










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.12 n.2 Lima ago./set 2005
Maximilian Weigend1, Eric F. Rodríguez2, César Arana3
1Institut für Biologie Systematische Botanik und Pflanzengeographie, Freie Universität Berlin.
2Herbarium Truxillense (HUT), Universidad Nacional de Trujillo, Trujillo-Perú .
3Museo de Historia Natural y Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos.
Número especial: Bosques relictos del NO de Perú y SO de Ecuador
Weigend, Rodríguez y Arana (Comps.)
Publicado online: 08/12/05
Resumen
Los bosques relictos o fragmentos de bosque en las vertientes andinas Noroccidentales del Perú y Suroccidentales de Ecuador, son frágiles ecosistemas, que en el pasado fue un todo continuo. La importancia y singularidad de los bosques como «refugios» y hábitats con poca dinámica radica en su compleja diversidad florística y endemismos, consecuencia de su complicada topografía y ecología de sus hábitats. En este artículo introductorio se analizan 12 trabajos de investigación expuestos en el Taller sobre Bosques Relictos de la Vertiente Occidental Andina del Norte del Perú y Sur del Ecuador en mayo del 2004 en el X CONABOT, Trujillo-Perú. Aquí, los conocimientos sobre el tema son consolidados y actualizados, es así que los fragmentos de bosque suman más de 20 en el norte del Perú, enfatizando a dos de gran importancia: Bosque La Oscurana (Cajamarca) y Kañaris (Lambayeque). Así mismo, se determinan las relaciones entre los bosques relictos, analiza su dinámica, caracterización y como puede lograrse su conservación inmediata. Se plantean los aspectos cruciales para alcanzar la conservación de estos bosques remanentes: investigar y mostrar urgentemente su gran biodiversidad, subrayar el valor económico potencial de la fitodiversidad que poseen para la región y revalora su papel como áreas de captación de agua para los ríos que son fuente prioritaria para la agricultura y vida urbana en la costa.
Palabras clave: bosques montanos relictos, Andes, vertiente occidental andina, Perú, Ecuador
Abstract
The forest fragments or relict forests on the Western slopes of the Andes in northwest Peru and in south-west Ecuador are fragile ecosystems, which were largely continuous at some stage in the past. The importance and uniqueness of these forests as «refuges» and stable habitats roots partly in their complex diversity and their high levels of endemicity. These in turn are the outcome of a complex topography and ecology. In this introductory chapter 12 studies are analysed, which were presented at the Taller sobre Bosques Relictos de la Vertiente Occidental Andina del Norte del Perú y Sur del Ecuador in May 2004 at the X CONABOT, Trujillo-Perú. This helps to consolidate and update our knowledge on these over 20 forests fragments in northern Peru and two previously unknown fragments are presented for the first time: Bosque La Oscurana (Cajamarca) and Kañaris (Lambayeque). Some ideas are exposed on the the relationships between the relict forests, how to analyse their dynamics, how to characterize their structure and how their conservation may be achieved. It is proposed that crucial elements in a conservation of these forests are a) urgently inventorizing them to demonstrate their high diversity b) emphasizing their economic potential with respect to the abundance of phytogenetic ressources c) evaluating their role as catchment areas for the rivers which are the primary source of water for agriculture and urban life on the coast.
