










Services on Demand
Journal

Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Peruana de Medicina Experimental y Salud Publica
Print version ISSN 1726-4634
Rev. perú. med. exp. salud publica vol.37 no.2 Lima Apr-Jun 2020
http://dx.doi.org/10.17843/rpmesp.2020.372.4759
Original article
Submicroscopic placental malaria: histopathology and expression of physiological process mediators
1 Grupo Salud y Comunidad-César Uribe Piedrahíta, Universidad de Antioquia, Medellín, Colombia.
2 Facultad de Medicina, Universidad de Antioquia, Medellín, Colombia.
INTRODUCTION
Submicroscopic placental plasmodial infection (SPPI) by Plasmodium falciparum is frequent in endemic countries and contributes to the development of maternal anemia and low birth weight 1. SPPI by P. vivax (SPPI-vivax) is almost unknown in the world.
The standard technique for malaria diagnosis in endemic areas is the thick blood smear, which, by definition, does not detect submicroscopic infections, which are usually asymptomatic. These two conditions, submicroscopic and asymptomatic, had kept SPPI in absolute oblivion until the beginning of the 21st century. But there are already strong arguments suggesting that asymptomatic infections have important consequences for health and society, and should be renamed as chronic malaria infections 2.
The immunopathogenesis of placental malaria (PM) should consider pregnancy to be a unique physiological state in which the maternal immune system must protect the mother against infection and other noxae, while modulating her immune response to prevent rejection by the semi-allogeneic fetus 3. The presence of Plasmodium and its byproducts (such as hemozoin) in the placental tissue essentially alters the immune environment that regulates the placenta 4.
In the case of gestational and placental malaria, increased cytokines TNF-α, IFN-γ and IL-10 are associated with trophoblastic damage, low birth weight and prematurity 5. The expression of placental proinflammatory cytokines is high in women with PM and the expression of anti-inflammatory cytokines is low 5 - 9.
PM alters angiogenic remodeling 10 , 11. Mononuclear-cell infiltration and its inflammatory products are associated with low birth weight; and a shorter and less impactful inflammatory process in multigravid women, when comparing with primigravid women. These differences may explain the different outcomes that these women and their offspring undergo 12 - 14.
In normal pregnancy, the evident physiological processes of apoptosis, inflammation, hypoxia, vasculogenesis and angiogenesis, among others, are strictly controlled. In these processes the role of cellular communication (CC) markers/mediators is decisive 15 - 19. The CC system is a complex network between cells, that sends signals to other cells, the latter with receptor functions, producing a physiological response and a change in cellular activity. In CC, there are many markers of physiological or physiopathological processes and they have different origins (hormones, growth factors, cytokines, prostaglandins, leukotrienes, etc.) 20.
The objective of the study was to determine the association of submicroscopic infection by P. vivax or P. falciparum with histopathological events, the behavior of immune cells, cytokine-associated gene expression, hypoxia, angiogenesis and inflammation in placentas.
KEY MESSAGES
Motivation for the study: There is limited evidence on the histopathological and inflammatory effects of submicroscopic placental infection by P. falciparum and P. vivax.
Main findings: Placentas of women who live in the largest endemic area of Colombia, in the northwest of the country, were affected by submicroscopic plasmodial infection (SPPI), not detected with thick drop, but with quantitative polymerase chain reaction (qPCR). The SPPI causes tissue damage in the placenta and affects mediators of processes, such as inflammation, hypoxia, angiogenesis, among others, compared to non-infected placentas. Both P. vivax and P. falciparum act as pathogens.
Implications: Diagnostic and treatment actions for gestational plasmodial infection in prenatal consultation need to be greatly improved and should necessarily include the thick drop test in every control.
MATERIALS AND METHODS
Design and study population
A cross-sectional study was conducted by comparing three independent groups. The sample size was defined for convenience, according to the existing placental samples in the tissue bank of our research group. The selection included twenty samples with PM by vivax (PM-V), 20 with PM by falciparum (PM-F) and 19 samples without PM (PM-no) or control. The diagnostic test for plasmodial infection was the real-time quantitative polymerase chain reaction (qPCR) in placental blood.
