










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.11 n.2 Lima jul./dic. 2004
José Sandoval, Bedsabé Delgado, Luis Rivas, Bertha Bonilla, Daniel Nugent y Ricardo Fujita*
(*) Instituto de Genética y Biología Molecular, Facultad de Medicina, Universidad de San Martín de Porres.
Presentado: 23/07/2004
Aceptado: 29/12/2004
Resumen
Los polimorfismos del ADN mitocondrial son herramientas en el estudio comparativo de poblaciones modernas y antiguas. Entre los más usados están los haplotipos mitocondriales basados en RFLP (polimorfismo de longitud de fragmentos de restricción) y un sistema de inserción /deleción. El presente estudio establece la frecuencia de estos haplotipos y compara un total de 144 individuos representativos de las islas Taquile y Amantaní (lengua quechua) y de las islas de Los Uros y Anapia (lengua aymara) del lago Titicaca, Perú. Nuestros resultados revelan la predominancia del haplotipo B1: 100% en Taquile (n=57); 88,6% en Amantaní (n=35); 75% en Los Uros (n=28) y 87,5% en Anapia (n=24), siendo las frecuencias más altas registradas en el mundo. Otros haplotipos se observan en menor proporción: 17,9% de A2 y 7,1% de D1 en Los Uros; 11,4% de la variante C1 en Amantaní; 4,2% de cada haplotipo C1, C2 y D1 en Anapia. La alta frecuencia de B1 indica que las poblaciones de Taquile, Amantaní y Anapia provienen de un grupo fundador reducido. Aunque hay afinidad entre las poblaciones aymaras de Anapia y Los Uros; la proporción de algunos alelos en los últimos, sugiere la persistencia de un acervo genético uru en contraposición a la idea de su extinción.
Palabras clave: ADN mitocondrial, isleños del Lago Titicaca, haplotipos, RFLP.
Abstract
We analyzed mitochondrial DNA haplotypes from 144 samples of islanders of the Taquile and Amantani (Quechua speakers) and Los Uros and Anapia (Aymara speakers) of the Lake Titicaca, Peru. We have found the highest frequency of B1 mtDNA haplotype ever reported: 100% in Taquile (n= 57); 88,6% in Amantani (n= 35); 87,5% in Anapia (n= 24) and 75% in Los Uros (n= 28). There are other haplotypes but in low frequencies: 11,4% of C1 in Amantani; 17,9% of A2 and 7,1% of D1 in Los Uros; 4,2% of C1, C2 and D1 haplotypes in Anapia. The high frequency of the B1 haplotype indicates that Taquile, Amantaní and Anapia islands have been populated by a reduced founder group. Although the genetic affinity between islanders of Anapia and Los Uros, the proportion of some alleles in the latter suggest the persistence of the uru genetic background.
Keywords: mitochondrial DNA, Lake Titicaca islanders, haplotypes, RFLP.
Introducción
La composición étnica en el Perú está formada por poblaciones de origen nativo, europeo, africano y asiático. En la actualidad se observa un mestizaje generalizado, sin embargo aún quedan algunas poblaciones homogéneas que posibilitan el estudio de sus orígenes. Las migraciones y el aislamiento geográfico a través de los siglos, han sido los factores determinantes en la variabilidad interpoblacional, así como en la homogeneidad genética dentro de cada una de ellas. El origen de muchas comunidades nativas actuales es desconocido y de ello solamente se cuentan con teorías basadas en sus tradiciones orales y crónicas o registros llevados por los conquistadores españoles a partir del siglo XVI (Bouysse-Cassagne, 1991).
Actualmente, los pobladores peruanos de las islas del lago Titicaca y en general de la región del Collao, hablan quechua y aymara. Sin embargo, las crónicas de la conquista española y la colonia registraron además las lenguas uruquilla y puquina. En aquella época había un grupo humano denominado uru o huma haque (gente del agua) en aymara; ellos hablaban uruquilla y habitaban las orillas del lago Titicaca, dedicándose a actividades primarias como pesca, caza y recolección. Hasta el siglo XVI, los urus eran aproximadamente el 25% de la población del Collao. Se distribuían por un eje acuático formado por el río Azángaro, lago Titicaca, río Desaguadero en Perú; lago Poopó, río Lacajahuira y el lago Coipasa en Bolivia. Se sabe además, que en esta época habían pescadores urus en la jurisdicción de Arequipa, en el desierto de Atacama y en la costa de Arica e Iquique (Manelis de Klein, 1973; Wachtel, 1986).
