











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
El cáncer colorrectal constituye la tercera causa más frecuente de cáncer en el mundo, su incidencia y letalidad han disminuido en pacientes mayores de 50 años, pero ha aumentado en pacientes más jóvenes 1. En EE.UU., el diagnóstico y tratamientos tempranos han logrado que las tasas de supervivencia a 5 años estén alrededor del 91 % para el estadio I y 82 % para el estadio II; sin embargo, solo el 20 % y 22 % de pacientes, respectivamente, son diagnosticados en esos estadios; la supervivencia declina a 68 % en estadio III y a 12 % en estadio IV 2. En el Perú, de acuerdo con el último registro de cáncer de Lima Metropolitana (periodo 2013-2015), la tasa de incidencia de cáncer colorrectal estandarizada es de 10,94 casos por 100,000 habitantes, lo que representó el 5 % del total de las neoplasias malignas 3. A pesar de no contar con estadísticas por estadio para Perú, los autores sospechamos, basándonos en nuestras experiencias profesionales, que los estadios avanzados sobrepasan largamente a los tempranos.
El tracto gastrointestinal constituye el sistema fundamental clave para la digestión de los alimentos y la absorción de nutrientes. En estos procesos existe un continuo recambio celular a través de la generación y desarrollo de las células madre intestinales, las cuales se diferenciarán en varios tipos celulares, incluidos enterocitos, células de Paneth, células caliciformes, células enteroendocrinas, entre otras 4. Las vías de señalización Wnt son un factor fundamental en la función intestinal normal, sobre todo, con relación a la replicación y mantenimiento de las células madre epiteliales, ubicadas en la base de las criptas intestinales. Las alteraciones en diferentes niveles de estas vías promueven la hiperactividad de señales y son críticas para la oncogénesis colónica, por lo que la vía de señales Wnt es considerada un conductor del cáncer colorrectal en prácticamente todos aquellos que poseen alteraciones en genes claves para su regulación 5. No se tiene aún claro cómo se produce una señal específica dentro de todas las posibles variantes en la vía Wnt, pero probablemente dependa del tipo celular y de qué clase de componentes de señalización se encuentra expresada en ese momento en el contexto celular 6.
Las alteraciones en la vía Wnt/β-catenina son muy importantes en la oncogénesis del cáncer colorrectal y, por tanto, es indispensable continuar investigando al respecto, pues, a pesar de la gran cantidad de publicaciones que se tiene sobre sus alteraciones en diversos tipos de cáncer, esta es insuficiente para todas las preguntas que se tienen aún por contestar.
El objetivo de esta revisión es conocer los nuevos mecanismos oncogénicos relacionados al cáncer colorrectal por alteraciones genéticas ligadas a la vía de señalización canónica (a la cual llamaremos Wnt/β-catenina); para ello, incluimos información sobre su historia, revisión de la vía de señalización dependiente de β-catenina, su interacción con los otros componentes, sus alteraciones a nivel genético y posibles blancos terapéuticos en base a la más reciente información disponible.
Estrategia de búsqueda
Se consultaron las bases de datos PubMed, Scopus y SciELO, a partir del año 2000 hasta el 2022. Se buscaron todas las referencias con relación a la vía canónica Wnt (WTN/β-catenina) asociadas al cáncer colorrectal, con las palabras clave: Vía de señalización Wnt, beta Catenina, neoplasias colorrectales, genes APC (español); Wnt proteins, Wnt signaling pathway; colorectal neoplasms; genes, APC (inglés). De los artículos seleccionados, se excluyeron aquellas revisiones repetitivas, y se consideraron las fuentes originales como prioritarias.
Cáncer colorrectal
El cáncer colorrectal (CCR) es una de las neoplasias malignas humanas más frecuentes en el mundo, la cual generalmente se asociaba a países con mejores condiciones de vida, pero cuyos casos van aumentando paulatinamente en países en vías de desarrollo 7. Los avances en el conocimiento molecular, en las estrategias de prevención y detección temprana, así como en el tratamiento, han conseguido resultados muy alentadores (tasa de supervivencia a 5 años del CCR en estadio localizado es del 91 %; si el cáncer se diseminó a los tejidos u órganos circundantes y/o a los ganglios linfáticos regionales, la tasa de supervivencia a 5 años es del 72 %) 8; sin embargo, sigue siendo un importante problema de salud pública en pacientes en estadio IV, en los cuales la supervivencia a 5 años no llegaba ni al 13 % en Perú en el 2019 9.
El CCR fue uno de los primeros tumores en los que se construyó un modelo de oncogénesis, propuesto por Vogelstein et al 10. Esto se hizo en forma secuencial y se explicó cómo se produce la formación de los adenomas y cómo evolucionaban hacia el fenotipo carcinomatoso a través de la acumulación de distintos eventos genéticos y epigenéticos. Este modelo describió las distintas alteraciones conductoras de los principales oncogenes (K-ras, N-ras, B-raf y PI3K) y los genes supresores de tumores (APC, TP53 y PTEN) implicados en la biología del CCR 11 (Figura 1).