Keywords: montane relict forest, Andes, Andean Western slopes, Peru, Ecuador
Introduction
Northwest Peru and Southwest Ecuador possess some unique relict forests. These have been repeatedly mentioned and discussed in the literature (e.g., Dillehay & Netherly, 1983; Dillon, 1993; 1994; Dillon & Cadle, 1991; Dillon et al., 1995; 2002; Ferreyra, 1977; Koepcke & Koepcke, 1958; Koepcke, 1961; Arana, 1994; Valencia, 1992; Weberbauer, 1945; Young & Reynel, 1997; Sagástegui & Dillon, 1991; Sagástegui et al., 2003). There is general consensus, that these forests have outstanding importance for the conservation of biodiversity, but also as catchment areas for the rivers running down to the coast. The present volume wants to address a wide range of topics, providing better understanding of these forests, some ideas on their importance, research needs and aspects of conservation. These relict forests are concentrated in the so-called Amotape-Huancabamba Zone (Weigend, 2002; 2004), an area of outstanding biodiversity where the flora of the northern and central Andes overlap and additionally a stunning amount of endemic species and genera are found. In this transitional area between the northern and the central Andes the climate does not allow a complete forest cover of the western slope, as it does further north, but these forests are naturally restricted to those areas which receive high precipitation. It seems plausible to assume that under different climatic conditions these forests where at some stage more or less coherent, at least to the southern limit of the Amotape-Huancabamba Zone (where the deep and arid Río Santa valley probably always formed a natural barrier to forest taxa).
Montane forests
About 2600 and 3200 m of altitude on the arid western slopes of the Central and Northern Andes, montane forests are localized (Weberbauer, 1945; Koepcke, 1954; Valencia, 1992). Although these forests can be considered parts of a near continuous strip in the past, however actually just some hundreds of relict forests are presented.
These relict forests present a strong gradient of their characteristics, with a increment in patch size, species richness, trees abundances and structural complexity toward north of distribution. These changes are consequence of the latitudinal gradient of the climatic conditions on the western slopes of the Andes with more humid and warm climate toward north.
In this volume we nearly exclusively consider the montane relict forests of the NW Peru and SW Ecuador, and leave the tropical dry forests of the lower regions and the mangrove forests largely aside (contributions by Linares-Palomino and Charcape & Moutarde). These montane forests are what has generally been refered to in the literature on these relict forests (Figs. 1 a 3). They are generally considered as the southernmost fragments of the montane forests of the western slopes of the Andes, which is (or was) largely continuous in Ecuador and Colombia, but is highly fragmented in the region under consideration. The small-scale climatic conditions responsible for the presence or absence of montane forests can not usually be measured by classical climatological means, but requires fine-scale methods, one of which is presented by Richter & Moreira-Múñoz (infra). The cloud forests are often found very locally where unusually strong condensation takes place (Figs. 4 y 5).





Each of these relict forests tends to have its own suite of endemic species in plant groups (Gentry, 1992; Cano & Valencia, 1992) such as the Solanaceae (e.g., Leiva, 1996; Leiva et al., 1998a; 1998b; 1998c; 1998d; 2003; Leiva & Mione, 1999; Leiva & Quipuscoa, 1998), Asteraceae (e.g., Dillon & Sagasteguii, 1994; Sagasteguii & Dillon, 2001; Sagasteguii & Quipuscoa, 1998; Sánchez & Dillon, 2000) and Loasaceae (Dostert & Weigend, 1999; Rodríguez & Weigend, 1999; 2004; Weigend & Rodríguez, 2003; Weigend et al., 1998; 2004a; Weigend, 2000a; 2000b; 2004), and Alstoemeriaceae-Bomarea (Hofreiter & Rodríguez, infra) but also in many animal groups (Dillon & Cadle, 1991; Duellman, 1979; Duellman & Pramuk, 1999; Cadle & Chuna, 1994; Cadle & Mc Diarmid, 1990, Franke, 1992; Salinas et al., 2003). Flanagan et al. (infra) provide an overview over the avifauna of the relict forests of NW Peru and show the exceptional wealth of the forests of Ayavaca underscoring the importance of these forest fragments for bird life.
The double nature of the relict forests
The relict forests of northwestern Peru are very rich in endemic species (see citations above, and Hensold, 1999) and the numerous endemic taxa in themselves are proof of the fact that some of these forest fragments must be natural fragments and that evolution of narrowly endemic taxa took place in them. However, some part of the fragmentation is undoubtedly man-made and there is good reason to believe that some natural forest fragments have vanished altogether and others were further fragmented into (man-made) relics of (natural) relict forests. Very little is known about the natural extent of these forests or to what degree pre-colombian cultures already decimated their extent: There are some important sites with pre-incaic ruins in or very close to these forests, e.g., next to the bosque San Mateo (Contumazá), in the Río Zaña valley (Dillehay & Netherly, 1983) and in Ayavaca (Vellinga et al., 2004).