Desde la época de la conquista hasta los tiempos actuales se ha percibido un descenso de la población uru, e incluso hay investigadores que sugieren la casi desaparición de este grupo del territorio peruano (Manelis de Klein, 1973; Gran Enciclopedia del Perú, 1998). Otros autores sugieren que paulatinamente, los urus fueron asimilados por el sistema colonial y republicano perdiendo las características de su cultura, muchos de ellos «aymarizándose» (Wachtel, 1986). Estos antecedentes conllevan a pensar que los habitantes de las islas Los Uros mantendrían todavía parte de su acervo genético proveniente de los urus originales.
En otro caso, en las islas Taquile y Amantaní hay evidencias arqueológicas e históricas de poblamientos sucesivos desde épocas preincaicas, incaica y colonial. En ellas se han encontrado restos de la cultura Tiahuanaco; luego habitada por mitimaes durante el Tahuantinsuyo. Según las crónicas españolas, la península de Capachica, islas Amantaní, Titicaca (isla del Sol) y Coati (isla de la Luna) fueron ocupadas por los collas considerados «los tejedores del Inca» (Bouysse-Cassagne, 1991) (precisamente, el arte textil es muy significativo en Taquile y Amantaní). Después de 1650 las islas Taquile y Amantaní fueron adquiridas por el encomendero español Pedro Gonzales de Taquila, trayendo consigo gente de lugares no determinados (Espinoza-Avendaño, 1989). Algunas crónicas relatan que Amantaní fue evacuada antes de ser repoblada por su hacendado; no hay referencia sobre Taquile, pero se asume que sufrióla misma suerte. En las tradiciones orales de los taquileños hay coincidencias sobre el hecho del repoblamiento, pero difieren en el grupo humano que lo hizo: i) original (antiguos isleños), ii) ribereños del lago, (Espinoza-Avendaño, 1989; Bouysse-Cassagne, 1991).Taquile y Amantaní se mantuvieron relativamente aisladas hasta 1960, por lo cual podemos asumir que la población es en su mayoría descendiente del repoblamiento del siglo XVII.
Por otro lado, en la isla Anapia-Suana (ubicada en el Lago Menor o Wiñaymarca) e islas cercanas como Suriki, Pariti, Anayutani, así como en Titicaca y Coati, también se han encontrado vestigios de las culturas Tiahuanaco e Incaica (Bouysse-Cassagne, 1991). Desde tiempos remotos los antiguos pobladores del lago Menor se dedicaban a la agricultura, ganadería y la pesca. En el siglo XV, la mayor parte de esta región era habitada por los collas (de lengua aymara), quienes fueron sometidos por el Inca Pachacutec. Luego, en la época del colonialismo europeo, las islas de Wiñaymarca fueron utilizadas como haciendas españolas, y posteriormente devueltas a sus comunidades.
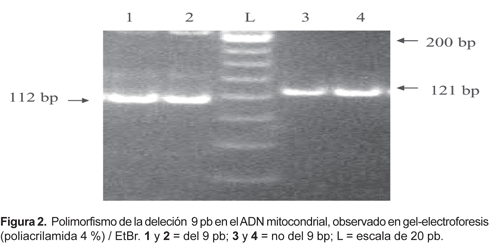
El parentesco genético puede ser determinado por el uso de polimorfismos de ADN estándar que permiten comparar datos obtenidos por diferentes grupos de investigadores (Merriwether et al., 1996; Simoni et al., 2000). Los polimorfismos de ADN más utilizados en estudios de poblaciones son los del ADN mitocondrial (ADNmt). Estos son revelados por la presencia o ausencia de sitios de restricción (RFLPs, polimorfismo de longitud de fragmentos de restricción) específicos en varios puntos del ADNmt y por la inserción/deleción de 9 pares de nucleótidos (Fig. 1, 2 y 3).