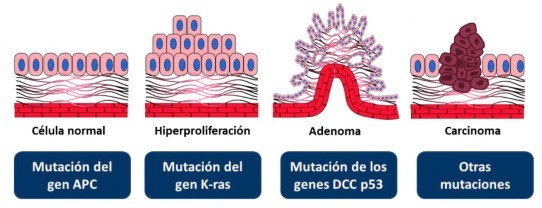
Figura 1 Modelo genético que explica el proceso de carcinogénesis al cáncer colorrectal desde el epitelio normal hasta el carcinoma esporádico. Las alteraciones se originan por diversas mutaciones adquiridas en células somáticas, que ocasionan la inactivación de genes supresores de tumores y la amplificación de oncogenes. Modificado de: Multistep model of carcinogenesis. The stepwise genetic alterations that lead to colorectal cancer. From Aspinall and Taylor-Robinson, 2001.
Las alteraciones moleculares patológicas del cáncer colorrectal han sido ampliamente estudiadas; no obstante, todavía se continúan estudiando para comprender mejor sus mecanismos e interacciones. El cáncer colorrectal puede clasificarse en función a sus características moleculares en dos tipos principales: A) tumores con inestabilidad cromosómica (CIN) y estabilidad microsatélite (MSS), no hipermutados, los cuales comprenden el 85 % de todos los adenocarcinomas del CCR y están asociados a la pérdida del gen supresor de tumores APC, como evento temprano en la formación de adenomas; B) tumores con inestabilidad de microsatélites o hipermutados (MSI), generalmente encontrados en el colon derecho y asociados con el fenotipo metilador de las islas CpG (CIMP) 12.
En los últimos años, los avances de la biología molecular están considerando una tercera vía para explicar la carcinogénesis del CCR, la cual no está asociada a cambios a nivel cromosómico ni a tumores con inestabilidad en microsatélites. Esta vía se produce por hipermetilación de los genes promotores en regiones específicas del ADN ricas en islas CpG. Esta vía se denomina vía aserrada, por la apariencia del tumor en la evaluación histológica 13.
Las vías de señalización WNT
Son un conjunto de vías de transducción de señales, las cuales participan en varios mecanismos del desarrollo celular, como proliferación, diferenciación, apoptosis, migración, mantenimiento de células madre, entre otros. La familia de genes Wnt es muy grande y su estructura genética está altamente conservada en el mundo animal, por lo que juega un papel fundamental en el desarrollo embrionario. Esta vía puede agruparse en dos tipos: a) vía canónica (o β-catenina dependiente), relacionada con la regulación de transcripción de genes; b) vía no canónica (o β-catenina independiente), que incluye la vía de la polaridad celular planar, que regula el citoesqueleto en relación con la forma de la célula y la vía no canónica Wnt/calcio, que regula este ion dentro de la célula, entre otras vías no canónicas actualmente en investigación. Esta división, sin embargo, es convencional, ya que estas vías forman una red de comunicación con diversas interacciones entre sus miembros con regulación mutua 14,15.
Existen 19 genes de la familia Wnt que son potestad de los animales multicelulares. Las proteínas Wnt pueden enviar señales a través de una amplia gama de receptores, pero aquellos más ampliamente reconocidos son los receptores Frizzled (Fzd), los cuales comprenden una familia de 10 receptores acoplados a proteínas G diferentes. Las proteínas Wnt pueden provocar una variedad de respuestas en la célula objetivo a través de la activación del receptor Fzd, pero la mejor estudiada es la vía de señalización canónica 16.
Las alteraciones que ocurren en la vía Wnt causan distintas enfermedades, tales como osteoporosis, artritis reumatoide y cáncer, por lo cual es un objetivo terapéutico importante en la búsqueda de nuevas moléculas selectivas e inteligentes 17.
Descubrimiento de la vía WNT
Sabemos que los estudios genéticos están inexorablemente ligados al estudio de las criaturas menos complejas en su estructura cromosómica; uno de estos es, sin duda, la mosca de las frutas. Las mutaciones que se lograron en estos insectos proporcionaron muchos conocimientos, como, por ejemplo, el hallazgo de un gen necesario para el desarrollo de las alas en la Drosophila melanogaster, el cual fue llamado gen wg (derivado del Wingless [sin alas]) y fue descubierto por Sharma 18. Años más tarde, en experimentos que integraban el virus del tumor mamario del ratón (MMTV), con protooncogenes activando la formación de tumores, se reconoció al gen int-1 19. Posteriormente se descubrió que había homología entre los genes wg e int-1 y se propuso llamarlos genes Wnt, que es un acrónimo creado a partir de los nombres Wingless e Int-1 20.