The women were enrolled and their placentas collected in 2009 and 2016, in the hospitals of Puerto Libertador and Tierralta, municipalities in the south of Córdoba, in northwestern Colombia, a region where malaria is highly endemic 21. South Córdoba, Urabá Antioqueño and Bajo Cauca Antioqueño make up the eco-epidemiological region that generates the most cases of malaria annually in Colombia 21.
The inclusion criteria were the following: permanent residence in this region for at least the last year; no history of pre-eclampsia/eclampsia, hypertensive pregnancy disease, diabetes, HIV, and toxoplasmosis, rubella, cytomegalovirus, herpes simplex (TORSCH); and giving birth in one of the hospitals having a 36-to-41-week gestation.
Gestational age was taken from the medical record. Women in the control group had to be afebrile and those in the plasmodial infection group may or may not have had a fever (only 1% had a fever); they should also voluntarily agree to participate in the study. The exclusion criteria were withdrawal of consent or the occurrence of any complication or disease.
Malaria diagnosis
The maternal peripheral blood was obtained at the time of delivery. The placenta was cleansed with saline solution (0.9%) and blood and tissue samples were taken from the maternal side of the placenta for diagnosis (thick drop, qPCR, histopathology) as indicated in other reports 22 - 25. The researchers read the thick drop samples and considered them negative when 200 fields, with 100x magnification, were free of parasites.
The DNA was extracted using the Chelex-saponin method and qPCR 22 , 23. The qPCR was run on the ABI 7500 FAST platform. Samples with a cycle threshold (Ct) <45 were analyzed in duplex species-specific reactions for P. falciparum and P. vivax 23. Amplification of the 18S rRNA genes of the DNA was used for quantification. The DNA copy number was quantified from the genus-specific reaction against a standard curve using a plasmid containing a fragment of the 18S gene for P. falciparum.
Histopathological study
Placental tissue was processed according to standardized procedures 24 , 25. Two fragments were taken from each placenta and from each one a plate was made for histological study by light microscopy with classic procedures. One fragment came from a point near the umbilical cord insertion and the other from the middle zone (equidistant between the cord and the placental border). A total of 40 fields were read (20 per fragment). The reading was compared against the results of thick drop and qPCR. Total magnification of 400X was used for general histological reading. Total magnification of 1000X was used to determine the presence of infected erythrocytes (iE) or hemozoin.
Quantification of the expression of genes associated with markers/mediators
A tissue fragment preserved with RNA Later® (Qiagen) at 4 °C was used to quantify the expression of genes associated with markers/mediators and cytokines in placental tissue. The mediators were grouped as follows: proinflammatory (IL-2, TNF, IFNɣ, COX-1, COX-2, C5a), anti-inflammatory (IL-10, IL-4), angiogenic (VEGF, VEGFR-1) and hypoxic (HIF).
Different markers, except C5a, were measured by relative quantification of mRNA by qPCR. Relative quantification was done to determine the expression levels of the study mediators in relation to the expression levels of the constituent gene and to obtain the relationship between the gene of interest and the constituent gene. The Pfaffl procedure was applied to determine delta-delta TC as follows 26:

By following the manufacturer’s recommendations, the commercial kit Human C5a Elisa Kit was used to quantify the complement C5a fraction in placental serum samples.
Secondary information sources and study groups
After inclusion, a clinical-epidemiological questionnaire was applied. The medical record was used as a data source. Parturient patients and their placentas were evaluated, distributed in three groups: 19 in the PM-no group or control group, 20 in the PM-F group and 20 in the PM-V group.
Statistical analysis
SPSS 18.0 and GraphPad Prism 5 were used. Significance decisions were made with a probability of less than 0.05. The Kolmogorov-Smirnov test was used to assess the normal distribution of quantitative variables. Levene’s test was used to assess the homoscedasticity of variances. The Mann-Whitney test was used to determine if there was a difference between two independent groups. The non-parametric Kruskal-Wallis test was applied to determine if there was a difference between three independent groups, then the Dunn test was used to identify the pairs of groups that differed.
RESULTS
From the total, 95% (38/40) of PM cases are submicroscopic infections. From the pregnant women total, 25% (15/59) had history of malaria during the current pregnancy.
