










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.12 no.1 Lima Jan./jul. 2005
José Antonio Ochoa*
Presentado: 12/09/2004
Aceptado: 29/11/2004
Resumen
Se presenta la distribución geográfica de 24 especies de escorpiones (cinco géneros y tres familias) para el corredor andino en el sur del Perú. Se discuten algunos aspectos zoogeográficos de las especies involucradas (19 exclusivas del área de estudio), las que pueden reunirse en siete áreas escorpiológicas: Desierto costero, Lomas, Serranía esteparia, Puna, Queswa, Valles interandinos cálidos y Yungas. Las Lomas, Serranía esteparia y Queswa, presentan la mayor riqueza específica. Se analizan los patrones de distribución de las especies, que sugieren la presencia de tres componentes faunísticos en el sur del Perú: andino patagónico, amazónico y caribeño.
Palabras clave: Escorpiones, Sudamérica, Andes, sur del Perú, patrones de distribución.
Abstract
The geographical distribution of 24 scorpion species (five genera and three families) from the Andean corridor in southern Peru is provided. Some zoogeographic details of the implicated species are discussed (19 are exclusive from the area); these species are grouped into seven scorpiological areas: Desierto costero, Serranía esteparia, Puna, Queswa, Valles interandinos cálidos, and Yungas. The Lomas, Serranía esteparia and Queswa areas, are the areas with the most richness of species. The distribution patterns of species, which suggests the presence of three faunistic components in southern Peru: andean-patagonian, amazonian and caribbean, are analysed.
Keywords: Scorpiones, South America, Andes, southern Peru, distribution patterns.
Introducción
El sur peruano al igual que todo el territorio del Perú, presenta una topografía muy accidentada debido fundamentalmente a la presencia de la Cordillera de los Andes, que atraviesa el país de sur a norte siguiendo una dirección paralela al litoral Pacífico. La Cordillera divide al país en tres macro-regiones: una zona desértica ubicada en el lado occidental, la región andina formada por numerosos valles, quebradas, cañones, punas etc. y la zona oriental formada por bosques húmedos que descienden a la Amazonía. La variedad ambiental en el sur del Perú es muy notable, ya que permite observar en desplazamientos de apenas unos cuantos kilómetros, variaciones significativas en la vegetación. Esta diversidad ambiental permite suponer la existencia de diferentes patrones en la diversidad animal del país.
Los estudios zoogeográficos en el Perú son muy escasos, y básicamente los autores han hecho referencia a esquemas y mapas zoo y biogeográficos hechos para Sudamérica o América Latina que naturalmente involucran territorio peruano. Entre las obras más citadas se encuentran las de Rapoport (1968), Fittkau (1969), Cabrera y Willink (1973); pero estas clasificaciones dividen al país de manera muy general y con límites muy imprecisos en sus divisiones (por lo menos en lo que corresponde al Perú); lo mismo ocurre incluso en propuestas más recientes como las de Morrone (1999, 2001, 2002). Todos coinciden en separar al territorio peruano en dos partes: una Andina con componentes andino-patagónicos y otra Amazónica o Guayano-Brasileña que involucraría componentes tropicales y subtropicales. Este hecho fue advertido ya por Sclater (1858) y Wallace (1876) en el siglo XIX, y en trabajos realizados en el Perú por Weberbauer (1945) y Pulgar Vidal (1981) entre otros.
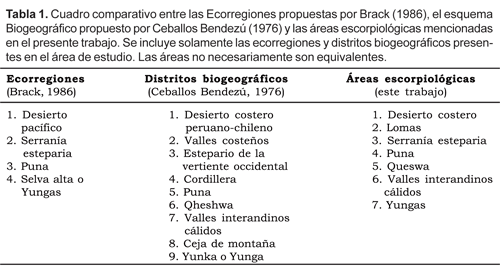
De las propuestas biogeográficas realizadas para el Perú, se ha tomado como base referencial para el presente estudio los trabajos de Ceballos Bendezú (1976) y de Brack (1986). A pesar de ser bastante coherente, el esquema de Ceballos Bendezú (Tabla 1), carece lamentablemente de un mapa, por lo cual resulta complicado plasmar una delimitación de las áreas. La obra de Brack incluye un mapa, pero en él ignora áreas muy importantes constituidas por los valles interandinos (Tabla 1), los cuales se ubican en las ecorregiones de Puna o dentro de las Yungas.
A pesar de que varios autores han considerado a los escorpiones como buenos indicadores biogeográficos -debido fundamentalmente a su limitada capacidad para desplazarse a grandes distancias y a su aparente fidelidad a ciertos ambientes (Mello-Leitão, 1945; Ringuelet, 1953; Maury, 1979; Lourenço, 1986, 1994, 2001; Acosta, 1989; Acosta y Maury, 1998)-; en el Perú no existen antecedentes previos, la precariedad del conocimiento taxonómico de estos arácnidos no ha pemitido realizar estudios de esta naturaleza. Sin embargo, en los últimos años se ha venido aclarando paulatinamente el panorama sistemático de escorpiones en la parte andina del sur del Perú (Ochoa, 1996, 2003a, 2003b, 2004; Ochoa y Acosta 2002a, 2002b, 2003; Acosta y Ochoa, 2000, 2001) lo cual permite dar una primera visión general.
El objetivo del presente estudio es ofrecer un panorama general de los patrones de distribución de los escorpiones en el sur andino peruano y su vinculación con patrones biogeográficos de Sudamérica, así como presentar una clasificación de áreas escorpilógicas preliminares para la zona de estudio. No obstante, al no existir una información filogenética apropiada en todos los taxa involucrados, además de la falta de datos adecuados en áreas adyacentes (norte de Bolivia, centro del Perú y Amazonía peruana), se limitan enormemente las conclusiones aquí presentadas, las cuales deben ser consideradas como muy preliminares.
Área de estudio
El área de estudio para el presente trabajo corresponde a la parte andina del sur del Perú, cuyos puntos extremos se localizan: en el norte en el departamento del Cusco en el límite de la selva alta con la llanura amazónica aproximadamente debajo del paralelo 12º S. En el sur, en el departamento de Tacna (frontera con Chile), el punto denominado «La Concordia» a orillas del Pacífico (18º 21' S). Al este en el departamento de Puno, frontera con Bolivia. Al oeste con el litoral Pacífico en el departamento de Arequipa y una línea entre la costa y el norte del departamento del Cusco a lo largo del límite departamental de Ayacucho (fig. 1).
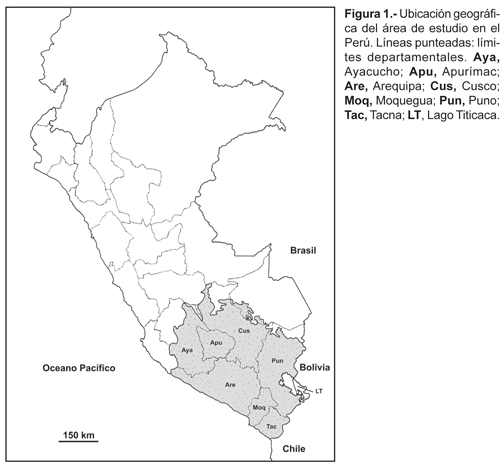
El área incluye la totalidad de los departamentos de Arequipa, Moquegua, Tacna, Apurímac y parte de Ayacucho, Cusco y Puno. Comprende altitudes desde nivel del mar en la costa Pacífica, pasando por las altas cumbres de la Cordillera de los Andes, valles y quebradas interandinas, hasta el lado oriental de los Andes; allí se tomó como referencia aproximadamente los 800 m de altitud, que sería el límite de las Yungas con la llanura amazónica.
Materiales y métodos
El presente estudio está basado en muestreos realizados por el autor en diferentes localidades de la parte andina y desierto del sur del Perú. Para la elección de lugares de colecta se tomaron como referencia las Ecorregiones del Perú propuestas por Brack (1986) y los distritos biogeográficos de Ceballos Bendezú (1976). Se realizaron viajes a más de 30 zonas, incluyendo todas las localidades mencionadas previamente en la literatura, para comprobar la presencia de las especies allí citadas. También se consideraron regiones en la que no existía ningún tipo de reporte y áreas de probable hallazgo de especies de interés.
Las colectas se realizaron mediante el uso de una lámpara UV portátil de empleo nocturno, debido a que los escorpiones permiten su detección por la fluorescencia del tegumento (Stahnke, 1972). Sin embargo este método sólo fue de utilidad en las zonas de desierto y en algunos lugares de las vertientes occidentales. De mayor eficiencia especialmente en zonas andinas, punas y la selva alta, resultó el muestreo durante el día, que consiste en el volteo de piedras. Muchos ejemplares también fueron hallados en cercos de las «chacras» (campos de cultivo). En muy pocas ocasiones se encontraron escorpiones dentro de viviendas humanas; ej. Machupicchu (Cusco), Cañón del Colca (Arequipa), entre otros. En lugares de desierto fue preciso, a veces, excavar el suelo (principalmente arenoso): se ubicaron pequeños hoyos y se procedió a escarbar el terreno sin perder el sentido de la galería; esta técnica no es muy efectiva porque gran parte de los agujeros se encontraban vacíos o con presencia de arañas y lagartijas, pero fue la única forma de encontrar ejemplares en lugares donde no hay piedras y el muestreo con lámpara UV no fue eficaz. Algunos ejemplares fueron capturados también mediante el uso de trampas de caída.