En 1991, las mutaciones del gen de la poliposis coli adenomatosa (APC) fueron descubiertas y asociadas como la causa subyacente del cáncer de colon hereditario al síndrome denominado poliposis adenomatosa familiar (FAP) 21,22. Dos años después se demostró que el gen APC interactuaba con la β-catenina, dentro de la vía canónica Wnt, para producir las alteraciones subyacentes al CCR 23,24. También se demostró que la pérdida de la función de APC producía un aumento de la actividad del factor de células T (TCF), un factor de transcripción nuclear cuando se asociaba a la β-catenina 25.
Posteriores estudios han ido completando los componentes y procesos identificados en la vía de señalización WNT, con el auxilio de la tecnología de secuenciación génica. Asimismo, se demostró que las mutaciones de esta vía están frecuentemente alteradas en algunos cánceres humanos 26.
Producción y secreción del ligando WNT
La comunicación celular es fundamental para el desarrollo de los organismos multicelulares y para su homeostasis. Una forma común de lograr esta comunicación se produce a través de la secreción de moléculas de señalización que son recibidas por los receptores de las células vecinas o a distancia, las cuales producirán diferentes efectos a nivel nuclear. Los ligandos Wnt son una gran familia de glicoproteínas secretadas, hidrófobas, que controlan una variedad de procesos. Sin embargo, para que el ligando Wnt sea activo, necesita ser procesado en el retículo endoplásmico y secretado a través del aparato de Golgi hacia la membrana celular 27.
Después de su síntesis, la proteína Wnt es modificada por lipidación en un residuo de cisteína, lo cual es fundamental para su función. La aciltransferasa puercoespín (PORCN) es una enzima residente en el retículo endoplásmico, necesaria para la unión del ácido palmitoleico al ligando Wnt 28; la inhibición de la aciltransferasa por la 2-bromopalmitato o por la remoción enzimática del palmitato o por otros medios da como resultado que el ligando no sea activo 29. La palmitoilación es necesaria en la vía de señalización de Wnt porque inicia el direccionamiento de la proteína Wnt a la membrana plasmática para su secreción y permite que se una a su receptor, debido a la unión covalente de los ácidos grasos 30. Una vez que Wnt está asociado a lípidos, es exportado al aparato de Golgi, donde se une a las proteínas transmembrana Wntless/Evenness interrupted (Wls/Evi/Mom-3) para ser secretados hacia la membrana plasmática 31,32. Después de la secreción del ligando Wnt, las proteínas Wls/Evi/Mom-3 se someten a endocitosis a base de clatrina y se reciclan al aparato de Golgi por el complejo retrómero (proteínas muy importantes en el reciclado de receptores transmembrana) 33.
Los ligandos WNT pueden estar en la membrana celular o salir de la célula por múltiples rutas, incluidas la liberación directa de la membrana plasmática por solubilización, la formación de exosomas o en partículas de proteínas lipídicas. Los diferentes mecanismos de liberación de los ligandos WNT pueden estar asociados a la función que tiene que cumplir.
Vía canónica del WNT (WNT/β-catenina)
Antes de describir los fenómenos y procesos que suceden en la vía canónica, es importante saber que algunos componentes son compartidos por ambas vías, como Fzd y Dvl (proteína Disheville). Asimismo, algunos efectos de la vía canónica pueden ser antagonizados por la vía no canónica y ambas vías están estrechamente relacionadas, de tal forma que no es posible concebirlas de forma paralela, sino, más bien, de forma complementaria. En lo concerniente a esta revisión, cabe destacar que la vía no canónica puede tener funciones tanto promotoras como represivas del CCR 34-37.
Vía Wnt/β-catenina no activa
La vía de señalización de Wnt actúa controlando la expresión génica y otras funciones celulares de forma autocrina o paracrina. Los ligandos Wnt, los cuales son glicoproteínas modificadas con lípidos de β40 kDa, actúan como morfógenos al unirse al receptor Frizzled y al correceptor RLP5 o RLP6 (proteína relacionada al receptor de LDL), formando un trímero, que inicia la cascada de acciones de la vía.
No se activa la vía Wnt/β-catenina en los siguientes casos: a) ausencia de los ligandos Wnt; b) presencia del ligando, pero no llegan a actuar con el receptor por presencia de algunos factores como WIF (factor inhibidor de Wnt); SFRP (proteína secretada relacionada a Frizzled), que impide que el ligando se acople al receptor FZD; DKK (proteína relacionada a Dickkopf), que inhibe a RLP5 o 6, lo que imposibilita la formación del trímero; c) inducción de la expresión de ligasas E3 que promueven la ubiquitinación de FZD y la internalización de este receptor para ser degradado por los proteasomas (Figura 2).