The age range was from 14 to 41 years and the average in the three groups was similar (PM-no: 22; PM-F: 25; PM-V: 23). The average gestational age was 38.6 weeks (range from 36 to 41). Previous pregnancies averaged 2.8. From the total, 36% were in their first gestation, 20% in their second and 44% were multi-gestational (3 to 9). Of the 59 deliveries, 5 were by caesarean section.
At the time of delivery, the average hemoglobin value was 11.1 g/dL. Women with PM had lower hemoglobin levels than uninfected women (10.86 g/dL and 11.69 g/dL, respectively). Hemoglobin was lower in the PM-V group (10.75 g/dL) than in the PM-F group (10.97 g/dL), but without significant difference (p=0.506). The mean neonatal weight in the PM-no group was 2,974 g; in the PM-F group, 2,852 g; and in the PM-V group, 2,737 g. The difference in mean neonatal weight was 237 g between children in the PM-V and PM-no groups, and 115 g between children in the PM-V and PM-F groups.
The control group (PM-no) had absence of necrosis and higher frequency of atherosis, abruptio and thrombus than the other two. The PM-V group presented similar results to PM-F in atherosis, necrosis, infarction, fibrin deposits and thrombi, but less hemozoin and infected erythrocytes. Fibrin deposits were observed in all placentas. In the intervillous space, 95-100% of placentas showed hemorrhage and 6 out of 10 placentas showed thrombi and calcifications. There was significant difference (p < 0.05) between the three groups regarding abruption, syncytial nodes and hemozoin. The PM-F group generated this difference. In the species comparison, there was no significant difference (p > 0.05) (Table 1).
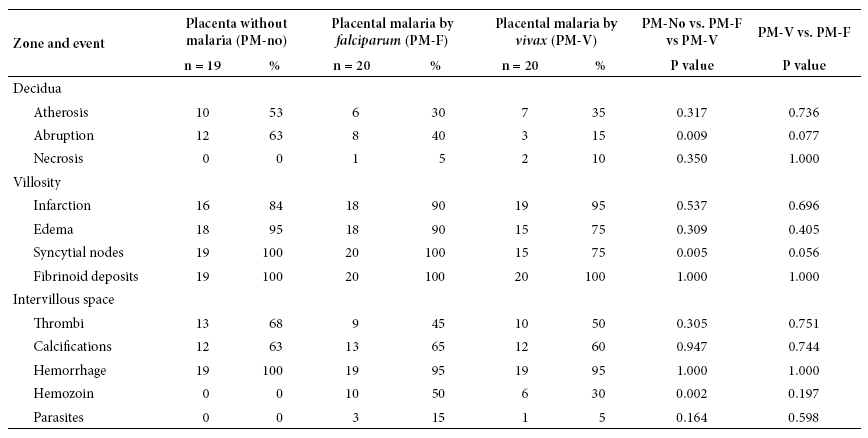
In the PM-no group, atherosis and abruption are rare, averaging 1.3 and 2.2 events, respectively. In the same order; infarction and villous edema had 9.1 and 8.7 events on average. In the same PM-no group, the amount of fibrin and syncytial node deposits is 72.5 and 109 on average. Placentas without infection have 366 villi, 1463 capillaries and 3.97 capillaries per villi. The average number of hemorrhages was 18, while the average number of thrombi and calcifications were 2 (they were in 60-70% of them) (Table 2).
Table 2 Magnitude of placental histological events, according to the presence of plasmodial infection
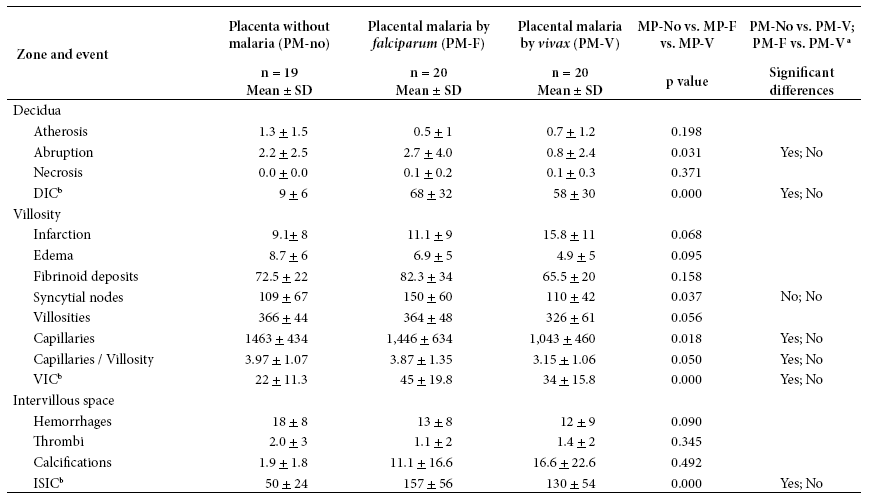
a Indicates whether the Dunn test showed a difference between each pair of comparisons (PM-no vs. PM-V; PM-V vs. PM-F) with p < 0.05.