Adicionalmente al material colectado se estudiaron materiales de diversas instituciones: AMNH: American Museum of Natural History, New York, USA. CDA: Cátedra de Diversidad Animal I, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Argentina IML: Instituto Miguel Lillo, Tucumán, Argentina. FMNH: Field Museum of Natural History, Chicago, USA. IBSP-Sc: Instituto Butantan, São Paulo, Brasil. MACN-Ar: Museo Argentino de Ciencias Naturales «Bernardino Rivadavia», Buenos Aires, Argentina. MHNC: Museo de Historia Natural, Facultad de Ciencias Biológicas, Universidad Nacional de San Antonio Abad del Cusco, Perú. MNHN: Muséum National d'Histoire Naturelle, Paris, France. MNRJ: Museu Nacional do Rio de Janeiro, Rio de Janeiro, Brasil. MUSM: Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Perú. MZUC: Museo Zoológico, Universidad de Concepción, Concepción, Chile. MUSA: Museo de Historia Natural, Universidad Nacional de San Agustín de Arequipa, Perú. USNM: National Museum of Natural History, Smithsonian Institution, Washington, USA.
Para la detección y descripción de patrones generales de distribución, se ha seguido la metodología mencionada por Maury (1979): (1) se han elaborado mapas de distribución de cada entidad estudiada, con la ubicación de localidades y demarcación de límites de dispersión de cada taxón presente en el área; (2) se analizaron las geonemias y su posible correlación con factores ambientales (altitud, tipo de vegetación, etc); (3) se determinaron los patrones generales de distribución de especies y géneros y (4) se realizaron análisis de las vinculaciones biogeográficas de los taxa involucrados y su comparación con geonemias conocidas de áreas relacionadas en Sudamérica. Diversidad de escorpiones
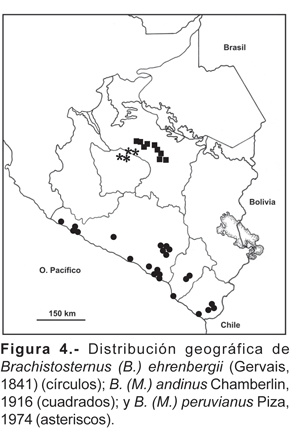
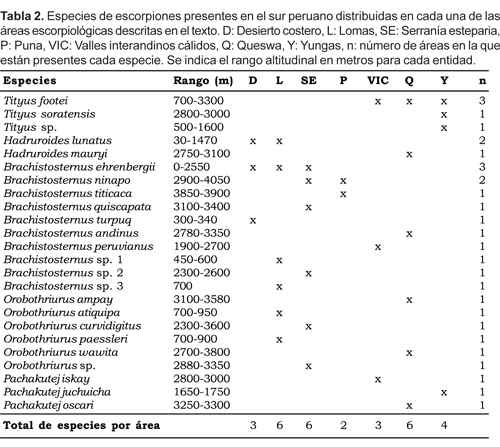
Se encontraron 24 especies y cinco géneros de tres familias de escorpiones en la parte andina del sur del Perú. La familia Bothriuridae aporta la mayor cantidad de elementos andinos (79,2%), siendo el género Brachistosternus el más representativo, con 10 especies (42%); las familias Buthidae y Caraboctonidae solamente aportan el 12,5 y 8,3% respectivamente. Son exclusivas del sur del Perú 19 especies, lo cual demuestra un alto grado de endemismo; las otras cinco especies: B. (B.) ehrenbergii, B. (L.) quiscapata, O. wawita, H. lunatus, y T. soratensis están presentes en otras áreas aledañas.
Familia Buthidae
1. Tityus footei Chamberlin, 1916.- Valles interandinos en los departamentos de Ayacucho, Apurímac y Cusco (fig. 2). Es una de las especies con mayor distribución en el sur del Perú desde los 700-800 m (Cirialo y Sahuayaco) en el límite inferior de la selva alta, es muy abundante en lugares como Santa Teresa, Machupicchu, Alfamayo, lo que correspondería a selva alta y ceja de montaña, donde sube hasta los 3000 a 3100 m. En el fondo de los valles, la distribución de esta especie llega a lugares como Limatambo, Challabamba (Paucartambo), todo el Valle Sagrado de los Inkas, Abancay, Huatatas (Ayacucho), hasta los 3300 m de altitud. También está presente en los cañones de los ríos Apurímac y Pachachaca en ambientes xerofíticos a alturas de 2000 m aproximadamente.
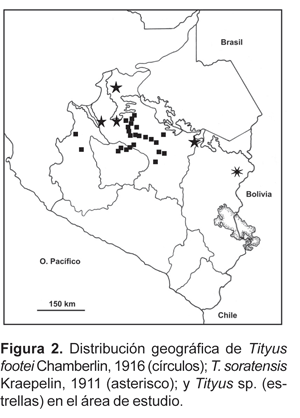
2. Tityus soratensis Kraepelin, 1911.- En el sur del Perú se distribuye solamente en la zona de Sandia en el departamento de Puno (fig. 2); el área corresponde a ambientes de selva alta. Se encuentra también en Bolivia.
3. Tityus sp. [aff. Tityus ecuadorensis].- Se ha registrado en la zona de Kiteni (500 m, La Convención) y en Iskaybamba (1600 m, Marcapata), también en la zona de Kimbiri y Camisea (fig. 2).
Familia Caraboctonidae
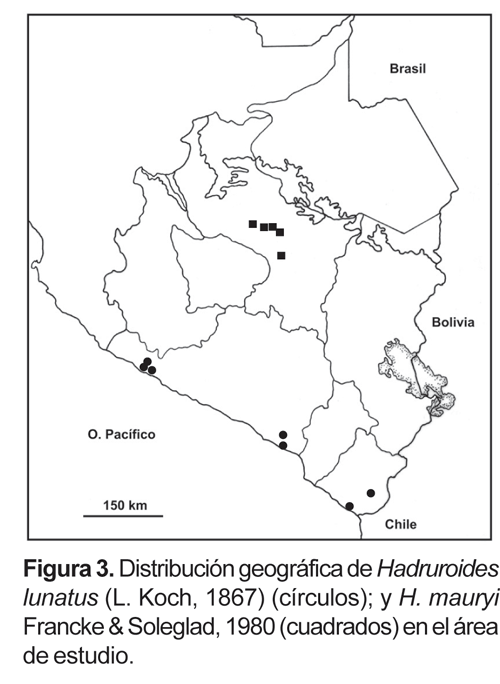
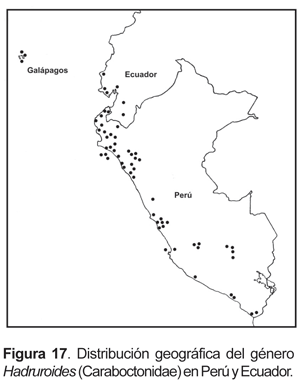
4. Hadruroides lunatus (L. Koch, 1867).- De amplia distribución en el desierto costero; está presente también en algunas islas del litoral Pacífico: isla Lobos de Tierra, isla Chincha. En el sur del Perú se tiene registros en: lomas de Atiquipa, lomas de Mejía, valle de Tambo y en el departamento de Tacna en la zona de Miculla y en quebrada de Burros (fig. 3).
5.- Hadruroides mauryi Francke & Soleglad, 1980.- Valles interandinos en el departamento del Cusco entre los 2750 a 3100 m (fig. 3), en localidades como Chillca, Ollantaytambo, Urubamba, Calca, Yucay y Paruro. Las referencias mencionadas para esta especie de Ayacucho y Huancavelica (Francke y Soleglad, 1980) probablemente correspondan a otra entidad (Ochoa, manuscrito en prep.)
Familia Bothriuridae
6. Brachistosternus (B.) ehrenbergii (Gervais, 1841).- Perú y norte de Chile (a lo largo del desierto costanero del Pacífico). En el sur del Perú, en los departamentos de Arequipa, Moquegua y Tacna desde el nivel del mar hasta los 2550 m, en la vertiente occidental de los Andes (fig. 4).
7. Brachistosternus (L.) ninapo Ochoa, 2005.- Serranía esteparia de los departamentos de Arequipa y Tacna entre los 2900 4050 m (fig. 6). En Arequipa desde Chiguata hasta Zimbral (cercanías del volcán Misti), y en Tacna en las laderas del volcán Yucamane.