However, there is general agreement that there has been a rapid process of deforestation, which has rather accelerated than stopped in recent years and more than 90% of the forests may already have been destroyed (Figs. 6 a 10).
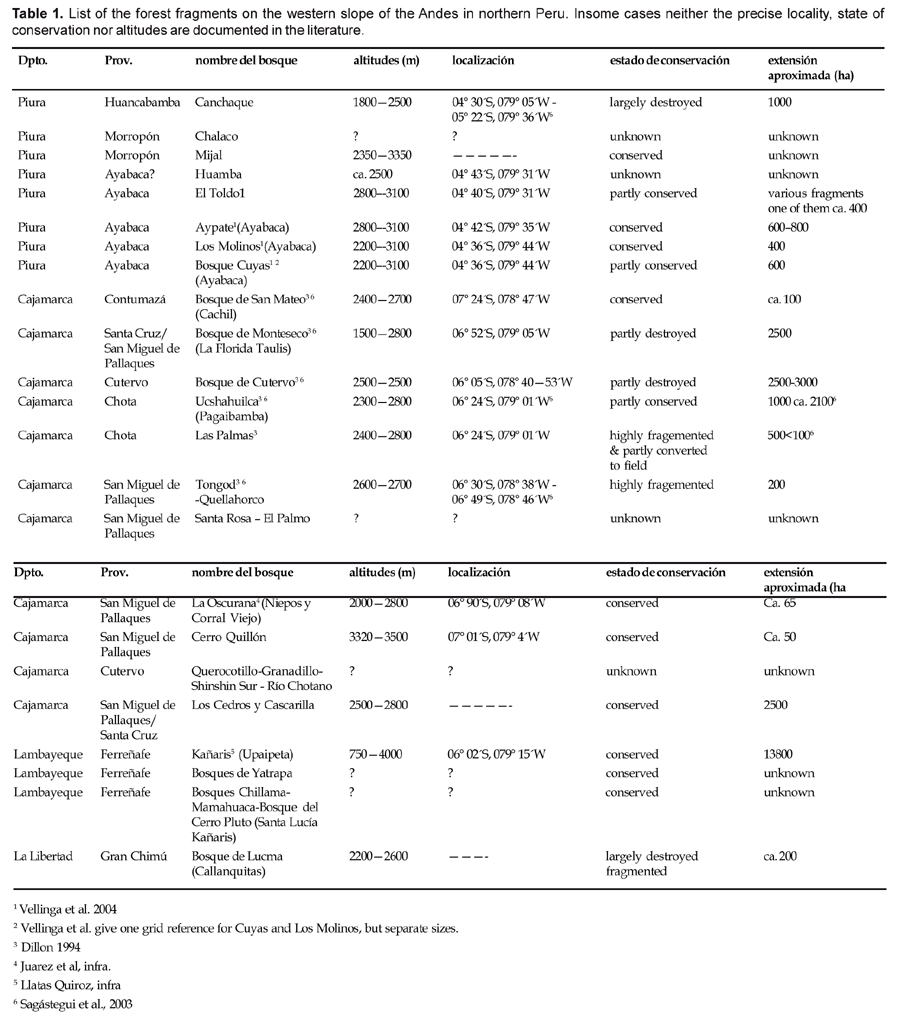
Thus the current relict forests are the isolated remnants of isolated natural forest vegetation and require our urgent attention with respect to investigation and conservation. Table 1 summarizes the known forest fragments from the Peruvian part of the Amotape Huancabamba Zone and give their rough location and extent. In some cases the forest fragments are known from hearsay only and precise localities could not be obtained localizing and investigating them should be a focus of our attention in the immediate fut





Particularly tenacious elements of the montane forest flora such as many members of Solanaceae (Leiva, 1996; Leiva et al., 1998a; 1998b; 1998c; 1998d; 2003; Leiva & Mione, 1999; Leiva & Quipuscoa, 1998), Urticaceae and Ribes (Weigend et al., infra) persist long after the forests have been replaced by fields and hedges. Their distribution can thus be indicative of former forest cover of a region and the data on Urtica and Solanaceae indicate that there used to be very extensive forests in Prov. Otuzco (La Libertad) where no well preserved forests are now left at all.
