










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.12 n.2 Lima ago./set 2005
Michael Richter* y Andrés Moreira-Muñoz
*Institut für Geographie, Friedrich Alexander Universität - Erlangen, Alemania.
Número especial: Bosques relictos del NO de Perú y SO de Ecuador
Weigend, Rodríguez y Arana (Comps.)
Publicado online: 08/12/05
Resumen
En el presente trabajo se propone un método para obtener informaciones higrotérmicas de alta resolución espacial con la ayuda de epífitas como indicadores, junto con las medidas de la temperatura del suelo en una profundidad de 60 cm, comparadas con datos de estaciones meteorológicas. El método provee mayores antecedentes para descifrar el funcionamiento del sistema regional de circulación atmosférica, así como para realizar una clasificación climática detallada. La resolución espacial del método permite además cartografiar la distribución de la vegetación natural potencial. Ello puede significar un aporte para la comprensión de los complejos factores ambientales que soportan altos niveles de riqueza florística y endemismo, por lo cual el sur de Ecuador es reconocido como uno de los centros de diversidad florística más importantes en el Neotrópico y también a escala global. El método resulta potencialmente aplicable en el norte del Perú, donde existe la misma situación de compleja orografía.
Palabras clave: fitoindicación, epifitismo, bioclimatología, Bosque Montano Tropical, Ecuador.
Abstract
In this paper hygric estimations by similarity analysis based on epiphytism are combined with singular measurements of temperatures 60 cm below ground to generate an accurate cartography of hygro-thermal patterns within a complex relief in southern Ecuador. The method improves comprehension of regional circulation patterns as well as providing an idea of the potential vegetation distribution. This can be of further help to comprehend the various environmental factors that support one of the highest levels of floral diversity in the Neotropics and even on the global scale. The method described is also applicable for the Andean part of northern Peru with its similarly complex orography.
Key words: Phytoindication, epiphytism, bioclimatology, Neotropical montane forest, Ecuador.
Introducción
La diversidad florística y vegetal del Ecuador ha llamado la atención de botánicos y naturalistas desde hace varios siglos, a partir de las exploraciones de La Condamine (1736-1743) y Jussieu (1735-1747). Luego vendrían las exploraciones de Alexander von Humboldt junto con Aimé Bonpland (1802-1803), que permitirían sentar las bases de la fitogeografía. A partir de aquellos notables precursores, muchas propuestas para la clasificación de la vegetación han sido realizadas hasta la fecha (ver revisión en Valencia et al., 1999a). Aun así, los bosques montanos tropicales, como tales, han sido sólo recientemente investigados en mayor profundidad (ver revisión en Churchill et al., 1995). Es así como los bosques montañosos relictos de los Andes del sur de Ecuador y del norte de Perú han sido recientemente reconocidos como uno de los más importantes centros de diversidad florística del mundo (Barthlott et al., 1999; Young y Reynel, 1997), y como hot spots en relación con su prioridad de conservación (Myers et al., 2000). De hecho esta zona constituye uno de los pocos lugares del planeta donde aún es posible descubrir y describir plantas vasculares nuevas para la ciencia (Weigend, 2002). En este trabajo se analizan y discuten antecedentes bioclimáticos y vegetacionales a través de un enfoque de fitoindicación, lo que permitirá enriquecer los antecedentes para la comprensión de los factores ambientales que favorecen la existencia de este centro de diversidad florística.
Caracterización regional
Uno de los sectores más bajos de la cadena de los Andes se localiza entre la depresión de Girón-Cuenca (Ecuador) y Huancabamba (Perú) (entre 2 y 6° S); muchos consideran esta región como el límite entre los Andes del norte y los Andes centrales (Fig. 1). Se trata asimismo de la zona en que la vegetación sobre el límite arbóreo recibe la denominación de «jalca». Los páramos norandinos son separados de las punas de los Andes centrales por esta formación. Aunque el uso de un término específico para esta zona de transición páramo-puna es difícil de justificar a partir de las comunidades vegetales, así como en cuanto a la composición de especies. Más importante resulta otra característica de los pisos andinos en la zona de Amotape-Huancabamba: esto es, la relativamente baja posición del límite arbóreo. El límite se localiza aquí entre 3200 y 3400 m de altitud, debido a que las especies de Polylepis, características de los pisos andinos altos, desaparecen en esta zona (con excepción de P. weberbaueri, que se presenta en hábitat secos aislados hasta 3600 m; Baumann, 1988). La razón para el desplazamiento altitudinal negativo del límite arbóreo son las extremadamente altas precipitaciones, por lo que la región resulta sólo apta para los elementos más hígricos. Mediciones de precipitación en la Cordillera Real del sur de Ecuador muestran montos de hasta 8000 mm a-1 entre 3000 a 3400 m de altitud (comm. pers. Dr. Paul Emck, Erlangen). Investigaciones mediante percepción remota (radar) en los sectores más inaccesibles de la confluencia de la Cordillera Real con la Cordillera del Cóndor permiten suponer precipitaciones superiores a 15000 mm a-1 (comm. pers. Dr. Rüdiger Rollenbeck, Marburg).
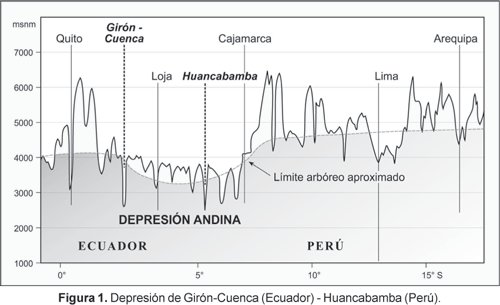
Las precipitaciones extremas y su influencia sobre la configuración del límite arbóreo resultan de las nubes de los alisios provenientes del Este que afectan las vertientes orientales. En todo caso hay que enfatizar que en la depresión andina los sectores altos son extremadamente húmedos durante todo el año y las vertientes de sotavento, en cambio, relativamente secas. Justamente aquí se encuentra un extremo gradiente vertical de precipitación que, junto con la fragmentación del relieve en valles y sierras, conduce a una extraordinaria heterogeneidad climática en la región. Dicha heterogeneidad climática es muy difícil de evaluar con los métodos tradicionales. Debido a que estas características son similares en el norte de Perú, se presenta a continuación una posibilidad metodológica que permite analizar la complejidad climática y fitogeográfica con mayor detalle.