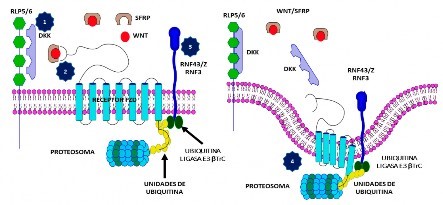
Figura 2 Mecanismos que impiden la formación del trímero activador de la vía Wnt/β-catenina (WNT-FZD-RLP5/6): 1) bloqueo del correceptor RLP5/6 por DKK; 2) bloqueo del ingreso del ligando al receptor FZD, por la presencia de SFRP; 3) activación de RNF43/ZRNF3 que acciona la ubiquitina ligasa E3 βTrC, lo que promueve la ubiquitinación del receptor FZD; 4) FZD, posteriormente, se internaliza y luego es destruida por los proteosomas. RLP5/6: proteínas relacionadas al receptor 5 o 6 del LDL; DKK: proteína relacionada a Dickkopf; SFRP: proteína secretada relacionada a Frizzled; RNF43: ring finger protein 43; ZNRF3: zinc and ring finger protein 3; β-TrC: repetición de beta-transducina (Ligasa E3).
La proteína β-catenina citoplasmática se degrada constantemente por la acción del complejo proteico destructor, el cual se encuentra compuesto por a) proteínas de andamiaje (Axin [inhibidor de Axis]; la proteína APC, producto del gen supresor de tumores Adenomatous polyposis coli); b) las proteínas quinasas (caseína quinasa 1 a [CK1β]; la glucógeno sintasa quinasa 3b [GSK3β]). CK1α inicia un conjunto de fosforilaciones en secuencia de la β-catenina, con la fosforilación del aminoácido serina en posición 45 y, posteriormente, GSK3β lo hace en los residuos serina en las posiciones 33 y 37, y en la treonina 41. La fosforilación secuencial de la región amino terminal de la β-catenina atrae a la proteína ubiquitina ligasa E3 βTrC, la cual inicia un proceso de ubiquitinación de la β-catenina y su posterior degradación en el proteosoma (Figura 3).
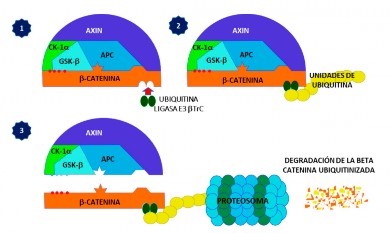
Figura 3 Destrucción de la proteína β-catenina citoplasmática por la acción del complejo proteico destructor. 1) El complejo proteico destructor está compuesto por dos proteínas de andamiaje: Axin y APC, y por las proteínas quinasas CK1α y GSK3b. CK1α fosforila el aminoácido serina en posición 45 de la β-catenina y, posteriormente, GSK3β lo hace en serina en las posiciones 33 y 37, y en la treonina 41. 2) La fosforilación secuencial atrae a la proteína ubiquitina ligasa E3 bTrC, e inicia un proceso de ubiquitinación de la β-catenina. 3) la β-catenina sufre degradación en el proteosoma. Axin (Inhibidor de Axis); APC: supresor de tumores Adenomatous polyposis coli; CK1a: proteína quinasa caseína quinasa 1-a; GSK3b: glucógeno sintasa quinasa 3-b.
βTrC es activado por un correceptor RLP5 o RLP6 (proteína relacionada al receptor de LDL), que usualmente permite que βTrC transfiera péptidos ubiquitinados a la β-catenina. Todo este proceso implica una baja concentración de la β-catenina a nivel citoplasmático y nuclear, por tanto, los factores nucleares TCF/LEF (factor celular T/factor de realce linfoide), que son proteínas promotoras de transcripción genética, son reprimidos por agentes supresores como Groucho 4,14,38-41 (Figura 4).
Vía Wnt/β-catenina activa
Cuando la vía de señalización se activa por la unión del ligando Wnt con el receptor Fzd y la proteína LRP5 o LRP6, formando un complejo ternario en la membrana extracelular, se activan quinasas que inducen la fosforilación de residuos de serina en los dominios intracelulares de LRP5/6, lo que resulta en el inicio de la cascada de señalización de Wnt. En consecuencia, la fosfoproteína Dvl se asocia a la membrana plasmática, por lo que atrae a las proteínas Axin y GSK-3 β del citoplasma, disociando el complejo de destrucción e interrumpiendo la destrucción de β-catenina. La β-catenina aumenta en el citoplasma y en el núcleo celular. A nivel nuclear forma un complejo con TCF/LEF en el cual la β-catenina actúa como activador de la transcripción, y desplaza a Groucho y recluta coactivadores como la proteína nuclear que se une a proteínas de unión, ligadas a elementos de respuesta a AMPc (CBP) o su homólogo p300. La señalización WNT mediada por CBP está asociada con la proliferación de células colónicas, y aquella mediada por p300 promueve la diferenciación 4,14,38-41.