b DIC: Deciduous immune cells. VIC: villous immune cells. ISIC: immune cells in intervillous space. SD: standard deviation
In PM-F and PM-V groups, the appearance of placental infection reduces the amount of atherosis, villous edema, capillary hemorrhage and thrombi, but increases villous infarction, calcifications and immune cells. Only a few events show significant difference (Table 2).
Immune cells were found in all placentas, in all three compartments, and regardless of infection. The quantities were different, in the PM-no group, 9 immune cells in decidua, 22 in villous and 50 in intervillous space were observed; in the PM-V group, 58, 34 and 130 were observed respectively; in the PM-F group the highest amount of immune cells were found: 68, 45 and 157, respectively (Table 2).
Table 3 shows the expression of inflammatory, angiogenic and hypoxic mediators by group. Inflammation mediators (COX-1, COX-2, IL-10, IFNɣ, TNF, C5a) were significantly higher in PM. IL-2 and IL-4 varied little among the three groups. Although C5a did not show statistically significant difference between the groups, it had higher values in PM-V compared to the two other groups. Regarding angiogenesis, VEGF and VEGFR-1 showed no significant difference between groups, but their expression increased in groups with PM. Regarding hypoxia, HIF-1α was significantly different and with higher values in groups with PM. In general, the mediators that showed no significant difference between the three groups were IL-2, VEGF, VEGFR-1 and C5a. For VEGFR-1 and C5a, the high variability within each group is probably a factor affecting statistical significance when comparing the three groups.
Table 3 Expression of inflammation, angiogenesis and hypoxia mediators according to study group
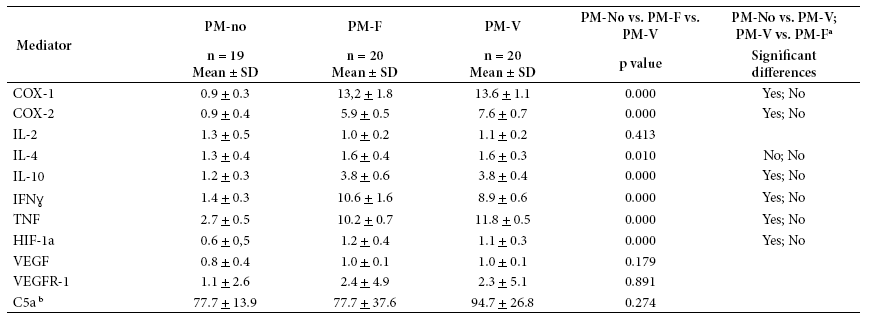
a Indicates whether Dunn’s test showed a difference between each pair of comparisons (PM-no vs. PM-V; PM-V vs. PM-F), with p < 0.05.
b For C5a, concentration in blood was measured and not the expression of the associated gene.