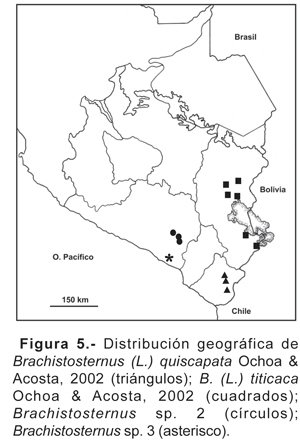
8. Brachistosternus (L.) titicaca Ochoa & Acosta, 2002.- Alrededores del lago Titicaca en el departamento de Puno, entre 3850 a 3900 m (fig. 5). Hasta el momento es la única especie de escorpión reportada para la meseta del Collao. Esta especie ocurre también en la parte boliviana que rodea al lago Titicaca (Ojanguren Affilastro, 2003).
9. Brachistosternus (L.) quiscapata Ochoa & Acosta, 2002.- Vertiente occidental de los Andes de 3100 a 3400 m en el departamento de Tacna y norte de Chile (Provincia Arica) (fig. 5). La zona corresponde a la parte alta de la Ecorregión de Serranía esteparia (Brack, 1986). B. quiscapata no fue encontrado por encima de los 3400 m y probablemente la presencia de la Puna marque el límite de su distribución.
10. Brachistosternus (L.) turpuq Ochoa, 2003.- Por el momento sólo se ha encontrado esta especie en los alrededores de la localidad de Mejía (340 metros de altitud) y en la zona de Cocachacra en el valle del río Tambo. Ambas localidades en el departamento de Arequipa (fig. 6).

11. Brachistosternus (M.) andinus Chamberlin, 1916.- Se distribuye en valles y quebradas interandinas del departamento del Cusco entre los 2780 a 3350 m (fig. 4). Todas las localidades corresponden a la cuenca del río Vilcanota principalmente en el Valle Sagrado de los Inkas y corresponde a ambientes de queswa.
12. Brachistosternus (M.) peruvianus Piza, 1974.- Valles interandinos y cañones profundos en la cuenca de los ríos Apurímac y Pachachaca, en los departamentos de Cusco y Apurímac, desde los 1900 hasta los 2700 m (fig. 4).
13. Brachistosternus sp. 1.- Corresponde a una especie innominada que fue colectada en la quebrada de Burros entre los 450 a 600 m (lomas de Sama) en el departamento de Tacna (fig. 6). El área corresponde a ambientes de lomas.
14. Brachistosternus sp. 2.- Esta especie innominada fue encontrada en los alrededores de la ciudad de Arequipa entre los 2300 a 2600 m (fig. 5), se halla en ambientes xerofíticos en lo que correspondería a la serranía esteparia de la vertiente occidental de los Andes.
15. Brachistosternus sp. 3.- Especie innominada hallada solamente en las lomas de Mejía en el departamento de Arequipa (fig. 5).
16. Orobothriurus ampay Ochoa & Acosta, 2003.- De acuerdo a los datos disponibles, esta especie se encuentra solamente en el bosque de Podocarpus glomeratus Don, del Santuario Nacional del Ampay (departamento Apurímac) entre 3100 a 3580 m (fig. 7).
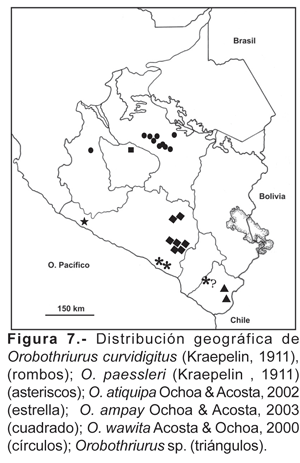
17. Orobothriurus atiquipa Ochoa & Acosta, 2002.- Especie encontrada solamente en las lomas de Atiquipa (entre los 700 a 950 m), ubicada en el norte del departamento de Arequipa (fig. 7). Por el momento no tenemos mayores datos para afirmar que sea una especie endémica para el área de captura, pero el aislamiento de esta formación vegetal y la presencia de otros endemismos en el área lo sugieren.
18. Orobothriurus curvidigitus (Kraepelin, 1911).- Vertientes occidentales de los Andes desde 2300 a 3600 m en el departamento de Arequipa, en lo que correspondería a la Ecorregión de la Serranía esteparia (Brack, 1986), en localidades cercanas a la ciudad de Arequipa, así como también en el cañón del Colca (fig. 7).
19. Orobothriurus paessleri (Kraepelin, 1911).- Departamento Arequipa, lomas de Yuta (Matarani) y lomas de Mejía (fig. 7). La cita para Toquepala en el departamento de Tacna (Maury, 1976) no ha podido ser confirmada. Hasta el momento esta especie sólo se ha encontrado en formaciones de lomas y probablemente está relacionada con este tipo de formación vegetal.
20. Orobothriurus wawita Acosta & Ochoa, 2000.- Valles interandinos de Ayacucho y Cusco, desde 2700 a 3800 m (fig. 7). La distribución correspondería a la zona conocida como queswa (en el sentido de Ceballos Bendezú, 1970, 1976) o Provincia Serrana (Mesoandino + Subpuna; Marín Moreno, 1961). La mayor parte de los ejemplares de O. wawita corresponden a alturas entre 3500 y 3800 m; posiblemente la presencia de la Puna marca en estos lugares el límite de su dispersión. Esta especie fue reportada también para la localidad de Potosí en Bolivia (Acosta y Ochoa, 2002).
21. Orobothriurus sp.- Departamento de Tacna en las localidades de Tarata y Palca entre los 2880 a 3350 m de altitud (fig. 7). El área corresponde a la serranía esteparia. Esta especie correponde a una especie innominada.
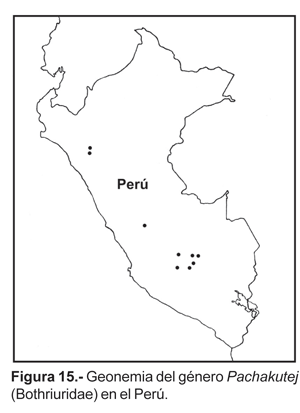
22. Pachakutej iskay (Acosta & Ochoa, 2001).- Valles interandinos entre los 2800-3000 m en los departamentos de Apurímac y Cusco (fig. 6). Colectada en las localidades de Carrizales (Limatambo, 2800 m), Tamburco (Abancay) a 2900 m y Andahuaylas (3000 m).
23. Pachakutej juchuicha Ochoa, 2004.- Conocida solamente para la localidad de Llactawaman (1650-1750 m), en la cordillera de Vilcabamba, en la cuenca del río Apurímac (Distrito de Kimbiri, departamento Cusco; fig. 6). El área corresponde a un bosque húmedo o de selva alta.
24. Pachakutej oscari Ochoa, 2004. Hasta el momento se ha colectado sólo en la localidad de Pallata cerca de Ollantaytambo (departamento del Cusco) entre 3250-3300 m (fig. 6), la localidad está ubicada en una subcuenca del Valle Sagrado de los Inkas (río Patacancha, afluente del río Vilcanota).
Áreas escorpiológicas
De acuerdo con la información hasta ahora obtenida, todas las especies pueden ser reunidas en las siguientes áreas escorpiológicas (fig. 8): Desierto costero, Lomas, Serranía esteparia, Puna, Queswa, Valles interandinos cálidos y Selva alta o Yungas.
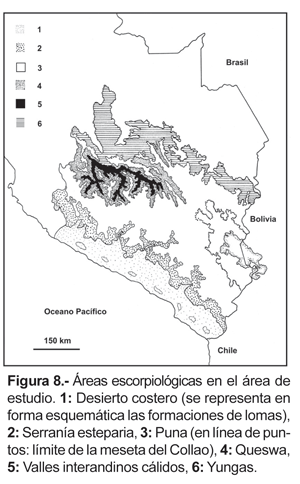
No obstante, los límites de las áreas presentados en el mapa de la figura 8, son en cierto modo convencionales, más aún tratándose del sur del Perú, donde el panorama resulta muy complicado debido a la topografía; ésta no permite que sea una tarea fácil delimitar áreas con precisión, además por la fisiografía muy accidentada que en recorridos relativamente de unos cuantos kilómetros se puede pasar por más de un ambiente.