Existen varios mecanismos de retroalimentación negativa para evitar una señalización excesiva de Wnt, así, por ejemplo, el gen AXIN2 (también conocido como conductin) ayuda a aumentar la actividad del complejo de destrucción citosólica y produce una disminución de transcripción mediada por β-catenina 42; la inducción de la expresión de las ligasas E3 unidas a la membrana ring finger protein 43 (RNF43) y zinc and ring finger protein 3 (ZNRF3) median la ubiquitinación de los receptores Wnt, lo que impulsa su internalización y degradación lisosomal 43,44. Sin embargo, dentro de la célula madre adulta, esta retroalimentación negativa es contrarrestada localmente por la familia de proteínas R Spondin, que forma un complejo con el receptor acoplado a la proteína G que contiene repeticiones ricas en leucina (LGR) y RNF43 o ZNRF3, lo que promueve su degradación lisosomal 45 (Figura 4).
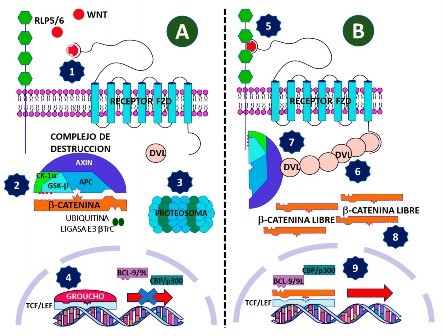
Figura 4 Actividad de la vía Wnt/β-catenina. A. Vía inactiva: 1) el ligando WNT no llega a formar el trímero de activación de la vía, por tanto, la proteína DVL no se asocia a la parte intracitoplasmática del receptor FZD y permanece inactiva. 2) El complejo de destrucción no migra hacia la membrana celular y atrae a la β-catenina, que es fosforilada por CK-1a y GSK3-b, activando la ligasa E3 bTrC. 3) La β-catenina es llevada al proteosoma, donde se destruye. 4) En el núcleo el factor de transcripción TCF/LEF está inactivo por el bloqueo de su actividad por la proteína Groucho, como consecuencia, no se produce activación de los genes derivados de la activación de esta vía. B. Vía activa: 1) el ligando WNT se une al receptor FZD y provoca un cambio de conformación. 5) El cambio conformacional permite su acople al correceptor RLP5/6 y la activación de la parte intracitoplasmática del receptor FZD, lo cual promueve su acoplamiento con DVL. 6) DVL sufre polimerización y secuestro del complejo de destrucción hacia la membrana celular. 7) CK-1a y GSK3-b fosforilan la cola intracitoplasmática de RLP5/6, y permanece allí. 8) Aumenta la β-catenina libre en el citoplasma, desplazándose al núcleo celular. 9) Groucho es desplazado por la β-catenina, lo que permite el acople de β-catenina al factor de transcripción TCF/LEF y el adose de BCL9/9L y CBP/p300, reguladores transcripcionales, con lo cual se inicia las transcripciones genéticas respectivas. Axin (Inhibidor de Axis); APC: supresor de tumores Adenomatous Polyposis Coli; CK1a: proteína quinasa caseína quinasa 1-a; GSK3b: glucógeno sintasa quinasa 3-b. RLP5/6: proteínas relacionadas al receptor 5 o 6 del LDL. DVL: proteina Dishevelled; TCF/LEF: factor celular T/factor de realce linfoide; BCL9/9L: B-cell CLL/lymphoma 9 protein; CBP/p300: CREB-binding protein.
Mutaciones genéticas en la via Wnt/β-catenina en cáncer colorrectal
El CCR es una enfermedad heterogénea en la que podemos encontrar diversas anomalías en las vías de transducción de señales sensibles, como Notch, TGFβ-Smads, Hedgehog, JAK-STAT, Ras-MAPK, PI3K-Akt, Wnt, p53, o procesos de reparación del ADN alterados, los cuales desempeñan un papel importante en el inicio y la progresión del CCR 46. En particular, la vía de señalización Wnt es importante en el desarrollo de la carcinogénesis del CCR, ya que esta vía está muy activa a nivel de las células basales de las criptas intestinales, donde su influencia es crucial para el mantenimiento de las células madre y la homeostasis tisular; por tanto, se espera que la activación aberrante de esta vía sea fundamental en la oncogénesis del cáncer colorrectal.
Las células tumorales en el CCR muestran acumulación nuclear de β-catenina y características de la transición epitelio-mesénquima (EMT) asociadas con activación local de la vía de señalización Wnt; sin embargo, es preciso tener en cuenta que la diseminación de las células cancerosas no se debe únicamente a los cambios genéticos, sino que también es importante el microambiente tumoral donde la matriz extracelular, factores de crecimiento y factores inflamatorios juegan un papel clave en la invasión tumoral 47.
El grupo de estudio del Atlas Genómico del Cáncer (TGCA) realizó un análisis del genoma completo en 96 muestras de cáncer colorrectal, y encontró 24 genes con mutaciones significativas sobre todo en los genes APC, TP53, SMAD4, PIK3CA y KRAS, ARID1A, SOX9 y FAM123B. La vía de señalización Wnt se alteró en el 94 % de todos los tumores (224 muestras), incluida la inactivación bialélica de APC o las mutaciones activadoras de CTNNB1 en el 80 % de los casos. Asimismo, el receptor Frizzled (FZD) estuvo sobrexpresado en 17 % de las muestras. En total, se encontraron 16 diferentes genes alterados de la vía Wnt, lo que refuerza la importancia de esta vía en CCR 48.