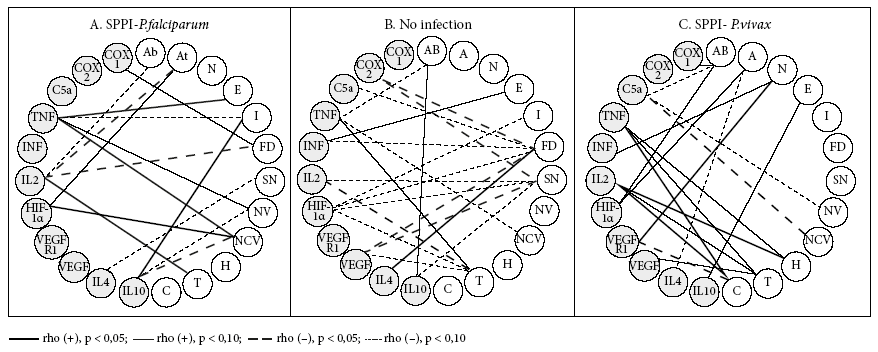
Figures 1, 2 and 3 show the bivariate linear correlations between histological events and process mediators. In the group without infection, the predominance of negative and weak (p < 0.10) significant correlations (SC) is clear. Fibrin deposits, syncytial nodes and thrombi have the highest amount of SC. The 6 SC that fibrinoid deposits have are made with 7 process mediators, which are inflammation promoters (COX-2 [but not COX-1], C5a, IFNɣ [but not TNF], VEGF) and hypoxia (HIF) and angiogenesis mediators (VEGF, but not its receptor). The 5 SC of the syncytial nodes are made with 3 of the mediators that are also present in the relations with the fibrinoid deposits (COX-2, HIF and VEGF) and with cytokines 2 (pro-inflammatory) and 10 (anti-inflammatory). The 4 SC of thrombi occur with the inflammation mediators, TNF, IL-2, VEGF and the hypoxia mediator HIF.
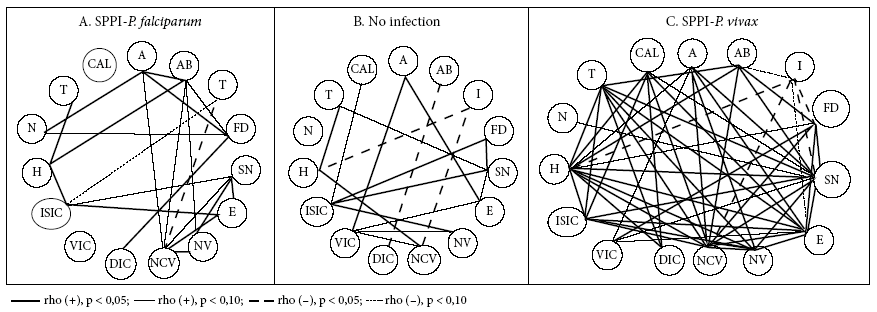
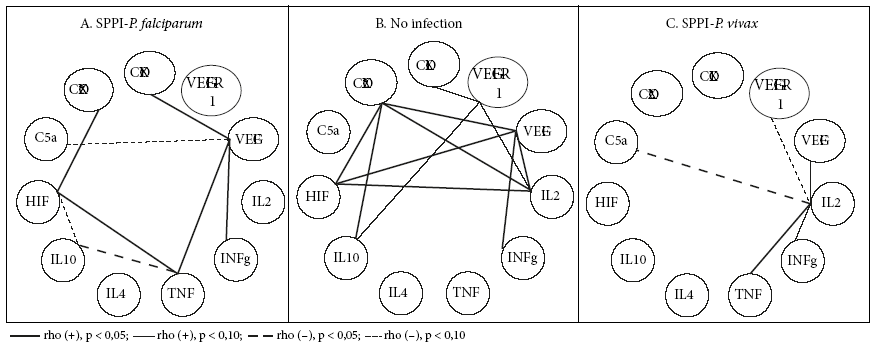
Figure 2 Significant bivariate linear correlations (CS) between mediators of placental processes of angiogenesis, inflammation and hypoxia.
On the other hand, it is necessary to highlight the SC that several mediators establish with some histological events. This is the case with HIF, which, in addition to being associated with syncytial nodes, fibrinoid deposits and thrombi, it is also associated with infarction. TNF is associated with abruptio, thrombi and villous capillaries.
In the PM-F group, SC reduce and the polarity changes; in this case, the poles are not the syncytial nodes, fibrinoid deposits and thrombi, but the only pole that stands out with the villous capillaries (three SC: with TNF, HIF and IL-10). TNF had SC with edema, infarction, the number of villi and the number of capillaries per villi; IL-2 had SC with abruption, atherosis, thrombi and fibrinoid deposits.
For the PM-V group, SC abound as in the absence of infection, but with different focuses: abruption, thrombus, calcifications. The SC of abruption are formed with COX-1, C5a and HIF-1α; those of thrombi and calcifications with TNF and IL-2. Thrombi are also associated with VEGF, whereas calcifications are related to VEGFR-1.