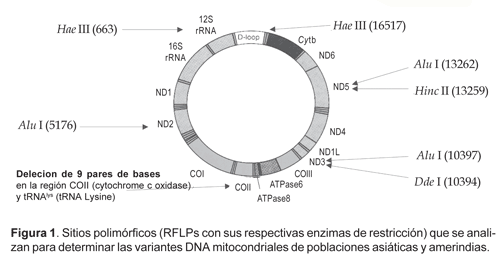
En la mayoría de los ADN mitocondriales de las poblaciones nativas americanas prevalecen 8 haplotipos: A1, A2, B1, B2, C1, C2, D1 y D2 del genoma mitocondrial. La frecuencia de cada tipo varía en las diferentes poblaciones; siendo B (B1) el más frecuente entre los quechuas y aymaras, asi como en un estudio de otros nativos peruanos (Merriwether et al., 1996; Rodríguez-Delfín et al., 2001).
En el presente trabajo se analizan los haplotipos mitocondriales de poblaciones nativas del lago Titicaca y se da una interpretación de las relaciones entre éstas.
Materiales y métodos
Luego del apropiado consentimiento, se obtuvo muestras de sangre de un total de 151 individuos voluntarios de 10 a 18 años de edad de cuatro islas del lago Titicaca. El muestreo sanguíneo se efectuó a fines de 1998. Diecinueve muestras del Centro Educativo Particular (C.E.P.) 70682 «Uros Torani Pata», 9 muestras del Centro Educativo Superior (C.E.S.) Técnico Artesanal «Uros Chulluni»; 41 muestras del C.E.S. Artesanal Taquile, 20 muestras de la Escuela Particular Nuestra Señora de Los Campos-Taquile, 23 muestras del C.E.S. Agro Artesanal «Miguel Grau» Amantaní, 13 muestras del C.E.P. 70037 Amantaní; y 26 muestras del C.E.S. Anapia. La sangre fue guardada en congelación para su transporte y conservación.
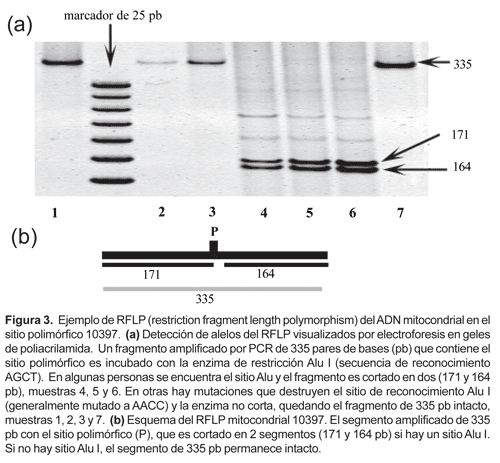
La extracción de ADN de las muestras de sangre congelada se efectuó por una técnica simplificada sin fenol-cloroformo (Fujita, 1991). La reacción de amplificación PCR (polymerase chain reaction) se realizó en microtubos dentro del termociclador Thermolyne Amplitron II. Los parámetros de PCR han sido optimizados según protocolos estándar con los oligonucleótidos (primers) específicos para la amplificación de ADNmt: L635/H708; L5054/H5189; L13257/H13393; 10235-10254/10569-10550; 16287-16306/16547-16527 (Sukernik et al., 1996; Merriwether et al., 1996; Easton et al., 1996; Stone & Stoneking, 1998). Para obtener los patrones de RFLP, los productos PCR fueron digeridos con sus respectivas enzimas de restricción (Fig. 1) según especificaciones del protocolo del fabricante (Corporación Promega). El polimorfismo de la deleción/inserción de 9 pares de bases (Fig. 2) se determina directamente por amplificación PCR con los primers L8215/H8297 (Stone & Stoneking, 1998). Los productos de PCR y RFLP fueron separados por electroforesis en geles de agarosa (2%) y poliacrilamida (4%), teñidos en solución de bromuro de etidio, analizadas con luz UV y registradas digitalmente con fotodocumentador BioDoc Systems, UVP (Fig. 3).
La distribución de frecuencias de subtipos de los haplogrupos ADNmt se indican directamente en porcentajes y para la determinación del índice de diversidad genética (h) se utilizó la fórmula de Nei (1987):
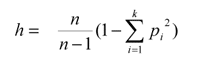
n= número de muestras,
k= número de haplotipos,
pi= frecuencia del haplotipo.