Área de estudio y métodos
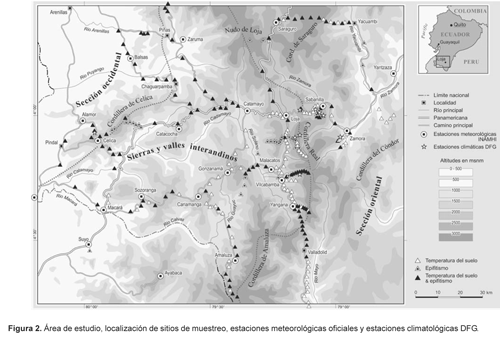
El área de estudio específica que abarca este trabajo se localiza en el sector norte de la depresión andina, y comprende partes de la cordillera occidental, las sierras y valles interandinos, y la cordillera oriental en el sur de Ecuador (Fig. 2). El «Nudo de Loja» culmina en los 3800 m de altitud en Fierro Urcu y Cerro Chinchilla. De allí, las montañas se derivan hacia el S y el SE, interrumpidas por los valles interandinos. Esta región de compleja orografía ocupa 25500 km2 y posee solamente 22 estaciones meteorológicas oficiales, la mayoría de ellas con datos interrumpidos y escasos. Todos los sitios se concentran en los valles y las cuestas. Solamente uno de estos sitios está localizado sobre los 2500 m. La escasa red de estaciones meteorológicas dificulta la obtención de datos exactos para la comprensión del clima en muchas áreas andinas. Esto es especialmente válido para los sectores más húmedos de las montañas, particularmente en los escarpes orientales de los Andes tropicales. Sin embargo, incluso un mayor número de estaciones difícilmente podría expresar la gran complejidad del clima en una región con tanta diversidad de condiciones ambientales, en un radio de acción de pocos kilómetros.
Debido a la escasez de estaciones meteorológicas, hemos aplicado un método alternativo de análisis bioclimático basado en tres aspectos:
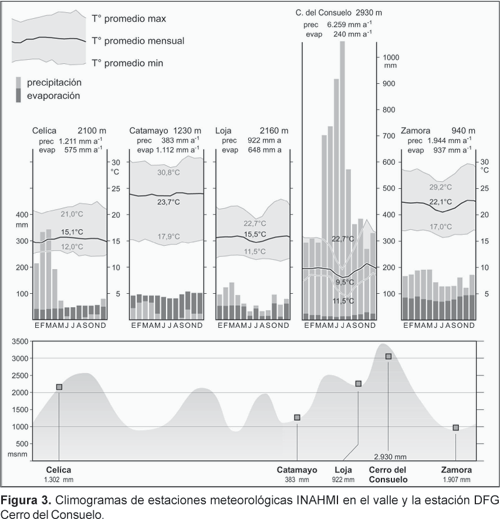
a) Complemento de los datos climáticos de las 22 estaciones meteorológicas INAMHI (dos de ellas en Perú, algunas provistas solamente de pluviómetro) mediante ocho estaciones climáticas automáticas (estaciones DFG) instaladas en lugares de menor accesibilidad. Ello permite caracterizar los pisos altitudinales según la temperatura media anual y la cantidad de meses húmedos (mes húmedo = mes en que la precipitación supera la evaporación, calculada según la fórmula de Penman). En la Figura 3 se aprecia las notables diferencias, sobre todo en precipitación, entre las estaciones del valle y la estación DFG instalada en el Cerro del Consuelo.
b) Fitoindicación mediante epífitas: Se ha analizado la forma de vida epífita como un indicador de diferenciación hígrica espacial. Para ello se han identificado las epífitas creciendo en ramas altas de árboles solitarios o en grupos abiertos. Los ejemplares han sido observados mediante binoculares. Para ello se han realizado 156 muestreos de terreno (relevés) a fines de marzo de los años 1998 a 2002. Cada sitio de muestreo se compone de cinco muestras efectivas en árboles vecinos cubriendo un largo total de rama de 15 ± 3 m. Se desarrolló una clasificación ad hoc compuesta de algunos taxa característicos (e.g. Tillandsia usneoides) y principalmente formas de crecimiento, en total se consideran 25 variables. Se estimó el porcentaje de cobertura de cada taxón o forma de crecimiento particular. Frecuentemente los líquenes y musgos forman superficies compactas alrededor de la rama, sirviendo de sustrato para el crecimiento de plantas vasculares. Es por ello que es común obtener coberturas totales de más de 100%. Con los valores de cobertura se realiza el análisis de ordenación. Los valores de taxa y formas de crecimiento han sido corregidos por la raíz cuadrada para evitar la total dominancia de líquenes o musgos y permitir la inclusión de grupos con menor cobertura en los cálculos. Se localizó expresamente sitios de muestreo en la vecindad de las estaciones INAMHI o DFG, para caracterizar las diferentes clases de epifitismo.
c) Temperatura del suelo: debido a que los atributos de fitoindicación también están afectados por la radiación solar y la disponibilidad de agua a microescala, el método es controlado mediante un parámetro directo casi constante, como es la temperatura del suelo a 60 cm de profundidad, que en áreas tropicales revela la temperatura anual del aire a 2 m sobre el suelo (Winiger, 1981). Se debe colectar los datos en condiciones de similar exposición y pendiente para disminuir los efectos de diferencias de humedad del suelo. Se tomaron 244 muestras de temperatura del suelo mediante un termómetro electrónico portátil. Primero se realiza un agujero de 50 cm de profundidad y luego se entierra el termosensor, llegando así a la profundidad requerida de 60 cm. El trabajo de terreno comprendió la época lluviosa desde diciembre de 1998 a abril de 1999. También se realizaron los correspondientes muestreos en la vecindad de las estaciones meteorológicas, para controlar los resultados.
Los sitios de muestreo de epifitismo y temperatura del suelo, así como las estaciones meteorológicas INAMHI y DFG, se representan en la Figura 2.
La integración de la información nos ha permitido generar una cartografía bioclimática compuesta de tres cartas: 1) carta hígrica, 2) carta térmica, 3) carta de vegetación potencial.
Resultados
Sistema de circulación regional
Presentamos a continuación una explicación sinóptica del sistema de circulación, la cual surge de la integración de la información hígrica y termal obtenida mediante el epifitismo, las temperaturas del suelo y las estaciones meteorológicas. Más antecedentes climáticos han sido expuestos para la zona de estudio por Richter (2003).
La Figura 4 da una idea de las situaciones frecuentes durante julio y febrero. Los vientos del este tempestuosos casi permanentes (easterlies), de la tropósfera media son los responsables del tiempo lluvioso a lo largo de todo el año en las crestas de las cordilleras orientales. Estas corrientes fuertes son el motor principal para la elevación de las masas de aire que divergen de los alisios de la tropósfera inferior. Los alisios pasan al este de la cadena montañosa sobre la Amazonía preandina, de sur a norte en el invierno meridional (julio) y de norte a sur en el verano (febrero). Los mismos flujos de aire causan nubes de turbulencia en las cumbres de las cordilleras, los cuales generan un flujo latente del calor y vientos fuertes descendentes hacia la región interandina. Estos foehns (vientos catabáticos secos y calientes) son especialmente eficientes en las áreas de Loja, Malacatos, Vilcabamba y Yangana. Las cordilleras orientales se caracterizan por las precipitaciones y las sierras interandinas por una situación de sotavento seca durante los easterlies; pero las circunstancias en la vertiente oeste de la cordillera occidental son totalmente diferentes. Aquí, el permanente flujo bajo del aire desde S a N a lo largo del llano costero y las colinas (alisios del Pacífico) es sobrepuesto por la corriente de los mismos vientos del este. Esto da lugar a una «corriente jet» baja con flujos de aire helicoidales que crean la capa de inversión típica entre 1200 y 2400 m de altitud. Neblinas orográficas son cortadas en este punto por los fuertes easterlies y forman una densa capa de nubes que se sitúa sobre las inclinaciones occidentales o traspasa las partes más inferiores de las líneas de cresta.
