. enterica.
REFERENCES
1. Vass M, Hruska K, Franek M. Nitrofuran antibiotics: a review on the application, prohibition and residual analysis. Vet Med (Praha). 2008;53(9):469-500. doi: 10.17221/1979-VETMED. [ Links ]
2. Carrasco Valiente JA. Prohíben importación y comercialización de diversos principios activos, así como el uso de los mismos en la fabricación de productos veterinarios o alimentos para animales destinados al consumo humano y establecen otras disposiciones. Resolución Directoral Nº 0072-2013-MINAGRI-SENASA-DIAIA [Internet]. Diario Oficial El Peruano; 2013 (citado el 10 de agosto de 2019): 503464-5. Disponible en: https://www.minagri.gob.pe/portal/download/pdf/marcolegal/normaslegales/resolucionesdirectorales/2013/setiembre/rd72-2013-minagri-senasa-diaia.pdf. [ Links ]
3. Pons MJ, Gomes C, Martínez-Puchol S, Ruiz L, Mensa L, Vila J, et al. Antimicrobial resistance in Shigella spp. causing traveller's diarrhoea (1995-2010): a retrospective analysis. Travel Med Infect Dis. 2013;11(5):315-9. doi: 10.1016/j.tmaid.2013.06.010. [ Links ]
4. Antunes P, Machado J, Peixe L. Illegal use of nitrofurans in food animals: contribution to human salmonellosis?. Clin Microbiol Infect. 2006;12(11):1047-9. doi: 10.1111/j.1469-0691.2006.01539.x [ Links ]
5. García V, Montero I, Bances M, Rodicio R, Rodicio MR. Incidence and genetic bases of nitrofurantoin resistance in clinical isolates of two successful multidrug-resistant clones of Salmonella enterica serovar Typhimurium: pandemic "DT 104" and pUO-StVR2. Microb Drug Resist. 2017;23(4):405-12. doi: 10.1089/mdr.2016.0227. [ Links ]
6. McCalla DR, Kaiser C, Green MHL. Genetics of nitrofurazone resistance in Escherichia coli. J Bacteriol. 1978;133(1):10-6. [ Links ]
7. Peterson FJ, Mason RP, Hovsepian J, Holtzman JL. Oxygen-sensitive and -insensitive nitroreduction by Escherichia coli and rat hepatic microsomes. J Biol Chem. 1979;254:4009-14. [ Links ]
8. Martínez-Puchol S, Gomes C, Pons MJ, Ruiz-Roldán L, Torrents de la Peña A, Ochoa TJ, et al. Development and analysis of furazolidone-resistant Escherichia coli mutants. APMIS. 2015;123(8):676-81. doi: 10.1111/apm.12401. [ Links ]
9. Whiteway J, Koziarz P, Veall J, Sandhu N, Kumar P, Hoecher B, et al. Oxygen-insensitive nitroreductases: analysis of the roles of nfsA and nfsB in development of resistance to 5-nitrofuran derivatives in Escherichia coli. J Bacteriol. 1998;180(21):5529-39. [ Links ]
10. Shanmugam D, Esak SB, Narayanaswamy A. Molecular characterisation of nfsA gene in nitrofurantoin resistant uropathogens. J Clin Diagn Res. 2016;10(6):DC05-09. doi: 10.7860/JCDR/2016/17280.7957. [ Links ]
11. Ho PL, Ng KY, Lo WU, Law PY, Lai EL, Wang Y, et al. Plasmid-mediated OqxAB is an important mechanism for nitrofurantoin resistance in Escherichia coli. Antimicrob Agents Chemother. 2015;60(1):537-43. doi: 10.1128/AAC.02156-15. [ Links ]
12. Aviv G, Tsyba K, Steck N, Salmon-Divon M, Cornelius A, Rahav G, et al. A unique megaplasmid contributes to stress tolerance and pathogenicity of an emergent Salmonella enterica serovar Infantis strain. Environ Microbiol. 2014;16(4):977-94. doi: 10.1111/1462-2920.12351. [ Links ]
13. Ruiz-Roldán L, Martínez-Puchol S, Gomes C, Palma N, Riveros M, Ocampo K, et al. Presencia de Enterobacteriaceae y Escherichia coli multirresistente a antimicrobianos en carne adquirida en mercados tradicionales en Lima. Rev Peru Med Exp Salud Publica. 2018;35(3):425-32. doi: 10.17843/rpmesp.2018.353.3737. [ Links ]
14. Salazar de Vegas EZ, Nieves B, Araque M, Velasco E, Ruiz J, Vila J. Outbreak of infection with Acinetobacter strain RUH 1139 in an intensive care unit. Infect Control Hosp Epidemiol. 2006;27(4):397-403. doi: 10.1086/503177. [ Links ]
15. Clinical Laboratory Standard Institute (CLSI). Performance standards for antimicrobial susceptibility testing; twenty-eight informational supplement [Internet] CLSI document M100-S28. Wayne: CLSI; 2018 [citado el 11 de agosto de 2019]. Disponible en: https://clsi.org/media/2663/m100ed29_sample.pdf. [ Links ]
16. Salamanca-Pinzón SG, Camacho-Carranza R, Hernández-Ojeda SL, Frontana-Uribe BA, Espitia-Pinzón CI, Espinosa-Aguirre JJ. Correlation of the genotoxic activation and kinetic properties of Salmonella enterica serovar Typhimurium nitroreductases SnrA and cnr with the redox potentials of nitroaromatic compounds and quinones. Mutagenesis. 2010;25(3):249-55. doi: 10.1093/mutage/geq001. [ Links ]
17. Pérez-Moreno MO, Pico-Plana E, de Toro M, Grande-Armas J, Quiles-Fortuny V, Pons MJ, et al. ß-Lactamases, transferable quinolone resistance determinants, and class 1 integron-mediated antimicrobial resistance in human clinical Salmonella enterica isolates of non-Typhimurium serotypes. Int J Med Microbiol. 2013;303(1):25-31. doi: 10.1016/j.ijmm.2012.11.003. [ Links ]
18. McCracken RJ, Kennedy DG. Furazolidone in chicken: case study of an incident of widespread contamination. Br Poult Sci. 2013;54(6):704-12. doi: 10.1080/00071668.2013.850152. [ Links ]
19. Ruiz J. Transferable Mechanisms of quinolone resistance from 1998 onward. Clin Microbiol Rev. 2019;32(4):e00007-19. doi: 10.1128/CMR.00007-19. [ Links ]
20. Li XZ, Plésiat P, Nikaido H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin Microbiol Rev. 2015;28(2):337-418. doi: 10.1128/CMR.00117-14. [ Links ]
Funding sources: This work was supported by the Sociedad Española de Enfermedades Infecciosas y Microbiología Clínica (search for antibiotics and resistant microorganisms in animal feed and in animals for human consumption); JR was supported by the I3 program of the Ministry of Economy and Competitiveness, Spain (grant number: CES11/012). “ISGlobal is a member of the CERCA Programme, Generalitat de Catalunya”.
Citation: Martínez-Puchol S, Pons MJ, Ruiz-Roldán L, Laureano-Adame L, Corujo A, Ochoa TJ, et al. Resistencia a nitrofuranos mediada por mutaciones en los genes cnr y snrA en Salmonella enterica procedentes de muestras cárnicas para consumo humano. Rev Peru Med Exp Salud Publica. 2020;37(1):99-103. Doi: https://doi.org/10.17843/rpmesp.2020.371.4745
Received: August 14, 2019; Accepted: January 22, 2020
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