DISCUSSION
This study found important differences in the placental effects of the two plasmodial species evaluated. The amount of abruption, syncytial nodes, villi, capillaries and capillaries by villi is significantly lower when P. vivax is present, compared to P. falciparum. On the other hand, placentas infected with P. vivax, when compared to placentas without infection, generated more infarction and less abruption, edema, villi, capillaries and capillaries by villi. There is evidence of the pathogenic nature of PM-SPPI due to P. falciparum in pregnant women and their children 1. Even in studies in the same geographical area where this study took place 27 - 29, submicroscopic placental infection by P. vivax causes harmful effects.
Comparisons of the P. vivax group against the P. falciparum group or against the group without infection strongly support the placental pathogenic capacity of P. vivax. These findings provide a basis for explaining its effects by a mechanism that does not necessarily involve the cytoadhesion of infected erythrocytes to the placental tissue, as occurs in P. falciparum infections.
There is scarce information on the correlations between histological events and process mediators. When there is no infection, SC between histological events and process mediators are predominantly less than 10%. This can be interpreted as a homeostatic state, in which no specific response predominates, but all of them are active, in basal and balanced states. Otherwise gestation would be at risk.
When there is a plasmodial infection, the panorama for SC changes radically, both in the case of placental plasmodial infection by P. falciparum (SPPI-F), but more so in the case of placental plasmodial infection by P. vivax (SPPI-V). SC in the non-infection state disappear and new ones arise in plasmodial infections. The centers or poles of SC that were the syncytial nodes and fibrinoid deposits disappear and are replaced by poles that now correspond to the process mediators, mainly inflammation. IL-2, TNF and IL-10 are poles in SPPI-F, while IL-2, TNF and C5a are poles in SPPI-V. In the absence of infection, the villi and capillaries are virtually CS-free; when infection occurs, they emerge as poles. All this would express the exacerbated inflammatory state of the placenta as a consequence of the infection.
In this study, no hormones were measured at any point in gestation. Progesterone, estrogens, androgens and glucocorticoids are involved in gestation from implantation to delivery; their biosynthesis and metabolism are the result of complex pathways involving the fetus, the placenta and the mother 30. Many sexual hormones clearly interact with the immune system and many mediators of physiological processes also interact with the immune system. There is, therefore, a complex physiological network between hormones, mediators of physiological processes and the immune system.
One of the strengths of this study is the comprehensive approach to PM, addressing the relationship between histopathological events and mediators of inflammation, angiogenesis and hypoxia. To our knowledge these relationships have not been studied before. We had a well-defined control group, which was negative for Plasmodium according to highly sensitive and specific technique (qPCR), without the presence of TORCH syndrome, HIV, eclampsia/preeclampsia, and diabetes. However, the presence of intestinal parasites and malnutrition could not be ruled out.
We consider the small sample size and the selection by convenience as limitations due to time and money available. The mediators evaluated basically represent the inflammatory process of the placenta at full-term, other process mediators were not sufficiently measured. The analysis of the correlations between histological findings and process mediators is superficial because there is little or no information available.
In conclusion, when SPPI occurs, there are placental tissue changes as well as changes in the expression of inflammatory process mediators, whether the causal agent is P. falciparum or P. vivax. If the parasite is pathogenic to the placenta, it is to be expected that when parasitemia increases, effects will be stronger. Placental infection with P. vivax contributes to the increase in placental histological findings associated with tissue damage and deterioration. The alteration of the placental structure was mainly associated with the decrease of villi and the number of capillaries per villi, as well as with the increase of ischemic degenerative lesions associated with calcifications and infarction. These results could be correlated with low birth weight and low hemoglobin levels.