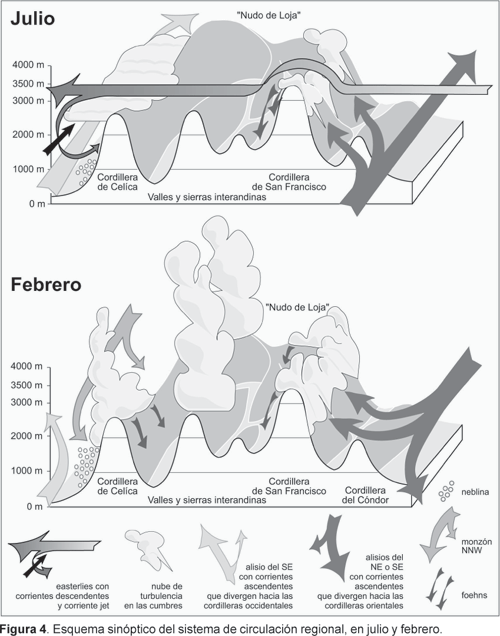
Los easterlies y los alisios que proveen de lluvia a la vertiente amazónica dominan a lo largo de todo el año pero se debilitan en la sierra interandina durante la estación húmeda entre diciembre y abril. Paralelamente, cortinas de nubes pueden cruzar la cordillera occidental del lado del Pacífico hacia el este y conducen los foehns de la mañana desde la Cordillera de Célica hacia los valles interiores. Además existe la posibilidad de una entrada de aire húmedo de los alisios del sur por el valle del río Catamayo (río Chira en Perú). Nubes de tempestad convectiva comienzan eventualmente a elevarse sobre los valles interandinos alrededor de mediodía. Son las únicas fuentes de precipitación para la sección central del área del estudio y pueden ser asignadas a una de las posiciones más australes de la ZCIT (Zona de Convergencia Intertropical) al oeste de los Andes. Un tercer acontecimiento del clima, que hemos denominado «ausencia total de lluvia» se restringe al período de octubre hasta abril, alcanzando la frecuencia más alta durante fines de octubre y noviembre siempre situaciones concentradas por algunos pocos días. Este fenómeno se denomina «veranillo de El Niño», y se traduce en un corto período seco encima de las cordilleras. Cuando se debilita el alisio del S, a veces entra desde el noroeste una corriente monzónica que puede traer lluvias hasta la vertiente oeste de la cordillera occidental.
Fitoindicación
Se han utilizado 25 formas de crecimiento de epífitas como indicadores. En la Figura 5 se ejemplifica con cinco situaciones características y el grado de cobertura para cada variable en la columna de la derecha.


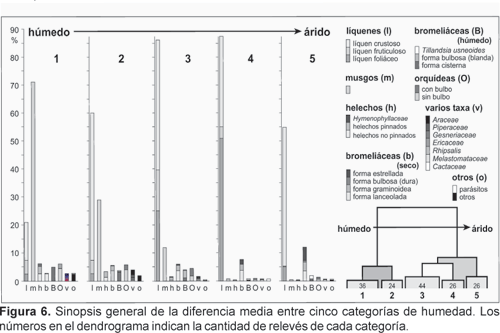
En la Figura 6 se muestra una sinopsis general que indica la diferencia media entre cinco categorías de humedad (pero no hay que perder de vista que en realidad cada relevé forma un grupo para determinar la humedad específica de un lugar). A primera vista, líquenes y musgos parecen ser decisivos debido a los altos valores de cobertura en todas las clases. Sin embargo, el papel significativo de porcentajes más pequeños de plantas vasculares no puede ser negado. Varias formas de crecimiento de bromeliáceas pueden generar una gran diferencia en los agrupamientos luego que su valor de indicación haya sido acentuado por la transformación por la raíz cuadrada. Ello es obvio al comparar los cluster 1 y 5 con los tipos hídricos respectivamente xéricos de bromeliáceas.
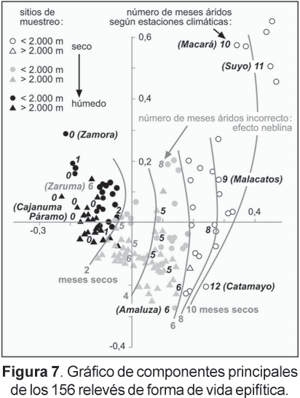
Los 156 relevés de forma de vida epífita han sido organizados mediante un análisis de componentes principales. La figura 7 ilustra el amplio patrón de comunidades epifíticas que comprenden desde áreas perhúmedas (izquierda) a peráridas (derecha). Los números indican los meses húmedos en los sitios localizados al lado de una estación meteorológica. Entre los círculos negros que representan sitios húmedos y los círculos vacíos que señalan sitios secos, los círculos grises del centro y en la base de la curva de forma de herradura indican una serie de localidades intermedias de clima semihúmedo a semiárido. Los números en color gris que no coinciden con las isolíneas se originan en estaciones climáticas con persistente presencia de neblina, y no son expresadas en los datos de precipitación. Por lo tanto, el número real de meses húmedos en estos sitios es más alto de lo representado en la Figura 7.
Carta hígrica basada en la fitoindicación
Cada punto de muestreo o relevé provee un valor local de humedad; estos valores más los de las estaciones se superponen a la carta topográfica, obteniéndose la carta de la situación hígrica para el área de estudio (Fig. 8). Los puntos actúan como estaciones meteorológicas, con la ventaja de aumentar en la práctica de 30 estaciones a 156 puntos de información.
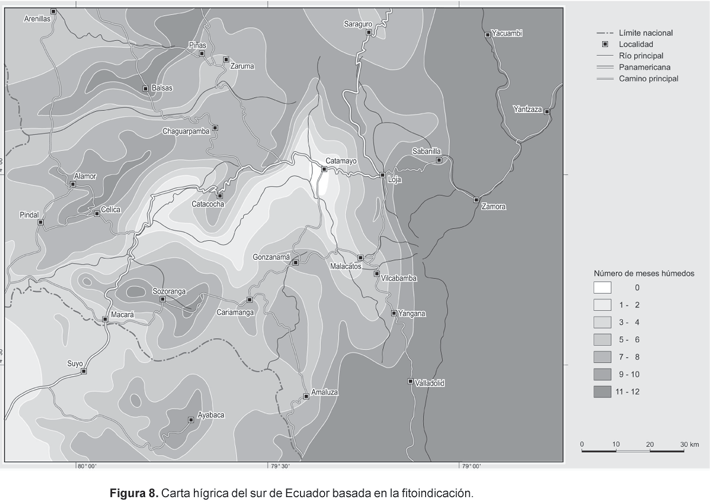
Los patrones que muestra la carta son los siguientes:
· La zona occidental está marcada por patrones de precipitación heterogéneos, pero en menor grado que el centro, debido a que las tierras bajas del Pacífico no son tan secas como los fondos de los valles interiores. Las crestas son menos húmedas que aquellas de las cordilleras orientales. Pero un visible wet spot desconocido hasta la fecha se localiza al oeste de Piñas y no coincide para nada con los registros de alguna estación cercana (Zaruma 6, Balsas 7 meses húmedos). Las condiciones subhumedas a perhúmedas señaladas por un tipo de epifitismo similar al de la región de Zamora al este de la Cordillera Real, se encuentran inducidas por las frecuentes neblinas durante la época de sequía.