How many forest fragments are there and what do we know about them?
The literature (e.g., Dillehay & Netherly 1983, Dillon 1993, 1994, Dillon & Cadle 1991, Dillon, et al. 1995, 2002, Ferreyra 1977, Koepcke & Koepcke 1958, Koepcke 1961, Weberbauer 1945; Valencia, 1992; Sagástegui et al., 2003) deals principally with a few well-known forest fragments, typically those of San Mateo, Monte Seco, Canchaque and Cutervo. This covers only a fraction of the total of over 20 forest fragments currently known and in this volume first data are given on two new, well-preserved and hitherto completely ignored forest fragments, the bosque de Oscurana and - most importantly - the forests of Kañaris (Llatas & López, infra). The latter are by far the single largest forest fragment and alone make up possibly more than a 1/3rd of the entire preserved area of montane forests in NW Perú. Investigating this little-known fragments should be first priority in the future.
Two studies on Podocarpaceae (Vicuña) and Orchids (Calatayud) document the enormous floristic wealth of these forest fragments in a species level and also show that the actual diversity far exceeds what has been previously documented in the literature. The herbaria of the region house a considerable wealth of specimens, and so do some herbaria overseas, but the conclusion from revisionary studies (Weigend et al., infra) is that a very considerable part of the forests is still largely or completely unknown: common elements such as Ribes or Urtica are not, or incompletely documented from the majority of forest fragments. Even some well-known forests, such as the forest of Cutervo, are certainly incompletely inventoried. Collection effort have to be intensified and spread evenly over the year to complete our inventories and to provide material for the systematical studies: Phenology in the forest fragments is complex, and many taxa are incompletely documented because they can not be found throughout the year (Bomarea: Hofreiter, Ribes: Weigend).
We expect that numerous new species from these forest relics still await their discovery, either in herbarium collections, where they are mis-filed under names of common and wide-spread congeners, or in the field, where many collection gaps still exist.
What are the relationships between these forests and are some of them particularly diverse?
Investigations on the relationships between the floras of these forests and to the montane forests of Ecuador and the western slope in Peru have been carried out at genus level (e.g., Dillon et al., 1995). This approach can give valuables clues on a larger scale, but the crucial patterns on disjunction are probably reflected only in the the relationships at species level in genera with narrowly endemic taxa such as Larnax (e.g., Leiva, 1996; Leiva et al., 1998b; 1998d), Jaltomata (e.g., Leiva & Mione, 1999; Leiva et al., 1998a), Iochroma (e.g., Leiva & Quipuscoa, 1998; Leiva et al., 1998c; 2003), Nasa (e.g., Dostert & Weigend, 1999), Ribes or Urtica (Weigend et al., infra) or the Passiflora lobbii group (Skrabal et al., 2002) with few exceptions the subgenus Bomarea s.str. and without exceptions subgenus Sphaerine (both Alstroemeriaceae) have in the relict forest their southern most distribution limit in the Cordillera Occidental (Hofreiter & Rodríguez, infra).
However, more or less comprehensive collections will be required before this can be sensibly untertaken. The present data very tentatively suggest, that there is some degree of closer relationship between the relict forests of Otuzco and Contumazá on the one hand, and the relict forests of San Miguel, Santa Cruz and Cutervo on the other. The relict forests of Ayavaca seem to be fairly closely related to those of Huancabamba and San Ignacio, which are more or less the westernmost outliers of the eastern slope forests (Rodriguez et al., 2002; Calatayud, infra), but they also appear to have close ties to the north (e.g., Sessea, Beltran & Galan, 2001). Also, the diversity of these last forests this related to the good conserved forests of the South of Ecuador (Loja and Zamora-Chinchipe), which indicates that in the past these were an all continuous one (Bussmann, 2003; Lozano et al., 2003; Rodríguez et al., 2005, en prep.)