REFERENCES
1. Arango E, Maestre A, Carmona-Fonseca J. Efecto de la infección submicroscópica o policlonal de Plasmdium falciparum sobre la madre y el producto de la gestación. Revisión sistemática. Rev Bras Epidemiol. 2010;13(3):373-86. doi: 10.1590/s1415-790x2010000300002. [ Links ]
2. Chen I, Clarke SE, Gosling R, Hamainza B, Killeen G, Magill A, et al. "Asymptomatic" malaria: a chronic and debilitating infection that should be treated. PLoS Med. 13(1): e1001942. [ Links ]
3. Mor G, Cardenas I. The immune system in pregnancy: a unique complexity. Am J Reprod Immunol. 2010;63(6):425-33. [ Links ]
4. Rogerson SJ, Hviid L, Duffy PE, Leke RF, Taylor DW. Malaria in pregnancy: pathogenesis and immunity. Lancet Infect Dis. 2007;7(2):105-17. [ Links ]
5. Menendez C, Ordi J, Ismail MR, Ventura PJ, Aponte JJ, Kahigwa E, et al. The impact of placental malaria on gestational age and birth weight. J Infect Dis. 2000;181(5):1740-5. [ Links ]
6. Chaisavaneeyakorn S, Lucchi N, Abramowsky C, Othoro C, Chaiyaroj SC, Shi YP, et al. Immunohistological characterization of macrophage migration inhibitory factor expression in Plasmodium falciparum-infected placentas. Infect Immun. 2005;73(6):3287-93. [ Links ]
7. Suguitan AL, Leke RG, Fouda G, Zhou A, Thuita L, Metenou S, et al. Changes in the levels of chemokines and cytokines in the placentas of women with Plasmodium falciparum malaria. J Infect Dis. 2003;188(7):1074-82. [ Links ]
8. Lucchi NW, Koopman R, Peterson DS, Moore JM. Plasmodium falciparum-infected red blood cells selected for binding to cultured syncytiotrophoblast bind to chondroitin sulfate A and induce tyrosine phosphorylation in the syncytiotrophoblast. Placenta. 2006;27(4-5):384-94. [ Links ]
9. Salih MM, Mohammed AH, Mohmmed AA, Adam GK, Elbashir MI, Adam I. Monocytes and macrophages and placental malaria infections in an area of unstable malaria transmission in eastern Sudan. Diagn Pathol. 2011;6:83. [ Links ]
10. Hromatka BS, Ngeleza S, Adibi JJ, Niles RK, Tshefu AK, Fisher SJ. Histopathologies, immunolocalization, and a glycan binding screen provide insights into Plasmodium falciparum interactions with the human placenta. Biol Reprod. 2013;88(6):154. doi: 10.1095/biolreprod.112.106195. [ Links ]
11. Souza RM, Ataíde R, Dombrowski JG, Ippólito V, Aitken EH, Valle SN, et al. Placental histopathological changes associated with Plasmodium vivax infection during pregnancy. PLoS Negl Trop Dis. 2013;7(2):e2071. doi: 10.1371/journal.pntd.0002071. [ Links ]
12. Duffy PE. Plasmodium in the placenta: parasites, parity, protection, prevention and possibly preeclampsia. Parasitology. 2007;134(Pt 13):1877-81. [ Links ]
13. Hartman TK, Rogerson SJ, Fischer PR. The impact of maternal malaria on newborns. Ann Trop Paediatr. 2010;30(4):271-82. [ Links ]
14. Kabyemela ER, Fried M, Kurtis JD, Mutabingwa TK, Duffy PE. Fetal responses during placental malaria modify the risk of low birth weight. Infect Immun. 2008;76(4):1527-34. [ Links ]
15. Lucchi NW, Peterson DS, Moore JM. Immunologic activation of human syncytiotrophoblast by Plasmodium falciparum. Malar J. 2008;7:42. doi: 10.1186/1475-2875-7-42. [ Links ]
16. Valdivia-Silva JE, González-Altamirano C, López-Molina K, Lazo-Velásquez JC, García-Zepeda E. Relación de la expresión del factor inducido por hipoxia-2a (HIF-2a) y sVEGF-R1/sFlt-1: implicación en la fisiopatología de preeclampsia. Clin Invest Ginecol Obstet. 2008;35(1):11-23. [ Links ]
17. Soeters PB, Grimble RF. The conditional role of inflammation in pregnancy and cancer. Clin Nutr. 2013;32(3):460-5. [ Links ]