· La zona central intermedia de valles y sierras está determinada por condiciones hígricas complejas. Mientras que los aguaceros durante la época húmeda proveen lluvias en las elevaciones superiores, los mismos eventos no necesariamente tocan las zonas bajas, debido a altas tasas de evaporación en los valles al mismo tiempo de la caída de lluvia. Adicionalmente, efectos de elevación de masas de aire causados por procesos convectivos resultan en una mayor y más frecuente precipitación en las cumbres («efecto Merriam», según Richter, 1996). Un evidente «dry spot» está centrado alrededor de la cuenca de Catamayo. Los valores anuales de precipitación (383 mm a-1) son ligeramente menores que en Macará, localizada 800 metros más abajo (427 mm a-1), y un poco más altos que en Zapotillo (368 mm a-1). Ambas estaciones están situadas en las tierras bajas en el límite con Perú. Sin embargo, el mes más húmedo en Catamayo aún muestra déficit hídrico en contraste con al menos un mes húmedo en Macará y Zapotillo.
· La sección oriental hasta la cresta de la Cordillera de Amaluza, la Cordillera Real y la Cordillera de Saraguro se caracteriza por climas perhúmedos y subhúmedos, debido al constante efecto de los alisios. En este punto se debe destacar que la distancia entre el sitio más seco y el más húmedo comprende menos de 30 km, desde las cumbres de la Cordillera Real hasta Catamayo (probablemente más de 6000 mm a-1 versus 383 mm a-1).
Temperatura del suelo
Fue necesario un ajuste según la estación, debido a que amplitudes de temperatura de 2-3 K son típicas en el curso anual y también interanual. En la época de medición las estaciones climáticas aún mostraban anomalías positivas producidas por efectos residuales de El Niño 1997-98. Las temperaturas en crestas elevadas fueron alrededor de 1 K y en los valles interandinos de hasta 3 K más altas que en los años no-Niño. La corrección estacional e interanual se logra mediante mediciones continuas en las estaciones DFG y con seis mini loggers en los respectivos 60 cm. Teniendo los datos corregidos es posible realizar los análisis expuestos en la Figura 9.
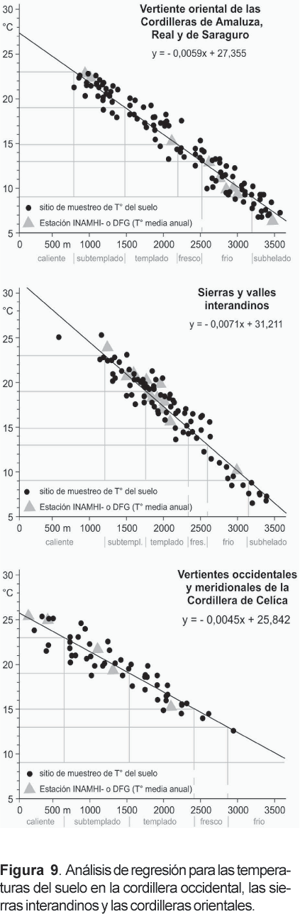
En la figura 9 los sitios de estudio han sido divididos de acuerdo con las tres principales subregiones (cordilleras orientales, sierras interandinas, cordillera occidental). La fórmula de regresión muestra un claro gradiente vertical de temperatura. Precipitaciones generadas por los alisios prevalecen a lo largo de los escarpes orientales de las cordilleras del este y conducen a condiciones adiabáticas húmedas expresadas en una disminución vertical de 0,59 K/100 m (Fig. 9a). En contraste, el gradiente de temperatura calculado para los valles interandinos (Fig. 9b) se caracteriza por condiciones secas reflejadas en una disminución vertical de 0,71 K/100 m. De hecho, la mayor pendiente del gráfico refleja la importante alteración altitudinal de los procesos climáticos: mientras que la tierra caliente muestra evidencia de cuencas cálidas y fondos de valle, en elevaciones de hasta 3000 m, precipitaciones más frecuentes y nieblas producen efectos de enfriamiento en las cumbres.
Finalmente, la vertiente occidental y las faldas meridionales de la Cordillera de Celíca (Fig. 9c), que disminuye hacia el desierto del norte peruano, son marcadas por un gradiente asombrosamente bajo de 0,45 K/100 m. Tales valores corresponden a los resultados de gradientes especialmente pequeños típicos para «desiertos tropicales de aire húmedo» (Lauer 1995; Weischet 1996). Si bien las surgencias de aguas frías usualmente no tocan la costa pacífica del sur de Ecuador, aires frescos cruzan la precordillera de Tumbes y el desierto costero de Sechura en Perú, ingresando a lo largo de las precolinas de la Cordillera de Celíca. Esto explica las menores temperaturas de las tierras bajas hasta los niveles inducidos por la niebla, que se extienden entre 1000 y 2400 m de altitud según la estación y hora del día (regiones de calor deficitario). Como el impacto orográfico sobre la niebla se reduce en altura, las áreas de cumbre sobrepasan el nivel de masas de aire advectivo frío y forman islas relativamente cálidas sobre el nivel de la inversión (regiones de calor surplus).
Los diferentes gradientes que muestra la Figura 9 implican regímenes de temperatura complejos. Los análisis de las desviaciones positivas y negativas de los valores de temperatura muestran distribuciones asimétricas de los pisos térmicos (anotados bajo el eje x). Las diferentes extensiones espaciales de los regímenes térmicos se representan en la carta de diferenciación térmica.