La evaluación de la influencia de cada variante del ligando Wnt con relación al CCR puede ser compleja y heterogénea; se han explorado diversos estudios preclínicos y clínicos tempranos, y se ha logrado demostrar que existen diferentes influencias sobre la vía de transducción respecto de las funciones de estos ligandos durante la tumorigénesis del CCR humano y que pueden estar asociadas tanto a la ruta canónica como a la no canónica 49.
Mutaciones en el gen APC
La poliposis adenomatosa familiar (FAP) es el resultado de mutaciones en la línea germinal del gen APC. Generalmente, las mutaciones somáticas de APC de la FAP colónica se encuentran en la región llamada "grupo de mutaciones colorrectales" (MCR, codones 1286 a 1581) y son los más adecuados para generar un nivel definido de señalización Wnt que favorece el crecimiento tumoral. También es importante señalar que, en los pacientes con FAP, el sitio de la mutación de la línea germinal determina el tipo de mutación somática (segundo golpe) de los pólipos colorrectales. Aproximadamente el 76 % del CCR esporádico tiene mutaciones en al menos un alelo que comprende un truncamiento de MCR 50. A pesar de que podemos encontrar la proteína APC incompleta, truncada por mutaciones en ambos alelos del gen correspondiente, en el inicio del CCR, la función de estas APC alteradas no está totalmente inactiva. Una explicación del modelo de señalización "suficientemente correcta" es que la eliminación de todos los sitios de unión a β-catenina termina siendo menos favorable para la célula tumoral, porque la activación constitutiva de la vía de β-catenina conduce a cambios extensos en la regulación génica que resultan en un mayor riesgo de muerte celular. Por el contrario, retener algunos de los sitios de unión de β-catenina puede permitir una regulación negativa parcial, lo que proporciona una actividad de señalización de Wnt óptima que es suficiente para conferir una ventaja de proliferación sin inducir la muerte celular 50.
La pérdida de la actividad por mutaciones en el gen APC puede ser compensada parcialmente por la acción de su homologo APC2, ya que su acción es mucho más débil. Una parte importante de pacientes con APC mutado en CCR muestra hipermetilación del promotor APC2, lo que puede indicar que el silenciamiento del gen por esta acción epigenética podría proporcionar una ventaja en el crecimiento tumoral 51.
Mutaciones en los genes AXIN
Las mutaciones AXIN1 y AXIN2 son detectadas en una pequeña fracción del CCR, sin embargo, el mecanismo más relevante puede ocurrir a través de su ubiquitinación y posterior destrucción por aumento de la actividad transcripcional de RNF146, una ubiquitina ligasa E3 que promueve la destrucción de AXIN1 y AXIN2, y de esa forma genera proliferación del CCR a través de la liberación y activación de la β-catenina. La sobreexpresión de RNF146 ocurre en el 6 % de los casos de CCR y se correlaciona con un mal pronóstico 52.
Otras mutaciones relevantes
La inactivación de los genes RNF43 y ZNRF3 es muy importante para el crecimiento tumoral, de tal forma que en los CCR aserrados humanos, a menudo, ambos genes están alterados de diversas formas. En el cáncer humano, el espectro mutacional de RNF43 está dominado por mutaciones truncadas y sin sentido, mientras que el ZNRF3 se ve afectado con mayor frecuencia por mutaciones sin sentido y deleciones. Los mecanismos de estas alteraciones pueden ser múltiples 53.
En estudios previos con animales se encontró que una sola mutación CTNNB1 impulsa la tumorigénesis en el intestino delgado, pero es insuficiente para inducir la transformación en el colon; el efecto discrepante en el intestino delgado y el intestino grueso se explica por una mayor expresión de E-cadherina que ancla el exceso de β-catenina a la membrana celular y, por lo tanto, compensa el efecto oncogénico de las mutaciones CTNNB1 heterocigotas en el colon. Sin embargo, el efecto supresor de tumor de la E-cadherina se supera en presencia de mutaciones homocigóticas del gen CTNNB1, lo que indica que la superación del umbral de los niveles de β-catenina mutada también juega un papel en la neoplasia colorrectal.