18. Laakkonen JP, Lähteenvuo J, Jauhiainen S, Heikura T, Ylä-Herttuala S. Beyond endothelial cells: Vascular endothelial growth factors in heart, vascular anomalies and placenta. Vascul Pharmacol. 2019;112:91-101. doi: 10.1016/j.vph.2018.10.005. [ Links ]
19. Melincovici CS, Bosca AB, Susman S, Marginean M, Mihu C, Istrate M, et al. Vascular endothelial growth factor (VEGF) - key factor in normal and pathological angiogenesis. Rom J Morphol Embryol. 2018;59(2):455-467. [ Links ]
20. Biocancer. Tipos y moléculas de señalización. Acceso: 7 junio 2019. Disponible en: http://www.biocancer.com/journal/1101/11-tipos-y-moleculas-de-senalizacion . [ Links ]
21. Carmona-Fonseca J. Región Urabá Antioqueño-Cuencas altas de ríos Sinú y San Jorge-Bajo Cauca Antioqueño : guarida del paludismo colombiano. Rev Salud UIS. 2017;49(4):577-589. [ Links ]
22. Plowe CV, Djimde A, Bouare M, Doumbo O, Wellems TE. Pyrimethamine and proguanil resistance-conferring mutations in Plasmodium falciparum dihydrofolate reductase: polymerase chain reaction methods for surveillance in Africa. Am J Trop Med Hyg. 1995; 52(6):565-8. doi: 10.4269/ajtmh.1995.52.565. [ Links ]
23. Shokoples SE, Ndao M, Kowalewska-Grochowska K, Yanow SK. Multiplexed real-time PCR assay for discrimination of Plasmodium species with improved sensitivity for mixed infections. J Clin Microbiol. 2009; 47(4):975-80. doi: 10.1128/JCM.01858-08. [ Links ]
24. Carmona-Fonseca J, Arango E, Maestre A. Placental malaria in Colombia: histopathologic findings in Plasmodium vivax and P. falciparum infections. Am J Trop Med Hyg. 2013; 88(6):1093-101. doi: 10.4269/ajtmh.12-0363. [ Links ]
25. Agudelo-García OM, Arango-Flórez EM, Carmona-Fonseca J. Submicroscopic and asymptomatic congenital infection by Plasmodium vivax or P. falciparum in Colombia: 37 cases with placental histopathology and cytokine profile in maternal and placental blood. J Trop Med. 2017; 2017:368078. [ Links ]
26. Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):e45. doi: 10.1093/nar/29.9.e45. [ Links ]
27. Carmona-Fonseca J, Agudelo O, Arango E. Asymptomatic plasmodial infection in Colombian pregnant women. Acta Trop. 2017; 172:97-101. doi: 10.1016/j.actatropica.2017.04.030. [ Links ]
28. Agudelo O, Aristizabal B, Yanow S, Arango E, Carmona-Fonseca J, Maestre A. Submicroscopic infection of placenta by Plasmodium produces Th1/Th2 cytokine imbalance, inflammation and hypoxia in women from Northwest Colombia. Malar J. 2014;13:122. [ Links ]
29. Agudelo-García OM. Malaria placentaria submicroscópica por Plasmodium vivax o Plasmodium falciparum: histopatología, células inmunitarias y expresión de genes asociados a hipoxia, apoptosis e inflamación. Trabajo de investigación para optar el título de Doctor en Ciencias Básicas Biomédicas. Corporación Académica Ciencias Básicas Biomédicas, Grupo Salud y Comunidad-César Uribe Piedrahíta, Universidad de Antioquia; Medellín, octubre 2017. [ Links ]
30. Morel Y, Roucher F, Plotton I, Goursaud C, Tardy V, Mallet D. Evolution of steroids during pregnancy: Maternal, placental and fetal synthesis. Ann Endocrinol (Paris). 2016;77(2):82-9. doi: 10.1016/j.ando.2016.04.023. [ Links ]
Funding sources: Colciencias Project No. 111577757051, Contract No. 755-2017; Sustainability Strategy Codi-Universidad de Antioquia 2016-2017 (code ES-84160127); Universidad de Antioquia.
Citation: López-Guzmán C, Carmona-Fonseca J. Submicroscopic placental malaria: histopathology and expression of physiological process mediators. Rev Peru Med Exp Salud Publica. 2020;37(2):220-8. doi: https://doi.org/10.17843/rpmesp.2020.372.4759.
Received: August 21, 2019; Accepted: February 26, 2020
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