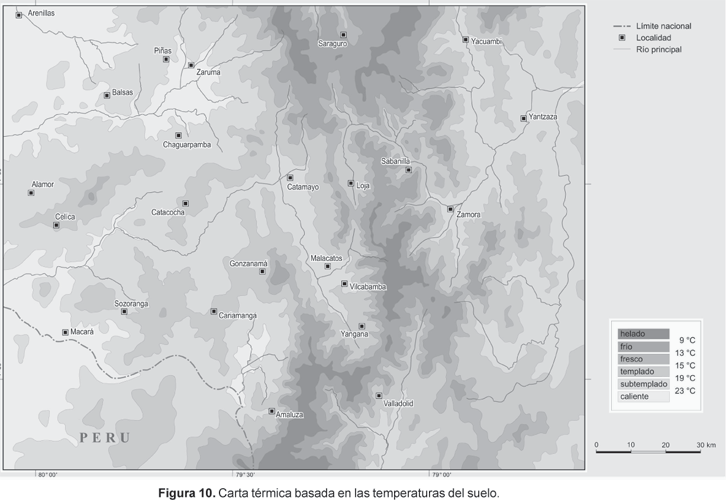
Carta térmica
En la Figura 10 se representa la carta térmica. En la parte occidental del área de estudio, la tierra caliente se reduce a una capa baja hasta ca. 700 m de altitud debido al mencionado impacto de frío. El mismo piso alcanza hasta los 1200 m en los valles interandinos. Las áreas con las desviaciones positivas de la temperatura que superan +2 K se concentran alrededor de la cuenca de Catamayo y a lo largo del sur de los valles de Gonzanamá, abrigado por múltiples cadenas e influenciado por fuertes vientos foehn que descienden de la Cordillera de Amaluza. En contraste, el área más húmeda a lo largo de la cresta de la Cordillera Real y especialmente en la parte norte del Parque Nacional Podocarpus entre Loja y Zamora, así como el casi permanente wet spots inducido por la niebla al oeste de Piñas, se caracterizan por las desviaciones negativas de la temperatura de más de 2 K. Nuevamente, los sitios más calientes (hot spots) y más fríos (cold spots) se sitúan cerca los unos de los otros, y coinciden con los ya mencionados sitios más secos (driest spot) y húmedos (wettest spot) lo cual enfatiza la relación termal con áreas de superávit y déficit de agua.
Tipos climáticos higro-termales
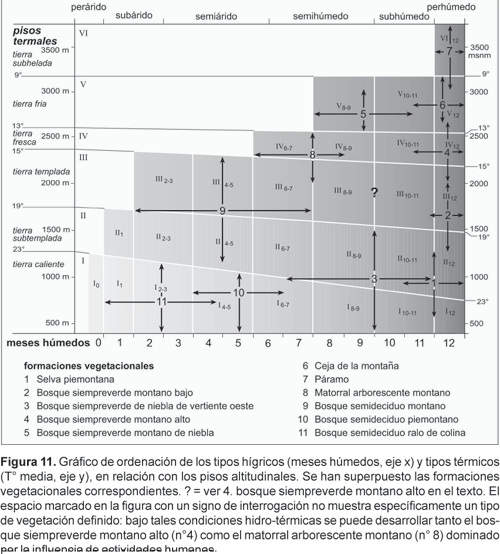
Teniendo los tipos hídricos y térmicos para el área de estudio, es posible generar un gráfico de ordenación que relaciona el número de meses húmedos y la temperatura media con los pisos altitudinales (Fig. 11). En este diagrama se nota la asimetría térmica, es decir, la tierra caliente sube en altura en la medida que aumenta la aridez, mismo fenómeno que se aprecia para la tierra subtemplada y templada. Pero a partir de la tierra fresca y hacia los pisos altitudinales superiores, la situación se vuelve más simétrica. Sobre el diagrama (Fig. 11) se ha superpuesto la distribución higro-térmica de la vegetación natural, que ciertamente se relaciona con los patrones de humedad y temperatura. Ello se ha realizado mediante observaciones en los fragmentos de vegetación remanentes, combinado con el tipo de epifitismo que domina en cada unidad de vegetación. La superposición de la información higro-térmica altitudinal con el mapa topográfico del área de estudio, permite generar la «carta de vegetación potencial» (Fig. 12).
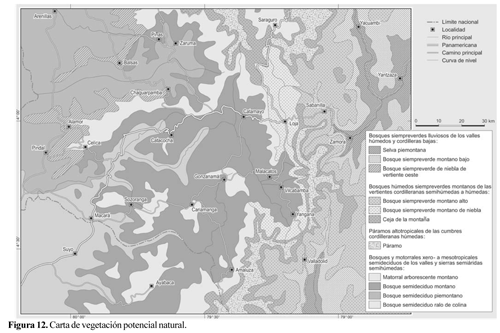
Carta de vegetación potencial
En estrecha relación con los patrones hídricos y térmicos analizados, es posible identificar y cartografiar la vegetación predominante, sobre la base de los fragmentos de vegetación remanente. La clasificación de formaciones vegetacionales se ha realizado tomando en cuenta anteriores trabajos, cartas y reportes para la zona de estudio (Grubb et al., 1963; Grubb y Whitmore, 1966; Kessler, 1991; Madsen y Øllgaard, 1993; Ulloa Ulloa y Møller Jørgensen, 1995; Valencia et al., 1999b; Balslev y Øllgaard, 2002; Bussmann, 2002; Lozano, 2002).
Se han diferenciado 11 formaciones vegetacionales naturales organizadas en cuatro tipos principales. Es muy probable que este número esté aún subestimado, pues no solamente la diferenciación florística y climática sino también la historia de la vegetación y diversos regímenes de disturbios son causantes de una extremada diversidad de formaciones y comunidades.
Tanto en la Figura 11 como en el mapa de vegetación (Fig. 12) se representan las siguientes formaciones vegetacionales:
· Bosques siempreverdes lluviosos (pluviotropicales e hidrotropicales, sensu Richter, 2001) de los valles húmedos y cordilleras bajas:
1. Selva piemontana (hasta 40 m de alto). 800-1300 m, alrededor de Yanzatza y Zamora en la vertiente oriental; tierra subtemplada, 10-12 meses húmedos. Flora característica: Beilschmiedia alloiophylla (Lauraceae), Billia columbiana (Hippocastanaceae), Caryodendron orinocense (Euphorbiaceae), Ceiba pentandra (Bombacaceae), Ceroxylon amazonicum, Iriartea deltoidea y Oenocarpus bataua (Arecaceae), Clarisia racemosa y Ficus subandina (Moraceae), Grias peruviana (Lecythidaceae), Guarea guidonia (Meliaceae), Leonia glycycarpa (Violaceae), Otoba glycycarpa (Myristicaceae), Terminalia amazonia y T. oblonga (Combretaceae).
2. Bosque siempreverde montano bajo (hasta 35 m de alto), 1200-2100 m, vertiente oriental de la Cordillera Real, e.g. alrededor de Sabanilla, tierra subtemplada y templada, 12 meses húmedos. Flora característica: Alchornea grandiflora (Euphorbiaceae), Alzatea verticillata (Lythraceae), Beilschmiedia sulcata (Lauraceae), Cedrela montana (Meliaceae), Clethra revoluta (Clethraceae), Clusia magnifolia y Vismia tomentosa (Clusiaceae), Cyathea caracasana (Cyatheaceae), Dictyocarpum lamarckianum (Arecaceae), Endlicheria sericea, Ocotea cernua y O. javitensis (Lauraceae), Guarea kunthiana (Meliaceae), Hedyosmum goudotianum (Chloranthaceae), Inga striata (Fabaceae), Nectandra acutifolia, N. membranacea, Persea caerulea y P. hexandra (Lauraceae), Piptocoma discolor (Asteraceae), Podocarpus sprucei (Podocarpaceae), Symplocos coriacea (Symplocaceae).