Blancos terapéuticos en la vía Wnt/β-catenina
La vía de señalización Wnt/β-catenina está frecuentemente alterada en las células cancerosas, sobre todo en el contexto del CCR, lo que confiere a las células características similares a las células madre, como autoduplicación, potencial diferencial y transición epitelial a mesenquimal, lo cual trae como consecuencia el desarrollo de una enfermedad más agresiva, con la consiguiente posibilidad de metástasis y una mayor resistencia a quimioterapia o radioterapia. El estudio permanente de las alteraciones de la señalización de Wnt/β-catenina ha abierto nuevas ventanas de investigación en el desarrollo de nuevas drogas que puedan contrarrestar la actividad oncogénica y posterior desenlace en el CCR (Figura 5). Esta visión se ve reforzada por el hecho de que, en el CCR en ratones, la restauración de la expresión de APC impulsa la diferenciación de las células tumorales y la restauración de la homeostasis del tejido normal 54. Sin embargo, a pesar de que se han explorado muchas moléculas en esta vía, aún no se ha logrado la aprobación comercial de ninguna droga. Dado que es una vía tan compleja, que interviene en procesos celulares muy importantes para la homeostasis, constituye un gran desafío para la ciencia buscar medicamentos que inhiban las señales de la vía Wnt en las células cancerosas, y evitar una toxicidad prohibitiva en el resto del organismo.
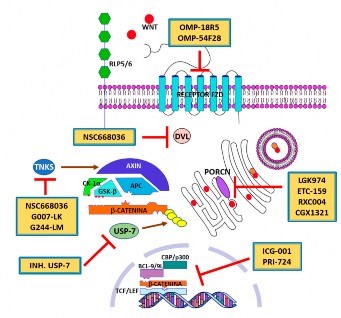
Figura 5 Principales blancos terapéuticos en la vía Wnt/β-catenina. Proteínas relacionadas al receptor 5 o 6 del LDL; Axin (inhibidor de Axis); APC: supresor de tumores Adenomatous Polyposis Coli; CK1β: proteína quinasa caseína quinasa 1-β; GSK3β: glucógeno sintasa quinasa 3β. RLP5/6: proteínas relacionadas al receptor 5 o 6 del LDL. DVL: proteína Dishevelled; TCF/LEF: factor celular T/factor de realce linfoide; BCL9/9L: B-cell CLL/lymphoma 9 protein; CBP/p300: CREB-binding protein: PORC: proteína porcupine; USP-7: proteasa específica 7 de ubiquitina. TNKS: tanquirasa.
Los anticuerpos monoclonales OMP-18R5 y OMP-54F28 interactúan con los receptores Fzd y bloquean la vía de señalización Wnt canónica a través del impedimento de los múltiples ligandos de la familia Wnt. Lamentablemente, la eficacia y la toxicidad de estos agentes no ha sido la mejor y se espera que las terapias combinadas tengan un mejor resultado 55.
Dishevelled (Dsh) es una familia de proteínas involucradas en las vías de señalización de Wnt canónicas y no canónicas. Dsh (Dvl en mamíferos) es una fosfoproteína citoplasmática que es reclutada a la membrana atrayendo a Axin y GSK3β, lo cual previene la degradación de β-catenina. NSC668036 es un pequeño inhibidor orgánico de Dvl, que suprime la transcripción génica impulsada por la β-catenina a través de la formación de un complejo con Dvl al unirse a su dominio PDZ. Esta droga aún no ha sido probada en CCR 56.
XAV939 es un poderoso potenciador de la actividad del complejo de destrucción, al promover la estabilización de AXIN en líneas celulares de CRC con mutación deAPC. XAV939 inhibe la PARilación (incorporación de la poli-ADP-ribosil) de las enzimas tankirasa (TNKS1 y TNKS2), lo cual impide la PARilación de AXIN1 y 2, y su posterior ubiquitinación y degradación mediada por RNF146. Lamentablemente, los estudios clínicos revelaron resultados conflictivos, debido, posiblemente, a la resistencia por otras alteraciones oncogénicas concurrentes en otras vías como KRAS, pero también se demostró que las dosis elevadas con inhibidores de TNKS provoca toxicidad intestinal, pérdida de peso y muerte en roedores. Sin embargo, la aplicación de dosis reducidas de los inhibidores de TNKS en combinación con otros regímenes de tratamiento revelaron respuestas antitumorales prometedoras 57,58. Existen nuevos inhibidores de tankirasa, G007-LK y G244-LM, que reducen la señalización de Wnt/β-catenina impulsada por mutaciones de APC, en aproximadamente un 50 %, en la mayoría de las líneas celulares de CRC 59.
La proteasa especifica de ubiquitina 7 (USP7) es una de las enzimas desubiquitinizantes que destruye la ubiquitina, protegiendo así a la proteína substrato de la degradación. De acuerdo con ello, la β-catenina puede desubiquitizarse y estabilizarse mediante la acción de la USP7 en el CCR mutado en APC, pero no el CCR con el gen APC no alterado. El mecanismo propuesto para esta actividad es que el dominio inhibidor de la β-catenina (CID) del APC no mutado protege la β-catenina de la desubiquitinación mediada por USP7, mientras que la APC que carece de CID expone la β-catenina a la USP7 para su desubiquitinación. Por lo tanto, la inhibición de USP7 en CRC con mutación del APC suprime la activación de Wnt al recuperar la ubiquitinación de β-catenina, lo que conduce a la diferenciación celular e inhibe el crecimiento tumoral 60. Estudios con inhibidores de la USP7 lograron suprimir la activación de Wnt en líneas celulares con APC mutado en CCR in vivo 61; sin embargo, estos hallazgos fueron cuestionados por otro estudio, en el cual los inhibidores de la USP7 fueron identificados como reguladores positivos de la vía Wnt/β-catenina mediante la inhibición de la estabilización de AXIN 62.