1. Desierto costero. Corresponde a la franja desértica y semidesértica que se extiende a lo largo del litoral Pacífico, desde el nivel del mar hasta los 2000-2500 m aproximadamente (fig. 8). Las precipitaciones son prácticamente ausentes; casi siempre menores a 100 mm anuales, incluso en algunos lugares como las pampas de Majes y Vítor en Arequipa la precipitación no supera los 10 mm (Peñaherrera del Águila, 1986). La vegetación es casi nula, hay líquenes y plantas esporádicas o adaptadas a las condiciones desérticas y de aridez como las cactáceas y algunas bromeliáceas; sin embargo este panorama se ve alterado por la presencia de los llamados «oasis fluviales» que bordean los ríos y riachuelos que van hacia el Pacífico. El límite superior del desierto ha sido considerado aproximadamente a los 1000 m (Brack, 1986), pero ello ocurre ciertamente en el norte y centro del Perú. En el sur del Perú las condiciones de extrema aridez se extienden aproximadamente hasta los 2000 m y su influencia puede llegar incluso a mayor altura (Marín Moreno, 1961; Pearson, 1982). Prueba de ello es la distribución de B. (B.) ehrenbergii que se encuentra desde el nivel del mar hasta los 2550 m, siendo la especie que más caracteriza a las zonas arenosas del desierto. La zona comprendida entre los 2000-2500 m seguramente corresponde a un área de ecotonía entre el desierto y la serranía esteparia. H. lunatus también presenta una distribución amplia en el desierto costero pero se ubica en otro microhábitat (zonas pedregosas); al parecer es más abundante en ambientes de lomas y contrariamente a la especie anterior, no se encuentra en zonas arenosas (Dávila Flores, 1982). Otra especie presente en el área pero con distribución más restringida es B. (L.) turpuq (fig. 6) que fue colectada solamente en suelos compactos de color rojizo en el sector de Mejía(Ochoa, 2003b).
2. Lomas.- Dentro de esta área se incluye una formación vegetal muy característica y única de la región costanera; corresponde a una vegetación efímera y periódica denominada comúnmente con el nombre de «lomas», las que se extienden desde Trujillo en el norte de Perú (8 ºS) hasta Coquimbo en Chile (30 ºS) (Ferreyra, 1986). Las lomas son parches de vegetación aislados completamente por desierto (Péfaur, 1981) que se desarrollan en las laderas inclinadas al oeste, cercanas al litoral Pacífico; se forman en época invernal (junio a octubre) entre los 700 a 1000 m, producto de la condensación de la alta nubosidad de la temporada, en época de verano se hallan relativamente libres de niebla (Bowman, 1938). La vegetación predominante corresponde a comunidades herbáceas de crucíferas, malváceas, umbelíferas, solanáceas y nolanáceas con varios géneros y especies endémicas (Marín Moreno, 1961). En el sur del Perú se han encontrado 6 especies de escorpiones en estos ambientes, dos de ellas (B. ehrenbergii y H. lunatus) son especies que se hallan también en el desierto propiamente dicho. Otras cuatro especies son propias de las lomas: dos especies del género Orobothriurus: O. atiquipa, O. paessleri, y dos especies innominadas del género Brachistosternus. A pesar de constituir áreas geográficas relativamente pequeñas, presentan un significativo número de especies de escorpiones.
3. Serranía esteparia. Comprende la franja de vegetación arbustiva y xerofítica ubicada en las vertientes occidentales de los Andes en los departamentos de Arequipa, Moquegua y Tacna, por encima de los 2300-2500 m aproximadamente, hasta 3800 m en el límite con la Puna; en esta área se incluyen los cañones profundos del Colca y de Cotawasi. Cinco especies de escorpiones han sido colectadas en estos ambientes: O. curvidigitus, Orobotriurus sp., B. (L.) ninapo, B. (L.) quiscapata y Brachistosternus sp. 2. La primera se distribuye de 2300-3600 m, pero es más abundante por encima de los 2800 m; en el límite inferior de su distribución, O. curvidigitus está presente solamente en el monte ribereño. B. (L.) ninapo se encuentra desde 2900-4050 m llegando incluso a introducirse en la Puna. B. (L.) quiscapata y Orobothriurus sp. presentan una distribución más restringida (2880-3400 m). Brachistosternus sp. 2. sólo fue encontrado entre los 2300 a 2600 m. Una sexta especie [B. (B.) ehrenbergii], puede ser hallada en la parte baja de la serranía esteparia, donde probablemente corresponda a un área de ecotonía con el desierto costero.
4. Puna. Abarca las altas mesetas, áreas dominadas principalmente por pastizales (pajonal), y algunos bosques altoandinos encima de los 3800 m. El único escorpión hasta la fecha capturado para esta zona es B. (L.) titicaca, presente en zonas del altiplano puneño en localidades cercanas al lago Titicaca (fig. 5). En las vertientes occidentales en el límite con la Puna se halla B. (L.) ninapo: Zimbral (4050 m) y Cardarave (3860 m); en dichos sectores se efectuó muestreos a mayor altitud, pero no hubo capturas. Al parecer esto indicaría un avance de B. (L.) ninapo a lugares de Puna, ya que es más abundante en las partes más bajas. Hasta la fecha no existen registros de escorpiones en otras regiones de Puna, incluyendo gran parte del Altiplano. Se ha realizado viajes de colección a diferentes lugares de Puna especialmente en la cordillera Oriental y provincias altas del departamento del Cusco; pero no se ha encontrado escorpiones. La cercanía de la selva alta y ceja de montaña con la Puna de la cordillera Oriental ubicada en los departamentos de Cusco, Apurímac y Ayacucho, hace que sea más húmeda en comparación con el altiplano puneño o la Puna presente en el lado occidental; probablemente esta alta humedad, constituya ambientes negativos para los escorpiones. Sin embargo, la presencia de escorpiones en la Puna de la cadena occidental queda sujeta a confirmación ya que en el centro del Perú fueron halladas otras especies de Orobothriurus por encima de los 4500 m (Maury, 1976; Ochoa, obs. pers.).
5. Queswa. Comprende las quebradas y valles interandinos presentes en los departamentos de Cusco, Apurímac y Ayacucho entre los 2600-3800 m (fig. 8). De clima templado y con una vegetación arbustiva y subarbustiva-espinosa (Marín Moreno, 1961), la queswa se ubica en el piso inmediato inferior a la Puna y es una zona que ya fue reconocida por varios autores como unidad biogeográfica. Corresponde a la «Zona Templada» de Chapman (1921), «Distrito Central» o del «Vilcanota» (Chávez Chaparro, 1947), «Qhechua» y «Suni» (Pulgar Vidal, 1981), «Provincia Serrana» (Marín Moreno, 1961) o «Distrito de Qheshwa» (Ceballos Bendezú, 1970, 1976). Sin embargo muchos autores, especialmente en esquemas generales, han ignorado esta área, ubicándola dentro de la Puna (Cabrera y Willink, 1973; Brack, 1986; Morrone, 1999, 2001, 2002; entre otros) debido principalmente al desconocimiento de sus características climáticas y de vegetación que son diferentes a la Puna. En la queswa se encuentran como especies exclusivas: Brachistosternus (M.) andinus [ 2780-3350 m], O. wawita [ 2700-3800 m], P. oscari [ 3250-3300 m] y H. mauryi [ 2600-3100 m] cuyas distribuciones coinciden apreciablemente con el sistema de valles interandinos (ver fig. 9). Otra especie que se halla en estos ambientes, aunque no exclusivo, es el Buthidae T. footei que se encuentra también en la selva alta y en los valles interandinos cálidos. O. ampay [ 3100-3580 m], que se distribuye exclusivamente en el bosque de Podocarpus glomeratus del Ampay, puede ser considerado como perteneciente a la queswa.
6. Valles interandinos cálidos. Incluye los cañones profundos de clima cálido seco, formados por los ríos Apurímac, Pachachaca y Pampas en los departamentos de Cusco, Apurímac y Ayacucho, entre los 1800-2800 m aproximadamente (fig. 8). La vegetación es xerofítica, formada por bosque y matorral pluvifolio acompañada de plantas espinosas (Weberbauer, 1945; Marín Moreno, 1961). Corresponde a la Provincia de «Montes Pluvifolios» de Marín Moreno (1961) y a la unidad biogeográfica «Pampas» (Lamas, 1982). En esta área están presentes B. (M.) peruvianus, P. iskay y T. footei; las dos primeras probablemente sean especies relacionadas exclusivamente con este ambiente, mientras que T. footei -como ya se mencionó- se encuentra también en otras áreas.