3. Bosque siempreverde de niebla de vertiente oeste (hasta 35 m de alto). 400-1600 m, vertiente occidental de la Cordillera de Celíca; tierra caliente y subtemplada, 7-11 meses húmedos (en el período seco con nieblas). Flora característica: Aiphanes grandis, Bactris setulosa, Geonoma cuneata, Iriartea deltoidea y Phytelephas aequatorialis (Arecaceae), Beilschmiedia alloiophylla (Lauraceae), Carapa guianensis (Meliaceae), Carludovica palmata (Cyclanthaceae), Castilla elastica y Ficus obtusifolia (Moraceae), Cedrela odorata (Meliaceae), Erythrina edulis (Fabaceae), Ruagea glabra (Meliaceae), Symplocos ecuadorensis (Symplocaceae) y Virola elongata (Myristicaceae).
· Bosques húmedos siempreverdes montanos (orotropicales) de las vertientes cordilleranas semihúmedas a húmedas.
4. Bosque siempreverde montano alto (hasta 25 m de alto), 2000-2700 m alrededor de Sabanilla en las vertientes y los valles orientales de la Cordillera Real, de Saraguro y de Amaluza; tierra fresca, 11 a 12 meses húmedos. Flora característica: Clethra revoluta (Clethraceae), Clusia elliptica y C. multiflora (Clusiaceae), Drimys granadensis (Winteraceae), Geissanthus vanderwerffii (Myrsinaceae), Geonoma densa (Arecaceae), Graffenrieda emarginata y Miconia rivetii (Melastomataceae), Heliocarpus americanus (Tiliaceae), Maytenus verticillata (Celastraceae), Myrica pubescens (Myricaceae), Myrsine andina (Myrsinaceae), Podocarpus oleifolius y Prumnopitys montana (Podocarpaceae), Purdiaea nutans (Cyrillaceae), Schefflera pentandra (Araliaceae) y Weinmannia pinnata (Cunoniaceae).
5. Bosque siempreverde montano de niebla (hasta 30 m de alto). 2500-2800 m sobre Yangana y Loja en las vertientes occidentales de la Cordillera Real; tierra templada y fresca, 8-10 meses húmedos. Flora característica: Cedrela montana (Meliaceae), Clusia alata y C. multiflora (Clusiaceae), Hedyosmum racemosum (Chloranthaceae), Ilex rupicola, (Aquifoliaceae), Myrcianthes rhopaloides (Myrtaceae), Myrica pubescens (Myricaceae), Ocotea rotundata y Persea ferruginea (Lauraceae), Prumnopitys montana (Podocarpaceae) y Ruagea hirsuta (Meliaceae).
6. Ceja de la montaña (hasta 8 m de alto). 2700-3100 m vertiente oriental y occidental de la Cordillera Real, de Amaluza y de Saraguro; tierra fría, 11-12 meses húmedos. Flora característica: Clethra ovalifolia (Clethraceae), Clusia elliptica (Clusiaceae), Disterigma acuminatum y D. pentandrum (Ericaceae), Gordonia fruticosa (Theaceae), Graffenrieda harlingii, Miconia ligustrina y M. theazans (Melastomataceae), Gynoxis cuicochensis (Asteraceae), Hedyosmum luteynii y H. cumbalense (Chloranthaceae), Hesperomeles ferruginea (Rosaceae), Oreopanax andreanus (Araliaceae), Weinmannia cochensis, W. elliptica, y W. fagaroides (Cunoniaceae), así como sitios de bambúes impenetrables compuestos de Chusquea falcata y Ch. scandens (Poaceae).
· Páramos altotropicales (en sentido florístico), jalcas (en sentido fisionómico) de las cumbres cordilleranas húmedas.
7. Páramo (tipo tolar alto hasta 1,5 m de alto, tipo ichu alto hasta 1 m). 3100-3700 m, cumbre de las Cordillera Real, de Amaluza y de Saraguro; tierra subhelada, 12 meses húmedos. Flora característica: Los páramos arbustivos poco intervenidos se componen de Baccharis genistelloides, Chuquiraga jussieui, Diplostephium empetrifolium y Hieracium frigidum (Asteraceae), Blechnum aurantium (Blechnaceae) Brachyotum andreanum (Melastomataceae), Escallonia myrtilloides (Grossulariaceae), Huperzia reflexa (Lycopodiaceae), Hypericum decandrum (Clusiaceae), Gaultheria amoena, Macleania rupestris, Pernettya prostrata y Vaccinium floribundum (Ericaceae), los bambúes Neurolepis elata, N. laegaardii y Chusquea neurophylla (Poaceae), Puya eryngioides, Puya nitida (Bromeliaceae), Rhynchospora vulcani (Cyperaceae) y Valeriana plantaginea (Valerianaceae). Sitios intervenidos mediante el fuego al norte, en la Cordillera de Saraguro, crean vastos y uniformes pastizales de Calamagrostis macrophylla, Stipa ichu y S. rosea (Poaceae).
· Bosques y matorrales semideciduos de los valles y sierras semiáridas a semihúmedas (xero- a mesotropicales).
8. Matorral arborescente montano (hasta 10 m de alto). 2000-2600 m, sierra interandina y vertiente occidental de la Cordillera Real; tierra templada y fresca, 6-8 meses húmedos. Flora característica: Alnus acuminata (Betulaceae), Baccharis alaternoides, B. latifolia y B. oblongifolia (Asteraceae), Cestrum tomentosum (Solanaceae), Hesperomeles heterophylla (Rosaceae), Lepechinia mutica (Lamiaceae), Oreocallis grandiflora y Roupala obovata (Proteaceae) y Tibouchina laxa (Melastomataceae). Formación intensamente transformada por actividades humanas, de forma que la reconstrucción de la vegetación original resulta de mucha dificultad. El efecto antrópico se refleja en la presencia de muchas especies características de bosque secundarios de sitios análogos en la vertiente oriental más húmeda.
9. Bosque semideciduo montano (hasta 15 m de alto). 1200-2300 m, sierra interandina y Cordillera de Celíca; tierra subtemplada y templada, 2-7 meses húmedos. Flora característica: Acacia macracantha (Fabaceae) y Tabebuia chrysantha (Bignoniaceae) en el estrato arbóreo y arbustos como Caesalpinia spinosa y Mimosa quitensis (Fabaceae), Ceiba insignis (Bombacaceae), Clethra fimbriata (Clethraceae), Croton wagneri (Euphorbiaceae), Heliotropium rufipilum (Boraginaceae), Jacaranda sparrei , Tecoma loxensis y T. sorbifoli (Bignoniaceae). Esta formación, intensamente transformada por el hombre, se encuentra ampliamente distribuida en la vertiente cordillerana occidental del departamento de Piura en Perú.