The crucial forests of Kañaris are at present virtually unknown and will play a central role in an understanding of the relationships between the relict forests of NW Peru in the future.
In spite of various efforts to classify the forests according to different levels of diversity, our data on the taxa actually present are probably to scanty to allow us to safely judge relative levels of diversity. It is not surprising that the particularly well-known forests of Monte Seco and Contumazá have a high number or taxa reported, but it seems unlikely that the huge forests of Kañaris will stay far behind and taxa previously considered as endemics of Monte Seco are only now being reported from other places, which had not been explored previously (Nasa insignis, N. humbolditana subsp. obliqua, Juárez et al., infra).
What are the dynamics of these forests and how can it be characterized?
The term relict forests implies that they are more or less «refugia» and stable habitats with little dynamicity. Strangely enough, however, the plant taxa with the highests incidences of endemism and the most narrow endemics (some Asteraceae, Solanaceae, Loasaceae) are nearly exclusively plants of disturbed sites and regular habitat disturbance in the form of, e.g., land slides may play a crucial role in generating and preserving the high diversity. Lozano & Bussmann (infra) explore the effect of land slides on the dynamcis of the forests in S Ecuador and we can safely assume that similar mechanisms are at work in the relict forests in NW Peru. An attempt for a phytosociological classification of the forests is provided by Bussmann (infra) and this should be seen as guide line for a more scientific classification of the relict forests in NW Peru in the mid-term, that goes beyond the rather bland classification as «montane forest».
How can their conservation be achieved?
The ecological and economical role of these forests and their extraordinary wealth in endemic taxa of both plants and animals very strongly argues for measures for their preservation. Charcape et al. (infra) explain how a wide range of activities involving large parts of the population can generate public awareness and prepare the ground for conservation measures. Clearly this approach is especially suited to relatively small forest fragments belonging to an individual municipality. Other measures, higher up in state autority, will be required to protect thus huge tracts of forest as the forest of Kañaris. Each region has its very particular problems and land-owner ship and unclear responsibilities as in the case of the forests of Monteseco which span three administrative units abound. One of the important aspects of these forests is the presence of potential agricultural and pharmaceutical ressources (Sagástegui 1996, Sagástegui & Dillon 1998) which are not at present at all appreciated.
The crucial aspects for achieving the conservation of these remnants of forest are evidently
a) investigating and show-casing their enormous biodiversity,
b) underscoring the potential economical worth of the phytodiversity of the region
c) emphasizing their role as catchment areas for the rivers which drive agriculture and urban life on the coast.
Literature cited
Arana, C. 1994. Biogeografía, Ecología y Conservación del Bosque Montano del Perú. Boletín de Lima 16: 12-13. [ Links ]
Beltran S., H. & A. Galan de Mera 2001: Nuevas adiciones a la flora del Perú, II. Arnaldoa 8(3): 6366.
Bussmann, R.W. 2002. Estudio fitosociológico de la Vegetación en la Reserva Biológica San Francisco (ECSF), Zamora-Chinchipe, Sur de Ecuador. Publicaciones Herbario LOJA N°8. 106 p.
Cadle, J.E. & R.W. McDiarmid. 1990. Two new species of Centrolenella (Anura: Centrolenidae) from northwestern Peru. Proc. Biol. Soc. Wash. 103: 746-768.
Cadle, J.E. & P. Chuna. 1994. A new lizard of the genus Macropholidus (Teiidae) from a relictula humid forest of northwestern Peru, and notes on Macropholidus ruthveni Nobel. Breviora 5001: 1-39.
Cano, A. & N. Valencia. 1992. Composición florística de los bosques nublados secos de la vertiente occidental de los Andes peruanos. Memorias del Museo de Historia Natural, UNMSM (Lima) 21:171-180.
Dillehay, T.D. & P.J. Netherly 1983. Exploring the upper Zana Valley in Peru: A unique tropical forest setting offers insights into the Andean past. Archaeology 34/4: 2230.