Los inhibidores de PORCN (enzima porcupine) se utilizan para interrumpir la señalización de Wnt al bloquear la secreción de proteínasWnt mediantela inhibiciónde unaenzimabiosintética clave. Existen hasta cuatro moléculas en actual investigación (LGK974, ETC-159, RXC004 y CGX1321), las cuales están siendo exploradas en estudios clínicos incipientes. A pesar de los prometedores resultados preclínicos, los efectos adversos de los inhibidores de PORCN, que incluyen pérdida ósea y disgeusia, posiblemente limiten la dosis a la que se pueden aplicar en la clínica 63. GSK-3β es una parte importante del "complejo de destrucción" en la vía Wnt/β-catenina. Pequeñas moléculas de GSK-3β están siendo usadas en melanoma maligno con un relativo éxito, sin embargo, no han funcionado igual en CCR 64.
La interacción TCF/β-catenina puede ser bloqueada eficazmente por moléculas pequeñas en el contexto nuclear, en células de CCR in vitro, por lo cual tienen un potencial grande para el tratamiento de esta neoplasia maligna. Existen múltiples substancias con estas características: PKF115-584, CGP049090 y PKF222-815, inhibidores de la transcripción de repuesta a la catenina: PNU-74654, 2,4-diamino-quinazolina, LF3, CWP232228, etc.; sin embargo, todas se encuentran en estudios preclínicos y se espera su llegada a los primeros estudios clínicos para evaluar su valía dentro del CCR. Algunas de las moléculas que bloquean la vía Wnt/β-catenina están en fases clínicas incipientes y pueden seguirse a través del portal ClinicalTrials.gov de la Biblioteca Nacional de Medicina de los Estados Unidos (Tabla 1).
Tabla 1 Principales inhibidores de la vía Wnt/β-catenina asociados a los protocolos de investigación de la Biblioteca Nacional de Medicina de los Estados Unidos. NCT: número del estudio clínico. Modificado de Zhong Z, Cancer. Mol Pharmacol. 2020 Feb;97(2):72-89 (65).
| Blanco terapeútico | Droga | NCT | Fase |
|---|---|---|---|
| Anti-Frizzled | Vantictumab | 1345201, 2005315, | I |
| 1,2,5,7,8 | (OMP-18R5) | 1957007, 1973309 | I |
| Anti-Wnt receptor | Ipafricept | 2069145, 2092363, | I |
| (OMP-54F28) | 2050178, 1608867 | I | |
| Inhibidor PORCN | ETC-159 | 2521844 | I |
| (bloqueo de la | RXC004 | 3447470 | I |
| secreción de Wnt) | CGX1321 | 3507998, 2675946 | I |
| LGK974 | 1351103, 2278133 | I-II | |
| Bloqueo interacción | PRI-724 | 1764477, 1302405 | I-II |
| β-catenina/CBP | 1606579, 2413853 | I-II |
También se han investigado antagonistas de coactivadores transcripcionales, los cuales interrumpen la interacción entre β-catenina y estos cofactores como CBP y BCL9. ICG-001, un antagonista de la interacción de β-catenina/CBP, ha demostrado que disminuye el crecimiento de las células de CRC en ratones con mutación de uno de los alelos de APC, al unirse específicamente a CBP, sin interactuar con su homólogo p300, el cual controla el cambio fundamental de células madre y/o progenitoras. PRI-724, otro agente antagonista específico de CBP/catenina de segunda generación, ha demostrado acciones similares a ICG-001, con un perfil de toxicidad aceptable 66.
Conclusiones
El CCR fue uno de los primeros tumores para el que se construyó un modelo de carcinogénesis de forma secuencial y a través de la acumulación de distintos eventos genéticos y epigenéticos. Las alteraciones que ocurren en la vía Wnt causan distintas enfermedades, como osteoporosis, artritis reumatoide y cáncer, por lo cual es un objetivo terapéutico importante realizar la búsqueda de nuevas moléculas selectivas e inteligentes. No hay duda de que la vía canónica (Wnt/β-catenina) es crítica en la génesis del CCR, ya que se han encontrado múltiples cambios genéticos y epigenéticos que alteran diversos componentes de la vía de señalización canónica WNT, sobre todo a nivel del gen APC. Los avances en la investigación biomolecular nos han proporcionado nuevos caminos para situar esta vía de señalización como un objetivo terapéutico importante, por lo cual existen muchas drogas en diferentes fases de la investigación. Lamentablemente, ninguna de ellas ha sido aprobada para su uso comercial en el CCR hasta la fecha.