10. Bosque semideciduo piemontano (hasta 25 m de alto). 200-1000 m, vertiente occidental de la Cordillera de Celíca; tierra caliente, 4-6 meses húmedos. Flora característica: Attalea colenda y Phytelephas aequatorialis (Arecaceae), Castilla elastica (Moraceae), Cecropia litoralis (Cecropiaceae), Chrysophyllum argenteum y Pradosia montana (Sapotaceae), Cochlospermum vitifolium (Cochlospermaceae), Eriotheca discolor (Bombacaceae), Erythrina poeppigiana, Gliricidia brenningii, Pithecelobium excelsum y Syzygium jambos (Fabaceae), Gallesia integrifolia (Phytolacaceae), Ochroma lagopus (Bombacaceae), Sorocea sprucei (Moraceae) y Tabebuia chrysantha (Bignoniaceae).
11. Bosque semideciduo ralo de colina (hasta 15 m de de alto) 200-1200 m, Cuenca Catamayo y Chira; tierra caliente, 1-4 meses húmedos. Flora característica: Acacia huarango, A. macracantha, Albizia multiflora, Caesalpinia glabrata, Cercidium praecox, Geoffroea spinosa, Leucaena trichoides y Prosopis pallida (Fabaceae), Bursera graveolens (Burseraceae), Capparis scabrida y C. avicennifolia (Capparidaceae), Cavanillesia platanifolia, Ceiba trichistandra y Pseudobombax millei (Bombacaceae), Cordia lutea (Boraginaceae), Eriotheca ruizii (Bombacaceae), Loxopterygium huasango (Anacardiaceae) y Tabebuia bilbergii (Bignoniaceae), arbustos de Jatropha curcas (Euphorbiaceae) y los cactus en candelabro Armatocereus cartwrigthianus y Espostoa lanata (Cactaceae). Esta formación se encuentra ampliamente distribuida en las colinas piemontanas de la cordillera occidental del departamento de Piura en Perú.
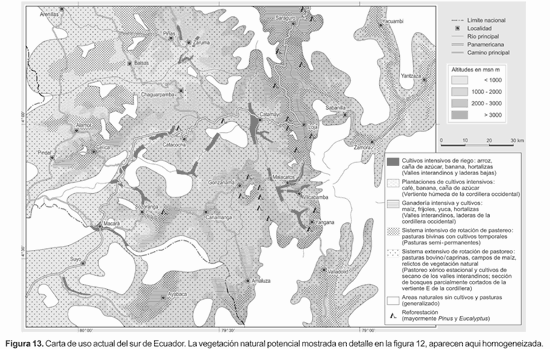
Los patrones de distribución de las principales formaciones de vegetación aclaran las relaciones ecológicas de la flora con el clima. Los resultados obtenidos en este caso para el sur de Ecuador constituyen una línea de base para el análisis del clima por medio de la fitoindicación epifítica. Sin embargo, la Figura 13 muestra el mapa de uso actual en el área, generado a partir de las observaciones en terreno. La vegetación natural remanente se encuentra muy fragmentada y la cobertura de utilización del suelo supera el 80%. Las principales actividades son cultivos intensivos de riego y de secano: arroz, café, caña de azúcar, banana, hortalizas, así como sistemas de rotación de pastoreo y cultivos extensivos: maíz, habas, yuca, hortalizas. Los sectores más intervenidos son los valles interandinos secos que han servido históricamente como áreas para el establecimiento humano. Los parches de vegetación natural más extensos se restringen evidentemente a los climas húmedos e inhóspitos de la cordillera oriental, incluido el Parque Nacional Podocarpus.
Conclusiones y discusión
Los bosques tropicales relictos de montaña entre las dos depresiones andinas en el sur de Ecuador y el norte del Perú (Girón-Cuenca y Huancabamba) presentan no sólo una enorme diversidad de especies sino también de formaciones vegetacionales. La reconstrucción de este complejo mosaico de formaciones vegetacionales naturales es extremadamente difícil, por tratarse de áreas muy intervenidas por el uso humano. El presente trabajo muestra un método adelantado de análisis bioclimático que permite realizar una cartografía de vegetación de detalle y de esta forma apoyar la toma de decisiones para la conservación en un centro de diversidad florística de importancia global. El hecho de existir este tipo de antecedentes para el sur de Ecuador constituye un desafío para su réplica mediante estas y similares técnicas en las cordilleras del norte de Perú, donde existe una topografía similarmente compleja con formaciones vegetacionales ricas en especies y necesitadas de protección.
Estudios biogeográficos en el norte de Perú demuestran la singularidad taxonómica de esta región, por ejemplo para familias como Loasaceae, Passifloraceae y Grossulariaceae (Weigend, 2002) y Solanaceae (Leiva, 2004). Los resultados demuestran que la zona de Amotape-Huancabamba, localizada entre el cauce del río Jubones en Ecuador y el cauce del río Chamaya en el Perú, constituye una zona fitogeográfica propia, como lo demuestra la cantidad de especies y grupos de especies endémicas de estrecho rango geográfico. La confluencia de elementos provenientes del norte como del sur, la diversidad de hábitat así como la historia geológica, parecen ser los factores más importantes que contribuyen a estos patrones de diversificación.
Gentry y Dodson (1987) proporcionan evidencias para la especiación asociada a un cambio genético rápido en géneros evolutivamente plásticos tales como Anthurium, Piper, y Cavendishia ("explosión evolutiva" sensu Gentry 1982). Dichos autores también precisan la diferenciación del micrositio que es típica para las cadenas andinas neotropicales, como catalizadores importantes de diversificación genética. Así, la zona de divergencia al sur del Nudo de Loja proporciona las mejores condiciones para la diversificación de especies, porque incluye una variedad amplia de hábitat húmedos y secos cercanos unos a otros debido al terreno extremadamente abrupto con laderas de diferente exposición. Además, el relieve cordillerano combinado con los regímenes de disturbios frecuentes y de diverso tamaño (roedores, fractura de copa, derrumbes, fuego, sequía) se pueden considerar situaciones óptimas para la especiación alopátrica, como consecuencia de la subdivisión microgeográfica del hábitat.
Largas distancias a lo largo de las crestas, la heterogeneidad de las áreas de la infiltración, la estructura topográfica con una alta fragmentación de diferentes mesoclimas y con diversos regímenes de disturbios, contribuyen a la extremadamente acelerada radiación adaptativa y riqueza de las comunidades especialmente en las regiones elevadas de la zona de Amotape-Huancabamba. Este hasta el momento aún poco investigado sector de los Andes debe ser reconocido como filogenéticamente muy importante, como lo demuestra el enorme endemismo en el sur de Ecuador y norte de Perú. Ello resulta de la fragmentación orográfica de la región, donde lograron desarrollarse ricas comunidades vegetales específicas en aislamiento (e.g. en los valles y cuencas secas entre Catamayo y Huancabamba, así como a lo largo del río Marañón). El alto número de taxa endémicos y el cambio genético rápido sugieren que en los valles y sierras existen hábitat poco saturados, receptivos frente a taxa plásticos de elementos florísticos hígricos y mésicos. Dichos elementos provienen de los sectores húmedos más altos a través de su migración latitudinal, y se establecen en los sectores más bajos en condiciones más secas. Es probable que en los sectores más secos se mezclen también elementos florísticos relictos que han evolucionado in situ en concordancia con el levantamiento andino (trazo árido interandino, según Ochsenius, 1999).