Resultados
Máxima frecuencia de haplotipo B1 y evidencia de efecto fundador
El presente estudio encontró diferencias en la distribución de frecuencias de haplotipos del ADN mitocondrial en las poblaciones de las diferentes islas (Tabla 1).
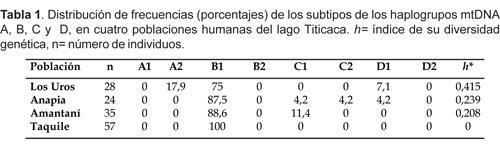
Distribución de otros alelos
El haplotipo A2 (17,9%) ha sido encontrado solamente en las islas Los Uros. La variante C1 está presente en Amantaní (11,4%) y en Anapia (4,2%) y en ésta última, además se observa C2 (4,2%). El haplotipo D1 se observa tanto en Los Uros (7,1%) como en Anapia (4,2%).
Discusión
La distribución de las poblaciones nativas actuales obedece a desplazamientos ocurridos durante miles de años en el territorio peruano. Sólo después de la llegada de los españoles tenemos registros de éstos, siendo necesario recurrir a otras fuentes para discernir acerca de las migraciones en la era prehispánica. Elementos antropológicos y sociológicos como la lingüística, las tradiciones culturales y orales nos ayudan a estudiar los orígenes de las diversas etnias.
Actualmente, es posible también establecer patrones genéticos individuales y poblacionales estudiando variantes en el ADN. Así, en estudios de poblaciones modernas y antiguas, las variantes polimórficas en el ADN mitocondrial son bastante usadas para deducir orígenes y desplazamientos de sus ancestros (Merriwether et al., 1996; Lell et al., 2002).
Se han propuesto posibles rutas migratorias para poblar Sudamérica y algunas incluyen dos subgrupos, uno migrando por la costa del Océano Pacífico y poblando las zonas andinas (mayormente portadores del haplogrupo B) y otro por la región Amazónica (en general portadores de haplogrupos A, C y D) (Greenberg, 1987; Merriwether et al., 1996; Rodríguez-Delfin et al., 2001). Nuestros resultados sobre el estudio de ADNmt en pobladores de las islas del lago Titicaca concuerdan con la predominancia del subtipo B1 en las poblaciones andinas. Cabe destacar que observamos la frecuencia más alta de B1 que en cualquier estudio previo de poblaciones nativas americanas: en Taquile (100%), Amantaní (88,6%), Anapia (87,5%) y en los Uros (75%). Por otro lado, en este estudio, la variante A2 (5 de 28 muestras) sólo ha sido encontrada en las islas Los Uros. Esto sugeriría un origen distinto de algunos de sus ancestros de estas islas con respecto a los de Taquile, Amantaní y Anapia. La heterogeneidad en la población de las islas Los Uros sería consecuencia de la mixtura con otros grupos del altiplano, ocurridas desde épocas preincaicas. Así gradualmente, los urus fueron asimilándose al grupo cultural de sus vecinos aymaras. Además, estudios lingüísticos y de grupos sanguíneos indican que los urus, estarían emparentados con la familia lingüística Arawak (diseminados en la selva de Paraguay, regiones de Ucayali-Madre de Dios-Cuzco) y tribus caribeñas, por lo que pertenecerían a un grupo étnico diferente a los aymaras y quechuas (Wachtel, 1986; Torero, 1992).
El grado de parentesco genético entre los cuatro grupos isleños está asociado más a las afinidades lingüísticas que a la cercanía geográfica. Por ejemplo, hay mayor afinidad entre grupos geográficamente alejados como Los Uros-Anapia (ambos aymara hablantes) que entre grupos cercanos como Los Uros-Taquile (aymara y quechua respectivamente).
Así, se encuentra una gran homogeneidad entre los habitantes de las islas Taquile y Amantaní de habla quechua, indicando que genéticamente son afines, acorde con su cercanía geográfica. Las altas frecuencias del subtipo B1 (100% en Taquile) sugieren que ambas poblaciones descienden de un pequeño grupo fundador que apareció probablemente en el repoblamiento del siglo XVII.