Dillon, M.O. 1993: Análisis florístico del Bosque Monteseco (Cajamarca, Perú) e implicancias para su comparación. Arnaldoa. Vol II (2): 29-42.
Dillon, M. O. 1994: Bosques húmedos del norte del Perú. Arnaldoa 2: 2942.
Dillon, M.O. & J. E. Cadle 1991: Bosque Monteseco: A cloud forest above the Peruvian desert. Bull. Field Mus. Nat. Hist. May/June 1: 14.
Dillon, M.O. & A. Sagasteguii 1994: Estudios en el tribu Liabeae (Asteraceae) en Peru: II. Un anueva espécie de Oligactis procedente del Norte del Peru y del Sud del Ecuador. Arnaldoa 2(2): 2530.
Dillon, M.O. & A. Sagasteguii 1994: Estudios en el tribu Liabeae (Asteraceae) en Peru: II. Una nueva espécie de Oligactis procedente del Norte del Peru y del Sud del Ecuador. Arnaldoa 2(2): 2530.
Dillon, M. O., A. Sagástegui A., I. Sánchez V., S. Llatas Q. & N. Hensold. 1995. Floristic inventory and biogeographic analysis of montane forests in northern Peru. Pages 251270 in S. P. Churchill, H. Balslev, E. Forero & J. L. Luteyn (eds.). Biodiversity and Conservation of Neotropical Montane Forests. The New York Botanical Garden Press, Bronx.
Dillon, M.O., I. Sánchez, G. Iberico, A. Sagástegui, M. Zapata & M. Kawasaki. 2002. Biogeografía, diversidad florística y endemismos en dos tipos de bosques en el norte del Perú. Arnaldoa 9(2): 111-120.
Dostert, N. & M. Weigend. 1999. A synopsis of the Nasa triphylla complex (Loasaceae), including some new species and subspecies. Harvard Papers in Botany 4: 439467.
Duellman, W. E. 1979. The herpetofauna of the Andes: Patterns of distribution, origin, differentiation, and present communities. Pages 1485 in Duellman, W. E. (ed.), The South American Herpetofauna: Its Origin, Evolution, and Dispersal. Univ. Kansas Mus. Nat. Hist. Monogr. 7.
Duellman, W. E. & J. B. Pramuk. 1999. Frogs of the genus Eleutherodactylus (Anura: Leptodactylidae) in the Andes of northern Peru. Scientific Papers Univ. Kansas Mus. Nat. Hist. 13: 178.
Ferreyra, R. 1977: Endangered species and plant communities in Andean and coastal Peru. pp. 150157 In: G.T. Prance & T.S. Elias (eds.) exctinciton is forever. NYBG Press.
Franke, I. 1992. Biogeografía y Ecología de las aves de los bosques montanos del Perú occidental. Memorias del Museo de Historia Natural, UNMSM (Lima) 21:181-188.
Gentry, A.H. 1992. Diversity and floristic composition of Andean forests of Peru and adjacent countries: Implications for their conservation. Memorias del Museo de Historia Natural, UNMSM (Lima) 21:11-29.
Hensold, N. 1999. Las angiospermas del Departamento Cajamarca, Perú. Arnaldoa 6: 141184.
Koepcke H.W. & M. Koepcke 1958: Los restos de bosques en las vertientes occidentales de los Andes peruanos. Bol. Comite Nac. Protecc. a la Naturaleza (Peru) 16: 2230.
Koepcke, H.W. 1961. Synökologische Xtudien an der Westeseite der peruanischen Anden. Bonner Geogr. Abh. 29: 1320.
Koepcke, M. 1954. Corte Ecológico transversal en los Andes del Perú central con especial consideración de las aves. Parte I: Costa, Vertiente occidentales y Región Altoandina. Memorias del Museo de Historia Natural, UNMSM (Lima) 3:1-119.
Leiva G., S. 1996: Dos nuevas especies de Larnax (Solanaceae: Solaneae) del Norte de Perú. Arnaldoa 4(1): 1522.