Para lograr comprender de mejor manera estos procesos evolutivos, así como para mostrar argumentos en favor de la conservación de la alta riqueza florística de las cordilleras del sur de Ecuador y norte del Perú, son ineludibles los conocimientos acerca del complejo clima regional. También en el norte del Perú se concentran las pocas estaciones meteorológicas en las menos complejas regiones costeras y de la Amazonía occidental. El método de fitoindicación mediante epífitas combinado con la evaluación de las temperaturas del suelo es especialmente útil para llenar el vacío de información climática en zonas montañosas, debido a su relativa facilidad de aplicación, así como los reducidos costos de tiempo y financiamiento.
Agradecimientos
El trabajo se desarrolló en el contexto del proyecto "Funcionalidad de un bosque tropical lluvioso montañoso del sur de Ecuador: el ecosistema con su biodiversidad, sus procesos dinámicos y el uso potencial del mismo" (<http://www.bergregenwald.de/>). El primer autor agradece a la DFG (Fundación Alemana de Investigación) por el generoso apoyo financiero. El hidrólogo Paul Emck (Erlangen) ha estado siempre disponible para una fructífera discusión acerca de la génesis climática en el sur de Ecuador. Eric Rodríguez (Trujillo) y Maximillian Weigend (Berlín) invitaron a M. Richter a participar en el muy informativo X Congreso Botánico Nacional en Trujillo en mayo de 2004.
Literatura citada
Balslev, H. y B. Øllgaard. 2002. Mapa de la vegetación del sur de Ecuador. En. Aguirre, Z., J.E. Madsen, E. Cotton y H. Balslev (eds.) Botánica Austroecuatoriana: Estudios sobre los recursos vegetales en las provincias de El Oro, Loja y Zamora. Ediciones Abya Yala, Quito, pp: 51-64. [ Links ]
Barthlott, W., N. Biedinger, G. Braun, F. Feig, G. Kier y J. Mutke.1999. Terminological and methodological aspects of the mapping and analysis of global biodiversity. Acta Botánica Fennica 162: 103-110.
Baumann, F. 1988. Geographische Verbreitung und Ökologie südamerikanischer Hochgebirgspflanzen. Beitrag zur Rekonstruktion der quartären Vegetationsgeschichte der Anden. Physische Geographie 28, Zürich.
Bussmann, R.W. 2002. Estudio fitosociológico de la vegetación en la Reserva Biológica San Francisco (ECSF) Zamora-Chinchipe. Herbario Loja 8. Loja.
Churchill, S.P., H. Balslev, E. Forero y J.L. Luteyn (eds).1995. Biodiversity and Conservation of Neotropical Montane Forests. Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium, The New York Botanical Garden, 21-26, June 1993. 702 p.
Gentry, A.H. 1982. Neotropical floristic diversity: phytogeographical connections between Central and South America, Pleistocene climatic fluctuations, or an accident of the Andean orogeny? Ann. Missouri Botanical Garden 69, 557-593.
Gentry, A.H. y C.H. Dodson. 1987. Diversity and biogeography of neotropical vascular epiphytes. Ann. Missouri Botanical Garden 74, 205-233.
Grubb, P.J., J.R. Lloyd, T.D. Pennington y T.C. Withmore. 1963. A comparison of montane and lowland rain forest in Ecuador. I. The structure, physiognomy, and floristics. J. Ecol. 51, 567-601.
Grubb, P.J. y T.C. Withmore. 1966. A comparison of montane and lowland rain forest in Ecuador. II. The climate and its effects on the distribution and physiognomy of the forests. J. Ecol. 54, 303-333.
Kessler, M. 1992. The vegetation of south-west Ecuador. In: B.J. Best, ed. The threatened forests of south-west Ecuador. Leeds, U.K.: Biosphere Publications, 240 p. pp. 79-100.
Lauer, W. 1995. Klimatologie. Braunschweig.
Leiva, S. 2004. Avances en la sistemática de las Solanáceas del Perú. En Libro de Resúmenes del X Congreso Nacional de Botánica, Trujillo, Perú. p. 46.
Lozano, P.E. 2002. Los tipos de bosques en el sur de Ecuador. En: Aguirre, Z., J.E. Madsen, E. Cotton y H. Balslev (eds.) Botánica Austroecuatoriana: Estudios sobre los recursos vegetales en las provincias de El Oro, Loja y Zamora. Quito, pp. 29-49.
Madsen, J.E. y B. Øllgaard. 1993. Inventario preliminar de las especies vegetales en el Parque Nacional Podocarpus. Ciencias agrícolas 22-23, UN Loja, 66-87.
Myers N, R.A. Mittermeier, C.G. Mittermeier, G.A.B. DA Fonseca y J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
Ochsenius, C. 1999. The Neotropical Aridity. Geowissenschaftliche Beiträge, Carl-Christian Ochsenius-Stiftung, Baden-Württemberg & Santiago.
Richter, M. 1996. Klimatologische und pflanzen-morphologische Vertikalgradienten in Hochgebirgen. Erdkunde 50, 205-237.
Richter, M. 2001. Vegetationszonen der Erde. Gotha y Stuttgart.
Richter, M. 2003. Using epiphytes and soil temperatures for eco-climatic interpretations in Southern Ecuador. Erdkunde 57: 161-181.
Ulloa Ulloa, C. y P. Møller Jørgensen. 1995. Árboles y arbustos de los Andes del Ecuador. AAU Reports 30, Aarhus.
Valencia, R., C. Cerón, W. Palacios y R. Sierra. 1999a. Los sistemas de clasificación de la vegetación propuestos para el Ecuador. En: Sierra, R. (ed). Propuesta preliminar de un sistema de clasificación de vegetación para el Ecuador continental. Proyecto INEFAN/GEF-BIRF y EcoCiencia,Quito, pp. 19-28.
Valencia, R., C. Cerón, W. Palacios y R. Sierra. 1999b. Las formaciones naturales de la sierra del Ecuador. En: Sierra, R. (ed). Propuesta preliminar de un sistema de clasificación de vegetación para el Ecuador continental. Proyecto INEFAN/GEF-BIRF y EcoCiencia, Quito, pp. 79-108.
Winiger, M. 1981. Zur thermischen Gliederung des Mount Kenya. Erdkunde 35, 121-150.
Weigend, M. 2002. Observations on the Biogeography of the Amotape-Huancabamba Zone in Northern Peru. The Botanical Review 68 (1): 38-54.
Weischet, W. 1996. Regionale Klimatologie. Teil 1, die Neue Welt. Stuttgart
Young, K.R. y C. Reynel. 1997. Huancabamba Region, Peru and Ecuador. En: Davis, S.D., V.H. Heywood, O. Herrera-MacBryde, J. Villa-Lobos y A.C. Hamilton. Centres of Plant Diversity: A Guide and Strategy for their Conservation,vol.3, The Americas: 465-469.
Correspondencia
*Institut für Geographie, Friedrich Alexander Universität - Erlangen, Alemania
E-mail:Michael Richter mrichter@geographie.uni-erlangen.de