En cambio, entre los pobladores de las islas Los Uros y Anapia de lengua aymara, hay heterogeneidad que concuerda con los antecedentes históricos de la «aymarización» lingüística de las diferentes etnias del Collao ocurrida desde épocas preincas hasta el final de la colonización española. Existe la hipótesis de que la etnia uru se extinguió de las islas de Los Uros desde hace muchas décadas y que los isleños actuales serían o descenderían de colonos recientes de otras etnias ribereñas (Vellard, 1959; Gran Enciclopedia del Perú, 1998). En contraposición a a esta idea, nosotros creemos que estos isleños todavía conservan un fuerte bagaje genético uru bajo una influencia cultural aymara. Por ejemplo, en esta población hay una alta proporción del alelo A2 (17,9%) comparada con 0% de otras islas (presente estudio), 0% de poblaciones aymaras de Perú y Chile (Merriwether et al., 1996) ó a 3.7% de una mezcla de aymaras y quechuas (Rodríguez-Delfín et al., 2001). Por otro lado, cabe notar que A2 se encuentra en mayor proporción en poblaciones amazónicas (de donde se originarían los urus) que en las poblaciones andinas (Rodríguez-Delfín et al., 2001). De manera similar el estudio de las frecuencias del gen MBL (mannose binding lectin) en isleños del lago Titicaca también muestra diferentes proporciones entre los habitantes de la isla de Los Uros y los de las otras islas del lago (Sandoval et al., 2002). Además no se debe descartar un mestizaje de urus con los ribereños en una proporción a determinar en futuros trabajos. La realización de un estudio comparativo con otras poblaciones urus de Bolivia, las ribereñas vecinas en el lago y las de los grupos amazónicos emparentados, serviría para descartar o comprobar la extinción de la etnia uru del Perú.
En una comparación global, se notan diferencias moderadas entre los pobladores quechua hablantes de Taquile y Amantaní con los pobladores de las islas Los Uros y Anapia que hablan aymara, confirmando que una parte de sus ancestros son de origen distinto.
Comparado con estudios en otras poblaciones humanas, podemos afirmar que cada isla todavía mantiene un relativamente alto grado de homogeneidad genética. Esto estaría causado por el establecimiento de un reducido grupo ancestral y por el relativo aislamiento geográfico que ofrecen las islas (efecto fundador). Estudios comparativos de la distribución de frecuencias de haplotipos ADNmt de diversas poblaciones nativas sudamericanas muestran afinidad entre los pobladores de Los Uros y poblaciones amazónicas. Esto concuerda con los estudios lingüísticos y de grupos sanguíneos que sugieren el origen amazónico de los urus (Wachtel, 1986; Torero, 1992).
Los datos históricos, complementados con las particularidades de los marcadores polimórficos de ADN, nos permiten sugerir que los habitantes de las islas Los Uros conservan todavía un importante componente genético uru dentro de su población.
Las diferencias, aunque moderadas, entre los pobladores de las islas Los Uros y Anapia, aymara hablantes, nos indicarían tomar con cautela el uso de las diferencias y semejanzas lingüísticas, ya que serían solamente referencias culturales y no un referente étnico biológico en los estudios genéticos de las poblaciones humanas.
Agradecimientos
El presente trabajo se dedica a los habitantes del Lago Titicaca, en especial a los escolares de Taquile, Amantaní, Los Uros y Anapia, quienes entusiastamente participaron en el estudio. Agradecemos el co-financiamiento CONCYTEC (Concurso de Proyectos de Investigación 2000, contrato No. 871-2000-OAL) con la Universidad de San Martín de Porres. Nuestro reconocimiento a los doctores Francisco M. Salzano (Universidade Federal do Rio Grande do Sul, Brasil) y Juan Herrera Carpio (Universidad Nacional del Altiplano, Puno), las autoridades del Hospital Regional MNB, Dirección Regional de Educación, la Capitanía de Puerto de Puno; Concejos Municipales de Mollendo y Puno. A los doctores Armando Guevara y Sonia Guillén, por la valiosa información antropológico-histórico y a todas las personas que de una u otra manera hicieron posible este estudio.