7. Selva alta o Yungas. Corresponde a los densos bosques nublados de clima muy húmedo, ubicados en las vertientes orientales de los Andes; abarcando las partes altas de las cuencas de los ríos Tambopata, Inambari, Kosñipata, Urubamba y Apurímac, comprende la parte oriental de las cordilleras de Carabaya, Vilcanota y Vilcabamba. Se halla desde los 500-600 m hasta los 3500-3800 m (fig. 8). Algunos autores han reconocido varios pisos de vegetación (Weberbauer, 1945; Marín Moreno, 1961; Terborgh, 1971). A nivel faunístico, en las partes altas las yungas se encuentran ligadas a elementos andinos y a medida que se desciende en altitud la influencia amazónica se hace más evidente. En cuanto a escorpiones se refiere, esta área es dominada por elementos de Buthidae; las tres especies de Tityus están presentes en este sector: T. footei, T. soratensis, Tityus sp. [aff. T. ecuadorensis]. Solamente un elemento de Bothriuridae (P. juchuicha) se encuentra en el área. Las dos últimas especies de Tityus y P. juchuicha aparentemente serían de carácter exclusivo, aunque solamente existen 1 ó 2 lotes en cada caso. Por el contrario se tiene buena documentación para T. footei, la cual presenta una amplia distribución desde 700 a 3100 m, siendo muy abundante entre los 1500 y 2500 m. El hecho de que sea la única especie de Tityus presente en ambientes netamente andinos como la queswa y los valles interandinos cálidos, hace suponer que se trata de un avance ulterior desde tierras bajas, aprovechando el fondo de los valles donde las yungas entran en contacto con ambientes andinos a 1800 m (valles interandinos cálidos) y 2500 m (queswa). La ausencia de esta especie por encima de los 3100 m en las laderas orientales y además de la presencia de la Puna son datos a favor de esta suposición. P. juchuicha por el momento sólo se ha encontrado en la cordillera de Vilcabamba en la cuenca del río Apurímac, entre 1650-1750 m, aunque por las condiciones de aislamiento de dicho sistema montañoso es muy posible que sea una especie endémica para el lugar. Es de destacar la ausencia en las yungas de otros géneros como Brachistosternus, Orobothriurus y Hadruroides.
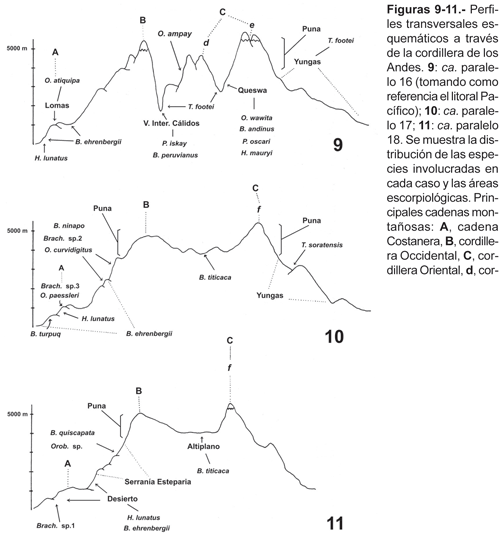
Distribución en perfiles transversales de los Andes
Para una mejor visualización y de manera esquemática se ha representado tres secciones transversales a través de la Cordillera de los Andes (figs. 9-11). Una de ellas tomando como punto de referencia en el litoral el paralelo 16º, otra cerca del paralelo 17º cortando aproximadamente a la altura de la ciudad de Arequipa, y la tercera cerca del paralelo 18º cortando los departamentos de Tacna y Puno. El primer perfil transversal (fig. 9) muestra una mayor complejidad geográfica con respecto a los otros dos perfiles (figs. 10 y 11). Es precisamente a dicho nivel donde se desarrollan peculiares formaciones vegetales como la queswa y los valles interandinos cálidos. Por el contrario, la fisiografía de los dos perfiles ubicados más al sur está dominada por la presencia de la meseta del Collao (cuenca del lago Titicaca).
Con excepción de H. lunatus y B. ehrenbergii que están presentes a lo largo de todo el desierto, los perfiles transversales muestran un cambio sustancial y reemplazo de la fauna de norte a sur; al mismo tiempo muestran claramente una disminución en el número de especies; esto último probablemente debido a que el Altiplano, que ocupa una buena proporción del área de estudio (fig. 8), solamente alberga una especie (B. titicaca). Por otro lado, es de destacar que ningún escorpión ubicado en las vertientes occidentales se halla en los valles interandinos al otro lado de la cordillera Occidental y lo mismo ocurre inversamente. La mayoría de las especies (20 spp.) están distribuidas (cada una de ellas) en una sola área escorpiológica; H. lunatus y B. ninapo en dos áreas, B. ehrenbergii en tres áreas, aunque su presencia en la serranía esteparia es periférica. Solamente T. footei presenta gran adaptación a vivir en áreas tan diferentes como las yungas, la queswa y los valles interandinos cálidos (Tabla 2). Las áreas con mayor diversidad específica son la queswa, lomas y serranía esteparia (6 spp cada una, Tabla 2); los dos primeros casos llaman la atención, porque ocupan una extensión geográfica relativamente menor comparado por ejemplo con las yungas, la puna y el desierto (fig. 8). Las zonas áridas en el sur peruano (desierto, serranía esteparia y valles interandinos cálidos), presentan una diversidad muy pobre, comparado con otros lugares de ambientes similares en otras latitudes (Polis, 1990). Por ejemplo el desierto costero alberga tres especies en el sur del Perú (Tabla 2), incluso en muchas localidades se pudo hallar solamente a B. ehrenbergii. En general en todo el sur del Perú existe un bajo número de especies que viven en simpatría: tres o cuatro especies a lo sumo; solamente en el Valle Sagrado de los Inkas es posible encontrar en espacios relativamente cortos (2 km) hasta cinco especies, pero estrictamente no parecen compartir el mismo microhábitat (Ochoa, obs. pers.). Lo mismo ocurre cerca de la localidad de Mejía en el departamento de Arequipa donde se pueden hallar cinco especies. Estos números resultan relativamente muy bajos al comparar con otras localidades de Sudamérica como Manaus en Brasil (8 especies; Lourenço, 1988) y Chancaní en Argentina (9 especies; Acosta, 1995). El récord a nivel mundial para una localidad corresponde al desierto de Baja California (12 especies; Polis, 1990).
Si bien la diversidad local (alfa) no es alta, a nivel regional (diversidad gamma) resulta importante, dedido al recambio de especies que es significativo; esto último, sumado al alto grado de endemismo de la región, son aspectos de suma importancia que podrían contribuir en la elaboración de estrategias de conservación de la zona, especialmente en áreas pequeñas como la queswa y las lomas que presentan la mayor diversidad de escorpiones.
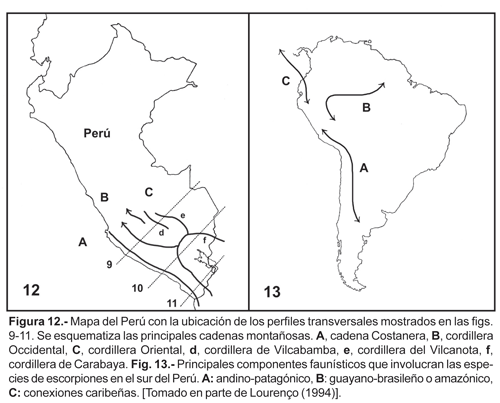
Vinculaciones
Todas las especies involucradas en el área de estudio, están ligadas a un patrón andino de distribución, [corredor escorpiológico Andino de Lourenço (1994)] este corredor discurre a lo largo de la cordillera, desde Colombia hasta la Argentina. Sin embargo la diversidad de la fauna de escorpiones presentes en el área de estudio tienen diferentes orígenes (fig. 13). Las especies de los géneros Brachistosternus, Orobothriurus y Pachakutej (Bothriuridae), están íntimamente relacionadas a la fauna de origen andino-patagónico. El género Tityus probablemente tenga un origen guayano-brasileño (en el sentido de Rapoport, 1968) o Neotropical s.s. (en el sentido de Morrone, 1996, 1999, 2002), mientras que el género Hadruroides al parecer tiene antiguas conexiones con elementos caribeños (Francke y Soleglad, 1981).
Patrón andino. En el área de estudio están presentes los tres subgéneros de Brachistosternus. B. (Brachistosternus), actualmente monotípico, se extiende desde el sur de Ecuador, todo el litoral peruano hasta el norte de Chile (Aguilar y Meneses, 1970; Maury, 1973; Francke, 1977), la geonemia del subgénero se limita solamente a lo largo del desierto y las vertientes occidentales, el cual podría corresponder a un corredor andino secundario («andino occidental», fig. 14a). El subgénero B. (Leptosternus), actualmente incluye 29 especies extendidas a lo largo de los Andes y el desierto, desde el sur peruano hasta la provincia de Santa Cruz en Argentina. Tres especies presentes en el área de estudio (B. ninapo, B. quiscapata y B. titicaca), están relacionadas con un grupo de especies, -habitantes también de alta montaña-, distribuidas en el oeste de Bolivia, norte de Chile y noroeste de Argentina, siguiendo las cadenas montañosas por sobre los 2500 m (Ojanguren Affilastro, 2003; Ochoa, 2005; Ochoa y Acosta, 2002b). Morfológicamente estas especies son muy parecidas, sumado al hecho de habitar grandes alturas, hace pensar que los procesos de especiación corresponden a eventos recientes; sin embargo no se dispone de otros datos que sustenten esta postura. La distribución de este grupo podría corresponder a otro corredor andino secundario (corredor «altoandino», fig. 14b). Por otro lado, B. (L.) turpuq y las tres especies innominadas del subgénero B. (Leptosternus) presentes en las vertientes occidentales (Brachistosternus sp1, sp2, sp3), están relacionadas con especies del norte de Chile que habitan similarmente zonas desérticas, y al mismo tiempo con otras especies distribuidas en el oeste, centro y sur de Argentina, que se encuentran también en zonas áridas (Maury, 1984b; Roig Alsina y Maury, 1984; Ojanguren Affilastro y Roig Alsina, 2001; Ojanguren Affilastro, 2002, com. pers.). La distribución de estas especies supondría una extensión del mismo corredor andino-occidental y además de otro corredor que correspondería al «peripampásico» propuesto por Acosta (1989) (fig. 14c); ambos corredores secundarios tendrían una posible conexión antigua en el centro-sur de Chile, cuando la cordillera Andina presentaba altitudes más bajas y las zonas desérticas de ambos lados estaban conectadas.