Leiva G., S., & T. Mione 1999: Dos nuevas espécies de Jaltomata Schlechtendal (Solanaceae: Solaneae) del Norte de Peru. Arnaldoa 6(1): 6574.
Leiva G., S. & Quipuscoa S., V. 1998: Iochroma nitidum y I. schjellerupii (Solanaceae: Solaneae) dos nuevas especies andina del Norte de Perú. Arnaldoa 5(2): 171-178.
Leiva G., S., T. Mione & Quipuscoa S., V. 1998a: Cuatro nuevas espécies de Jaltomata Schlechtendal (Solanaceae: Solaneae) del Norte de Perú. Arnaldoa 5(2): 179-192.
Leiva G., S., V. Quipuscoa S. & N. W. Sawyer 1998b: Nuevas espécies andinas de Larnax de Ecuador y Perú. Arnaldoa 5(1): 8392.
Leiva G., S., V. Quipuscoa S. & N. W. Sawyer 1998c: Iochroma stenanthum (Solanaceae: Solaneae) una nueva espécie del Norte de Perú. Arnaldoa 5(1): 77-82.
Leiva G., S., E. Rodríguez R. & J. Campos de la Cruz 1998d: Cinco nuevas especies de Larnax (Solanaceae: Solaneae) de los bosques montanos del Norte de Perú. Arnaldoa 5(2): 193210.
Leiva G., S., P. Lezama A. & V. Quipuscoa S. 2003: Iochroma salpoanum y I. squamosum (Solanaceae), dos nuevas espécies andinas del Norte de Perú. Arnaldoa 10(1): 95104.
Lozano, P.; T. Delgado & Z. Aguirre. 2003. Estado Actual de La Flora Endémica exclusiva y su distribución en el Occidente del Parque Nacional Podocarpus. Publicaciones de la Fundación Ecuatorina para la Investigación y Desarrollo de la Botánica. Loja. Ecuador.
Rodríguez, E., E. Alvítez & S. Leiva. 2002. Estudio de la Flora Vascular de la Provincia de San Miguel Pallaques, Dpto. Cajamarca-Perú (II). Libro de Resúmenes del IX CONABOT, Iquitos-Perú. p. 85.
Rodríguez, E. & M. Weigend. 1999. Nasa umbraculifera (Loasaceae: Loasoideae), una nueva especie con hojas peltadas del Perú. Arnaldoa 6(1): 49-56.
Rodríguez, E. & M. Weigend. 2004. Nasa longivalvis (Loasaceae: Loasoideae), una nueva especie del Departamento de La Libertad, Perú. Arnaldoa 11(2): 67-78.
Sagástegui, A. ; S. Leiva; P. Lezama; N. Hensold & M.O. Dillon. 1995. Inventario Preliminar de la Flora del Bosque de Cachil. Arnaldoa 3(2). 19-34.
Sagasteguii, A. 1996: El «gashmin» (=Eugenia quebradensis): un nuevo recurso alimenticio. Arnaldoa 4(1): 47-56.
Sagasteguii, A. & M. O. Dillon. 1991. Inventario Preliminar de la Flora del Bosque de Monteseco. Arnaldoa 1(1). 35-52.
Sagasteguii, A. & M. O. Dillon 1998: Una nueva espécie de Chrysophyllum (Sapotaceae) con frutos comestibles del noroeste de Perú. Arnaldoa 5(1): 67-76.
Sagasteguii, A. & Dillon, M.O. 2001: Una nueva especies de Critoniopsis (Vernonieae: Asteraceae) del norte de Perú. Arnaldoa 8(1): 2530.
Sagasteguii, A. & Quipuscoa S., V. 1998: Tres nuevas especies de Verbesina (Asteraceae: Heliantheae) de los Andes del Perú. Arnaldoa 5(2): 141-149.
Sagástegui, A.; I. Sánchez-Vega; M. Zapata &.O. Dillon. 2003. Diversidad Florística del Norte de Perú: Bosques Montanos. Graficart. Tomo II. 3005 p.