Literatura citada
Bouysse-Cassagne, T. 1991. Poblaciones humanas antiguas y actuales. ORSTOM, HISBL, Bolivia, p. 481-498. [ Links ]
Easton R.D.; A.D. Merriwether; D.E. Crews; and R.E. Ferrell 1996. mtDNA Variation in the Yanomami: Evidence for Additional New World Founding Lineages. Am. J. Hum. Genet. 59:213-225. [ Links ]
Espinoza-Avendaño, J. 1989. Taquile. Historia-Economía-Artesanía. Instituto de Investigaciones para el desarrollo social del altiplano. Convenio UNA-NUFFIC, 148 p. Reimpresión,1990 por Talleres IIDSA, Puno. [ Links ]
Fujita, R. 1991. Localisation du gene de l'ataxie de Friedreich par sa liaison génétique et physique avec les loci D9S5 et D9S15. These de Docteur de l'Universite Louis Pasteur de Strasbourg, France, 80 p. [ Links ]
Greenberg, J.H. 1987. Language in the Americas. Stanford Univ. Press, Stanford, CA. [ Links ]
Lell, J.T.; R.I. Sukernik; Y.B. Starikovskaya; J.L. Su B; T.G. Schurr; P.A. Underhill and D.C. Wallace. 2002. The Dual Origin and Siberian Affinities of Native American Y Chromosomes. Am. J. Hum. Genet. 70:192-206. [ Links ]
Manelis de Klein, H.E. 1973. Los Urus: El extraño pueblo del altiplano. Estudios Andinos, N7, V3, N1:129-150. [ Links ]
Merriwether, D.A.; W.W. Hall; A. Vahlne and R.E. Ferrell 1996. MtDNA Variation Indicates Mongolia May Have Been the Source for the Founding Population for the New World. Am. J. Hum. Genet. 59:204-212. [ Links ]
Nei, M. 1987. Molecular Evolutionary Genetics. Columbia Univ. Press, New York. [ Links ]
Rodriguez-Delfin, L. A.; V. E. Rubin-de-Celis, M. A. Zago. 2001. Genetic Diversity in an Andean Population from Peru and Regional Migration Patterns of Amerindians in South America: Data from Y Chromosome and Mitochondrial DNA. Hum. Hered 51:97-106. [ Links ]
Sandoval J, Madsen HO, Garred P, Fujita R (2002) Mitochondrial and mannose-binding lectin haplotypes in Quechua and Aymara islanders of the Lake Titicaca. Cold Spring Harbor Symposium «Human Origins and Diseases» Abstracts book. pp. 86 [ Links ]
Simoni L.; F. Calafell; D. Pettener; J. Bertranpetit and G. Barbujani. 2000. Geographic Patterns of mtDNA Diversity in Europe. Am. J. Hum. Genet. V.66:262-278. [ Links ]
Stone, A. and M. Stoneking. 1998. MtDNA Analysis of a Prehistoric Oneota Population: Implications for the Peopling of the New World. Am. J. Hum. Genet. V62:1153-1170. [ Links ]
Sukernik, R. I.; T. G. Schurr; E. B. Starikovskaya and D. C. Wallace. 1996. Mitochondrial DNA Variation in Native Siberians, with Special Reference to the Evolutionary History of American Indians. Studies on restriction polymorphism. Genetika, T.32, N3:432-439. [ Links ]
Torero, A. 1992. Acerca de la familia lingüística uruquilla (Uru-Chipaya). Revista Andina, N1, Cusco, p. 171-189. [ Links ]
Wachtel, N. 1986. Men of the water: The Uru problem (XVI and XVII centuries). Cambridge University Press & Editions de la Maison des Sciences de L' Homme, pp.282-310. [ Links ]
Vellard, J. 1959. Poblaciones indígenas del lago Titicaca. Boletín Sociedad Geográfica Lima, 76 (3-4 trimestre): 17-40. [ Links ]
Gran Enciclopedia del Perú. 1998. Editores Lexus, España, p.927. [ Links ]
(*) Instituto de Genética y Biología Molecular, Facultad de Medicina, Universidad de San Martín de Porres. Alameda del Corregidor Cdra. 15, La Molina, Lima, Perú.
Email: José Sandoval jsandoval@usmp.edu.pe
Email: Ricardo Fujita rfujita@amauta.rcp.net.pe