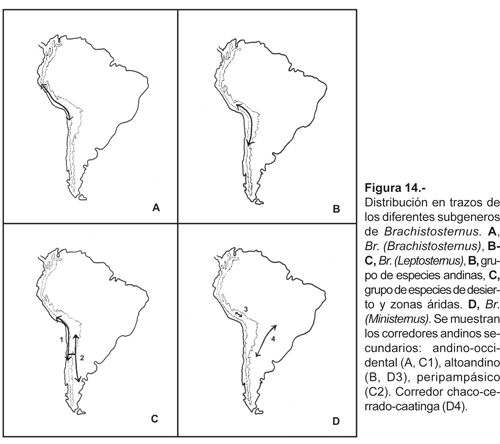
El subgénero B. (Ministernus) presenta una distribución disyunta, las dos especies presentes en el área de estudio, B. (M.) andinus y B. (M.) peruvianus, se hallan en ambientes altoandinos; mientras que las otras dos especies, B. (M.) ferrugineus (Thorell, 1876) y B. (M.) simoneae Lourenço, 2000, están relacionadas con el corredor «Chaco-Cerrado-Catinga» (Lourenço, 1994, 2000) (fig. 14d). La presencia de estas especies en ambientes de Chaco y Cerrado podría ser interpretada como una distribución apocórica en todo el género con base en que las dos especies peruanas del subgénero mantienen una distribución andina, característica de la mayoría de las especies del género (Acosta, 1989; Acosta y Maury, 1998).
El género Orobothriurus, es el taxón que más fielmente se refleja en el corredor Andino; en el área de estudio están presentes cuatro especies en las vertientes occidentales (O. atiquipa, O. curvidigitus, O. paessleri y Orobothriurus sp.; una especie en los valles interandinos (O. wawita) y otra (O. ampay) exclusiva del bosque de Podocarpus del Ampay. La geonemia de todo el género [ 8 especies descritas y varias innominadas, todas de distribución alopátrida (Ochoa, 2004)], abarca desde el centro de Perú (cordillera Blanca en el departamento de Ancash), norte y centro de Chile, oeste de Bolivia y llega al centro-oeste de Argentina, comprendiendo ambientes andinos y subandinos (fig. 16). El género como tal probablemente tenga un origen más antiguo que el levantamiento de la cordillera Andina, lo cual condujo a eventos vicariantes que ocasionaron el aislamiento y especiación de sus integrantes. Varias especies viven en alta montaña por sobre los 4000 m, y una especie innominada, presenta el récord de colecta a mayor altitud del mundo; no obstante no se ha podido registrar este género en la Puna del sur del Perú. Ninguna de las especies de Orobothriurus incursiona en ambientes de yungas y tampoco en los valles interandinos cálidos. Las dos especies que se encuentran en las zonas cercanas al litoral (O. atiquipa y O. paessleri) en realidad están relacionadas a ambientes de lomas, sin estar presente en el desierto propiamente dicho. La presencia de estos Orobothriurus podría ser explicada por la existencia de antiguas conexiones de las lomas con ambientes netamente andinos y que posteriormente dichas formaciones quedaron como parches de vegetación aislados al incrementarse las condiciones de aridez en la costa peruana y produciendo una especiación ulterior en estos lugares. La afinidad florística de las lomas con elementos andinos fue postulada hace muchos años por Herrera (1930), quien encontró que la mayoría de familias y géneros de plantas eran comunes entre las lomas del departamento de Arequipa y los valles interandinos del Cusco. Otros ejemplos de esta afinidad se da también en vertebrados como los roedores (H. Zeballos, com pers.). Un punto a favor sobre esta postura se da en el análisis cladístico de Orobothriurus (Ochoa, 2004), donde la especie más relacionada con O. atiquipa y O. paessleri es O. curvidigitus que precisamente se distribuye en la serranía esteparia en el mismo lado occidental de los Andes por sobre los 2300 m.
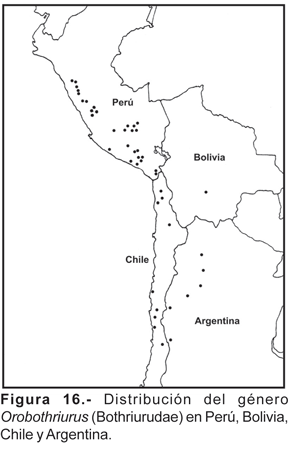
El género Pachakutej es exclusivo del Perú y presenta una distribución en los valles interandinos y lugares de selva alta (fig. 15). Es importante resaltar que a diferencia de otros géneros de Bothriuridae (ej. Bothriurus, Brachistosternus, Orobothriurus) ninguna especie de este género se encuentra en las vertientes occidentales ni en el desierto, ni tampoco llegan a lugares de Puna. Las especies se distribuyen en pisos altitudinales cortos entre los 2800-3300 m, solamente una especie se distribuye a menor altura (1650-1750 m: P. juchuicha). Por el momento Pachakutej presenta una distribución disyunta con respecto a Bothriurus [probablemente el grupo hermano de acuerdo al análisis cladístico realizado para el género (Ochoa, 2004)]. Las especies de distribución más cercanas de Bothriurus corresponden a B. bocki Kraepelin, 1911 y B. inermis Maury, 1981; la primera de ellas está presente en localidades de alta montaña en Bolivia cerca a la frontera peruana, y B. inermis se encuentra desde Santa Cruz en Bolivia y llega a introducirse en la periferia de la selva amazónica en Madre de Dios (Maury, 1981, 1984a; Acosta y Ochoa, 2002; Mattoni, com. pers.). Al parecer el altiplano de la cuenca del Titicaca y la influencia amazónica por el lado oriental pueden constituir barreras para ambos géneros. Sin embargo no se puede dar más que una primera aproximación de la geonemia del género, debido fundamentalmente al escaso número de ejemplares colectados en todas las especies y las pocas citas de localidades.
Patrón neotropical. Tityus footei, T. soratensis y T. ecuadorensis estaban agrupadas junto con otras especies relacionadas en el ex-complejo «Tityus bolivianus» (Lourenço y Maury, 1985). Según los datos que se disponen de este grupo de especies (en el que se puede incluir también a Tityus sp.), se hallan en su mayoría distribuidas a lo largo de las yungas o selva alta, desde Ecuador hasta el noroeste argentino, lo cual refleja una relación marginal con el corredor «Andino» (Lourenço, 1986, 1994). Sin embargo la mayor parte de especies de Tityus presentes en Sudamérica, están ligados a los corredores, «Chaco-Cerrado-Caatinga», « «Amazónico» y «Guayano» de Lourenço, por lo cual es casi seguro que dichas especies presentes en el área tengan un origen guayano-brasileño (fig. 13).
Conexiones caribeñas. La vinculación que presenta el género Hadruroides es un tanto diferente. Si bien es cierto que la mayoría de las especies de este género junto con su grupo hermano Caraboctonus (integrantes de la familia Caraboctonidae) presentan una distribución andina y en el desierto, una de las especies de Hadruroides está presente en las Islas Galápagos (fig. 17), las cuales biogeográficamente están relacionadas con elementos caribeños ya que forman parte del Dominio o Subregión Caribe o Caribeña (Cabrera y Willink, 1973; Morrone, 1999, 2002). El asunto es mucho más complicado en un contexto más amplio, ya que el otro género americano en la familia Caraboctonidae se distribuye en México y el Oeste de los EE.UU. (Hadrurus). Esta distribución fue explicada, por Francke y Soleglad (1981), como el track «SO de Perú-Galápagos-México» de Croizat (1958). De acuerdo a estos autores, durante el Mesozoico e inicios del Terciario en que estuvieron separados Norteamérica y Sudamérica, habría quedado aislado Hadrurus de los elementos sudamericanos. Posteriormente con la separación de las Islas Galápagos durante el Eoceno e inicios del Oligoceno, se produjo el aislamiento y subsecuente especiación de Hadruroides galapagoensis Maury, 1975. Las otras entidades especiaron en el continente y actualmente la mayoría de Hadruroides están presentes en el norte de Perú y sur de Ecuador (Maury, 1975), sólo dos especies llegan al sur de Perú y probablemente correspondan al límite meridional del género (fig. 17). No se tienen datos de que el género se encuentre en actual territorio boliviano, y tal como fue manifestado para el género Pachakutej, el altiplano de la meseta del Titicaca podría haber actuado como una barrera. Por el lado del desierto costero es muy probable que el género se encuentre también en el norte de Chile.