Salinas, L.; Samame, M.; Franke, I. & J. Fjeldsa. 2003. First record of Black-chested Fruiteater Pipreola lubomirskii (Aves, Cotingidae) in the western slopes of the Andes. Rev. peru biol.10: 93-97.
Sánchez V., I. & Dillon, M.O. 2000: Una nueva especie de Mikania (Eupatorieae: Asteraceae) del norte de Perú. Arnaldoa 7(1-2): 712.
Skrabal, J., Tillich H.-J., Weigend, M. 2001: A revision of the Passiflora lobbii group (Passifloraceae), including some new species and subspecies. Harvard Papers in Botany 6: 309339.
Valencia, N. 1992. Los bosques nublados secos de la vertiente occidental de los Andes del Perú. Memorias del Museo de Historia Natural, UNMSM (Lima) 21:155-170.
Vellinga, W.-P-, J. N. M. Flanagan & T. R. Mark 2004: New and interesting records of birds from Ayabaca province, Piura, north-west Peru. Bull. B. O.C. 124(2): 124142.
Weberbauer, A. 1945. El Mundo Vegetal de los Andes Peruanos. Estudio Fitogeografico. Est. Exp. Agr. La Molina. Lima-Perú. 776 p.
Weigend, M. 2000a. A revision of the Peruvian species of Nasa ser. Alatae. Nord. J. Bot. 20: 1532.
Weigend, M. 2000b. Loasaceae. In: L. Andersson & G. Harling (eds.), Flora of Ecuador 64. 192.
Weigend, M. 2002. Observations on the Biogeography of the Amotape-Huancabamba Zone in Northern Peru. - En: K. Young, C. Ulloa U., J. L. Luteyn & S. Knapp, Plant Evolution and Endemism in Andean South America. - Bot. Review 68(1): 3854.
Weigend, M. 2004. Four New Species of Nasa ser. Alatae (Loasaceae) in the Amotape-Huancabamba Zone of Peru. Novon 14: 134146.
Weigend, M. & E. Rodriguez R. 2001. Nasa picta (Hook.f.) Weigend subsp. pamparomasii (Loasaceae), a new subspecies of Nasa picta from Ancash, Peru. Arnaldoa 7(12): 1926.
Weigend, M. & E. Rodriguez R. 2003. A revision of the the Nasa stuebeliana group [Nasa ser. Saccatae (Urb. & Gilg) Weigend, Loasaceae] with notes on morphology, ecology, and distribution. Bot. Jahrb. Syst. 124(4): 345382.
Weigend, M., E. Rodríguez R. & N. Dostert. 1998. Nasa insignis y Nasa glandulosissima, dos especies nuevas de Nasa con hojas peltadas del norte del Perú. Arnaldoa 5: 151157.
Weigend, M., T. Henning, & C. Schneider. 2004a. Notes on the Systematics, Morphology, Distribution and Pollination of Nasa Ser. Carunculatae (Loasaceae subfam. Loasoideae). - Syst. Bot. 29(1): 765781.
Young, K. R. & C. Reynel 1997. Huancabamba Region, Peru and Ecuador. Pages 465469 in S.D. Davis, V.H. Heywood, O. Herrera-MacBryde, J. Villa-Lobos & A.C. Hamilton (eds.), Centers of Plant Diversity, a Guide and Strategy for their Conservation, Vol. 3. The Americas. IUCN Publications Unit, Cambridge.
Weigend, M. 2004: Additional Observations on the Biogeography of the AmotapeHuancabamba Zone in Northern Peru: Defining the South-Eastern limits. Rev. Peru. Biol. 11(2): 127-134
Correspondencia
1Institut für Biologie Systematische Botanik und Pflanzengeographie, Freie Universität Berlin
E-mail:weigend@zedat.fu-berlin.de
2Herbarium Truxillense (HUT), Universidad Nacional de Trujillo, Trujillo-Perú
E-mail:efrr@unitru.edu.pe
3Museo de Historia Natural y Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos
E-mail:caranab@unmsm.edu.pe