Sin duda que el conocimiento actual que tenemos de aspectos biogeográficos de los escorpiones en el sur del Perú es muy limitado, aquí solamente se ha presentado una visión general, pero existen muchos vacíos de información que necesitan ser llenados para comprender de mejor manera la distribución de estos arácnidos. Hay muchas áreas que requieren ser exploradas más intensivamente no solamente en el sur peruano, sino en todo el país. Es fundamental continuar con estudios de sistemática alfa en los distintos grupos; así como también con trabajos de corte ecológico que son prácticamente inexistentes para la región. Por otro lado se requieren análisis filogenéticos de la mayoría de taxa involucrados, que ayuden a entender e interpretar los eventos históricos.
Agradecimientos
Expreso mi agradecimiento a los Drs. Oscar Ochoa Mendieta, Luis E. Acosta, Beatriz Rosso de Ferradás, Pablo Goloboff, Sergio Roig y Camilo Mattoni, por los consejos y sugerencias para el presente estudio. También a un árbitro anónimo por las correcciones y sugerencias en el manuscrito. La mayor parte de los datos presentados en el presente trabajo corresponden a colecciones personales del autor y expreso mi especial reconocimiento a todas las personas que me ayudaron en las tareas de colecta: Blgos. Horacio Zeballos P., Evaristo López, Elias Ponce M., Roberto Gutiérrez, Ulrich Zanabria, Sirley Rivera, Joaquín Ugarte, Lizette Tejada, Henrry Alayo, Alejandro Pari, José L. Velásquez, Eduardo Berneo, Enrique Oscco, John Achicahuala, Juan Carlos Chaparro, Oscar Mujica, Gorky Valencia, Wilfredo Mendoza, Ronald Aimituma, Daniel Muñiz, Juan Flores, Edilberto Orosco, B. Romero, E. Santi. De la misma manera a los curadores y responsables de las colecciones de los museos e instituciones que facilitaron ejemplares para su estudio: Lorenzo Prendini (AMNH), Luis E. Acosta (CDA), Luis Grosso, Marcela Peralta (IML), Alfred F. Newton (FMNH), Denise Candido (IBSP-Sc), Cristina Scioscia, Andrés Ojanguren Affilastro (MACN-Ar), Olintho Aguilar (MHNC), Wilson Lourenço (MNHN), Adriano Kury (MNRJ), Gerardo Lamas (MUSM), Jorge Artigas (MZUC), Evaristo López, Alberto Morales (UNSA), Jonathan Coddington, Scott Larcher (USNM). Agradezco también a las personas que me dieron apoyo logístico en los viajes de colecta: Ing. Marco Arenas y Blgo. Arturo Cornejo (INRENA-RNLM), Prof. Guido Zúniga (UNAP); Blgos. Percy Jiménez, Luis Villegas y Aldo Ortega (IRECA). Mi reconocimiento a Andrés Ojanguren Affilastro por brindarme información de especies chilenas de Brachistosternus. Este trabajo forma parte de mi Tesis doctoral realizada en la Facultad de Ciencias Exactas, Físicas y Naturales de la Universidad Nacional de Córdoba (Argentina).
Literatura citada
Acosta L.E. 1989. La Fauna de Escorpiones y Opiliones Arachnida) de la provincia de Córdoba. Tesis doctoral, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, i-vi & 333 pp. [ Links ]
Acosta L.E. 1995. The scorpions of the Argentinian Western Chacho II. Community survey in the Llanos District. Biogeographica, 71 (2): 49-59. [ Links ]
Acosta L.E. y E.A. Maury. 1998. Scorpiones. En: Morrone, J.J. y Coscaron, S. (dirs.), Biodiversidad de Arthropodos argentinos. Una perspectiva biotaxonómica. Pp. 545559. Ediciones Sur, La Plata. [ Links ]
Acosta L.E. y J.A. Ochoa. 2000. Nueva especie de Orobothriurus Maury del Perú (Scorpiones, Bothriuridae). Revue Arachnologique, 13 (10): 135-144. [ Links ]
Acosta L.E. y J.A. Ochoa 2001. Two new species of Orobothriurus Maury, 1976 from Argentina and Peru, with comments on the systematics of the genus (Scorpiones: Bothriuridae). In: V. Fet & P.A. Selden (eds.). Scorpions 2001. In memoriam Gary A. Polis. Pp. 203-214. Burnham Beeches, Bucks: British Arachnological Society. [ Links ]
Acosta L.E. y J.A. Ochoa. 2002. Lista de los escorpiones bolivianos (Chelicerata: Scorpiones), con notas sobre su distribución. Revista de la Sociedad Argentina de Entomología, 61 (3-4): 15-23. [ Links ]
Aguilar P.G. y O. Meneses. 1970. Escorpiones y Escorpionismo en el Perú I: Nota preliminar sobre los Scorpionida peruanos. Anales Científicos de la Universidad Nacional Agraria La Molina, 8: 1-5. [ Links ]
Bowman, I. 1938. Los Andes del Sur del Perú. Editorial La Colmena S.A., Arequipa, Perú, 267 pp. [ Links ]
Brack A. 1986. Ecología de un país Complejo. En: Gran Geografía del Perú. Naturaleza y Hombre. Volumen II, Pp. 175-319. ManferJuan Mejía Baca, Barcelona, España. [ Links ]
Cabrera A.L. y A. Willink. 1973. Biogeografía de América Latina. Monografías Científicas OEA, Serie Biología Nº 13, pp I-VI, 1-117. [ Links ]
Ceballos-Bendezú I. 1970. Los pisos zoogeográficos del Departamento del Cusco. Publicaciones del Departamento Académico de Zoología y Entomología, Universidad Nacional de San Antonio Abad del Cusco,(1): 1-26. [ Links ]
Ceballos-Bendezú I. 1976. Nuevo esquema biogeográfico del Perú. Revista Universitaria, Cusco, 130: 19-44. [ Links ]
Chapman F.M. 1921. The distribution of bird-life in the Urubamba valley of Peru. Bulletin U.S. National Museum, 117: 1-138. [ Links ]
Chávez-Chaparro J. 1947. Ensayo zoogeográfico del Departamento del Cusco y descripción de sus principales Mamíferos. Editorial H. G. Rozas S.A., Cusco, Perú, 98 pp. [ Links ]
Croizat L. 1958. Panbiogeography. Vol. 1. The New Word. Publicado por el autor, Caracas, Venezuela, 1018 pp. [ Links ]
Dávila-Flores J. 1982. Escorpiones de las Lomas de Matarani - Islay. En: Salinas, P. (Ed.), Actas del VIII Congreso Latinoamericano de Zoología. Merida, Venezuela, pp. 547-551. [ Links ]
Ferreyra R. 1986. Flora y vegetación del Perú. En: Gran Geografía del Perú. Naturaleza y Hombre. Volumen II, Pp. 1-174. Manfer-Juan Mejía Baca, Barcelona, España. [ Links ]
Fittkau E.J. 1969. The fauna of South America. In: Fittkau, E. J.; Illies, J; Klinge, H.; Schwabe, G.H. y Sioli (Eds.). Biogeography and ecology in South America, 2. Junk, The Hague. Pp. 624-650. [ Links ]
Francke O.F. 1977. Escorpiones y Escorpionismo en el Perú VI: Lista de especies y claves para identificar las familias y los géneros. Revista Peruana de Entomología, 20: 73-76. [ Links ]
Francke O.F. & M.E. Soleglad. 1980. Two new Hadruroides Pocock, from Peru (Scorpiones-Vaejovidae). Occasional Papers of the Museum, Texas Tech University, 69: 1-13. [ Links ]
Francke O.F. & M.E. Soleglad, 1981. The family Iuridae Thorell (Arachnida, Scorpiones). The Jounal of Arachnology, 9: 233-258. [ Links ]
Herrera F.L. 1930. La Vegetación de la Costa Peruana. Revista Universitaria, Cusco, 59 (1): 71-79. [ Links ]
Lamas G. 1982. A preliminary Zoogeographical Division of Peru, Based on Butterfly Distributions (Lepidoptera, Papilionoidea). Pp. 336-357. In: Prance, G. T. (Ed.), The biological model for diversification in the tropics. New York, Columbia University Press. [ Links ]
Lourenço W.R. 1986. Les modèles de distribution géographique de quelques groupes de Scorpions néotropicaux. Compte Rendu des Séances de la Société de Biogéographie, 62 (2): 61-83. [ Links ]
Lourenço W.R. 1988. Synopsis de la fauna scorpionique de la région de Manaus, État d'Amazonas, Brésil, avec déscription de deux nouvelle espèces. Amazoniana, 10 (3): 327-337. [ Links ]
Lourenço W.R. 1994. Biogeographic patterns of tropical South American Scorpions. Studies on Neotropical Fauna and Environment, 29 (4):219-231. [ Links ]
Lourenço W.R. 2000. Confirmation de la présence du genre Brachistosternus Pocock (Scorpiones, Bothriuridae) au Brésil et description d'une espèce nouvelle. Revue Arachnologique, 13 (6): 93-100. [ Links ]
Lourenço W.R. 2001. Scorpion Diversity in Tropical South America. Implication for Conservation Programs. Pp. 406-415. In: P. Brownell & G. Polis (Eds.). Scorpion Biology and Research. Oxford University Press, New York. [ Links ]
Lourenço W.R. y Maury, E. A. 1985. Contribution à la connaissance systemátique des Scorpions appartenant au «complexe» Tityus bolivianus Kraepelin, 1895 (Scorpiones, Buthidae). Revue Arachnologique, 6 (3): 107-126. [ Links ]
Marín-Moreno F. 1961. Panorama fitogeográfico del Perú. Revista Universitaria, Cusco, 120: 9-68. [ Links ]
Maury E.A. 1973. Las tricobotrias y su importancia en la sistemática del género Brachistosternus Pocock, 1894 (Scorpiones, Bothriuridae). Physis, secc. C, 32 (85): 247-254. [ Links ]
Maury E.A. 1975. Escorpiones y escorpionismo en el Perú IV: Revisión del género Hadruroides Pocock, 1893 (Scorpiones, Vejovidae). Revista Peruana de Entomología, 17 (1): 9-21. [ Links ]
Maury E.A. 1976. Escorpiones y escorpionismo en el Perú V. Orobothriurus, un nuevo género de escorpiones altoandinos (Bothriuridae). Revista Peruana de Entomología, 18 (1): 14-25. [ Links ]
Maury E.A. 1979. Apuntes para una zoogeografía de la escorpiofauna Argentina. Acta Zoológica Lilloana, 35 (2): 703-719. [ Links ]
Maury E.A. 1981. A new Bothriurus from Bolivia (Scorpiones, Bothriuridae). Bulletin of the American Museum of Natural History, 170 (1):29-33. [ Links ]
Maury E.A. 1984a. Redescripción de Bothriurus bocki Kraepelin, 1911 (Scorpiones, Bothriuridae). The Journal of Arachnology, 12 (3): 351-356. [ Links ]
Maury E.A. 1984b. Una nueva especie de Brachistosternus de la Argentina (Scorpiones, Bothriuridae). Revista de la Sociedad Entomológica Argentina, 43 (1-4): 113-118. [ Links ]
Mello-Leitão C. 1945. Escorpiões sul-americanos. Arquivos do Museu Nacional, 40: 7-468. [ Links ]
Morrone J.J. 1996. The biogeographical Andean subregion: A proposal exemplified by Arthropod taxa (Arachnida, Crustacea, and Hexapoda). Neotropica, 42 (107-108): 103-114. [ Links ]
Morrone, J. J. 1999. Presentación preliminar de un nuevo esquema biogeográfico de América del Sur. Biogeográfica, 75 (1): 1-16. [ Links ]
Morrone J.J. 2001. Biogeografía de América Latina, y el Caribe. Manuales & Tesis SEA, Sociedad Entomológica Aragonesa, vol. 3, Zaragoza, 148 pp.
Morrone J.J. 2002. Presentación sintética de un nuevo esquema biogeográfico de América Latina y el Caribe. En: C. Costa, A. Vanin J.M. Lobo y A. Melic (Eds.). Proyecto de Red Iberoamericana de Biogeografía y Entomología Sistemática, PRIBES 2002, pp. 267-275.
Ochoa J.A. 1996. Los escorpiones en el departamento del Cusco. Tesis de Biólogo. Facultad de Ciencias Biológicas, Universidad Nacional de San Antonio Abad del Cusco, Perú. 63 pp.
Ochoa J.A. 2003a. Sistemática y patrones de distribución de los escorpiones (Chelicerata) del corredor andino, en el sur del Perú. Tesis doctoral. Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, i-iv, 259 pp.
Ochoa J.A. 2003b. Nueva especie de Brachistosternus Pocock (Scorpiones: Bothriuridae) del Sur del Perú. Revista Peruana de Biología, 9 (2): 55-63.
Ochoa J.A. 2004. Filogenia del género Orobothriurus y descripción de un nuevo género de Bothriuridae (Scorpiones). Revista Ibérica de Aracnología, 9: 43-73.
Ochoa J.A. 2005. Brachistosternus ninapo una nueva especie (Scorpiones:Bothriuridae)de los Andes occidentales en el sur del Perú. Rev peru biol. 11 (2): 139-148.
Ochoa J.A. y Acosta, L. E. 2002a. Orobothriurus atiquipa, a new Bothriurid species (Scorpiones) from Lomas in Southern Peru. The Journal of Arachnology, 30: 98-103.
Ochoa J.A. & L.E. Acosta. 2002b. Two New Andean Species of Brachistosternus Pocock (Scorpiones: Bothriuridae). Euscorpius, Occasional Publications in Scorpiology, 2: 1-13.
Ochoa J.A. y L.E. Acosta. 2003. Una nueva especie de Orobothriurus (Scorpiones: Bothriuridae) del Santuario Nacional Ampay, Apurímac, Perú. Revista Peruana de Entomología, 43: 1-6.
Ojanguren-Affilastro A.A. 2002. Nuevos aportes al conocimiento del género Brachistosternus en Chile, con la descripción de dos nuevas especies (Scorpiones, Bothriuridae). Boletín de la Sociedad Biológica de Concepción, Chile, 73: 37-43.
Ojanguren-Affilastro A.A. 2003. Las especies andinas de Brachistosternus (Leptosternus), con la descripción de tres nuevas especies (Scorpiones, Bothriuridae). Revista Ibérica de Aracnología, 8: 23-36.
Ojanguren-Affilastro A.A. y A. Roig-Alsina. 2001. Brachistosternus angustimanus, una nueva especie del norte de la Patagonia, Argentina (Scorpiones, Bothriuridae). Physis, secc. C, 58 (134-135): 15-22.
Pearson O. 1982. Distribución de pequeños mamíferos en el altiplano y los desiertos de Perú. En: Salinas, P. (Ed.), Actas del VIII Congreso Latinoamericano de Zoología, Mérida, Venezuela. pp. 263-284.
Péfaur J.E. 1981. Composition and phenology of epigeic animal communities in the Lomas of southern Peru. Journal of Arid Environments, 4: 31-42.
Peñaherrera-del-Águila C. 1986. Geografía Física del Perú. En Gran Geografía del Perú. Naturaleza y Hombre. Volumen I, Pp. 1-221. Manfer-Juan Mejía Baca, Barcelona, España.
Polis G.A. 1990. Ecology. In: Polis, G. A. (ed.), The Biology of Scorpions. Pp. 247-293. Stanford University Press, Stanford, California.
Pulgar-Vidal J. 1981. Geografía del Perú. Las Ocho Regiones Naturales del Perú. 8va. Edición. Editorial Universo, Lima, 313 pp.
Rapoport E.H. 1968. Algunos problemas biogeográficos del Nuevo Mundo con especial referencia a la Región Neotropical. En: D. Debouteville y E.H. Rapoport (Eds.). Biologie de l'Amerique Australe, CNRS y CNYCT, Paris, vol. IV, pp. 55-110.
Ringuelet R.A. 1953. Geonemia de los escorpiones en la Argentina y las divisiones zoogeográficas basadas en su distribución. Revista del Museo de La Plata (nueva serie), Zoología, 6 (43): 277-284.
Roig-Alsina A. y E.A. Maury. 1984. Sistemática y distribución geográfica de Brachistosternus (L.) pentheri Mello-Leitão, 1931 (Scorpiones, Bothriuridae). Physis, secc. C, 42 (102): 17-21.
Sclater P.L. 1858. On the general geographic distribution of the members of the class Aves. Zoological Journal of the Linnean Society, 2: 130-145.
Stahnke H. 1972. U.V. light, an useful field tool. BioScience, 22(10):604-607.
Terborgh J. 1971. Distribution on Enviromental gradients: theory and a preliminary interpretation of distributional patterns in the avifauna of the Cordillera de Vilcabamba, Peru. Ecology, 52: 23-40.
Wallace A.R. 1876. The geographical distribution of animals, 2 vols. Reimpreso en 1962. Haefner Publishing Co.,New York.
Weberbauer A. 1945. El Mundo Vegetal de los Andes Peruanos. Estación Experimental Agrícola de La Molina, Lima, 776 pp.
Correspondencia
* Urb. Kennedy «A» Los Brillantes A-19, Wanchaq, Cusco, Perú.
Email:jaochoac2000@yahoo.com ; ochoas@unsaac.edu.pe
