










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.12 n.2 Lima ago./set 2005
Maximiliam Weigend1, Asunción Cano2, Eric F. Rodríguez3
1 Institut für Biologie Systematische Botanik und Pflanzengeographie, Freie Universität Berlin, GERMANY
2 Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, Lima, PERÚ.
3 Herbarium Truxillense (HUT), Universidad Nacional de Trujillo, Trujillo, PERÚ.
Número especial: Bosques relictos del NO de Perú y SO de Ecuador
Weigend, Rodríguez y Arana (Comps.)
Publicado online: 08/12/05
Abstract
The taxonomy of the Ribes andicola group and Urtica is investigated for the relict forests in the Amotape-Huancabamba Zone and especially the relict forests of NW Peru. Three new species of Ribes are described, Ribes contumazensis as local endemic from Contumazá, R. colandina as widespread across the Andes from Lambayeque and La Libertad to Amazonas and R. sanchezii as endemic to forests overlooking the Marañon. Additionally, Ecuadoren R. austroecuadorense is recorded for the first time from Peru from forest relics in Huancabamba (Piura) and Chota (Cajamarca). Two new species of Urtica are described, U. urentivelutina as a local endemic on the slopes overlooking the Marañon near Chagual (La Libertad), and U. lalibertadensis which is widespread in La Libertad from Otuzco to Tayabamba. Urtica longispica is reported as new to the flora of Peru and U. macbridei is reported from northern Peru for the first time. Both Urtica and the Ribes andicola group can be considered as indicators of former forest cover, since they tenaciously persist after the felling of the forests. Their distribution indicates that a) some of the current forest fragments were once largely coherent forest belts (e.g., those of Santa Cruz and San Miguel) and b) some areas which have no coherent forest cover at present once had large and coherent montane forests (e.g., large parts of the Prov. Otuzco). Comparing the known distribution data for Urtica, the Ribes andicola group, the Passiflora lobbii group and Nasa (Loasaceae) it becomes evident, that a) the relict forests are still very poorly known, b) they are exceptionally rich in narrowly endemic taxa, some of which are here described and many of which undoubtedly remain to be discovered and c) they represent southernmost outposts for many Ecuadorean species and species groups.
Key words: Urtica, Ribes, relict forests, biodiversity, Amotape-Huancabamba zone.
Resumen
Se investiga la taxonomía del grupo de Ribes andicola y Urtica para los bosques relictos presentes en la zona de Amotape-Huancabamba y especialmente aquellos de la vertiente NO del Perú. Se describen tres nuevas especies de Ribes: Ribes contumazensis como especie local endémica de Contumazá, R. colandina distribuida en los Andes desde Lambayeque y La Libertad hasta Amazonas y R. sanchezii como endémica a los bosques de altura del Marañón. Adicionalmente, la especie ecuatoriana R. austroecuadorense es registrada por primera vez para el Perú procedente de los bosques relictos de Huancabamba (Piura) y Chota (Cajamarca). Así mismo, se describen dos nuevas especies de Urtica: U. urentivelutina, endémica local a las laderas altas del Marañón cerca a Chagual (La Libertad), y U. lalibertadensis distribuida desde Otuzco hasta Tayabamba en el departamento de La Libertad. Urtica longispica es registrada como nueva especie para la flora del Peru y U. macbridei es registrada por primera vez para el norte del Perú. Urtica y el grupo Ribes andicola pueden ser considerados como indicadores de la cubierta de bosques anteriores, puesto que persisten después que los bosques han sido talados. Su distribución indica que a) alguno de los actuales fragmentos de bosque fueron alguna vez grandes bosques coherentes a modo de una faja contínua (como los de Santa Cruz y San Miguel) y b) algunas áreas que en el presente no tienen ninguna cubierta de bosques contínuos, en el pasado fueron grandes y coherentes bosques montanos (e.g., grandes partes de la Prov. Otuzco). Comparando los datos de distribución conocida para Urtica, el grupo de Ribes andicola, el grupo de Passiflora lobbii y Nasa (Loasaceae), es evidente, que a) los bosques relictos son aún muy pobremente estudiados, b) ellos son excepcionalmente ricos en taxa estrechamente endémicos, algunos de los cuales son aquí descritos y muchos de ellos indudablemente permanecen aún sin ser descubiertos y c) ellos representan los límites más sureños para muchas especies ecuatorianas y ciertos grupos de especies.
Palabras claves: Urtica, Ribes, bosques relictos, biodiversidad, zona de Amotape-Huancabamba.
Introduction
The existence of relict forests in NW Peru has been known for a long time and their ecology is discussed at length in, e.g., Koepcke (1961). The most well-known forest fragments from a botanical point of view are found in the Department of Cajamarca (Bosque de Cutervo, Bosque de Monte Seco, Bosque de Cachil), but there are actually quite a large number of forest fragments also in Piura, Lambayeque and La Libertad. These relict forests are largely found on the western slope of the Andes and in the upper reaches of the interandean valleys. The northern parts of the Department Cajamarca (Provinces Jaén and Cajamarca) also have some interandean forests which are more or less continuous with the eastern slopes of the Andes (Depto. Amazonas).
Recent years have led to the description of numerous new and usually narrowly endemic taxa of various plant families from these forest fragments. However, there are still many plant groups which are very incompletely known in these relict forests, including some abundant and widespread elements. The present article wants to resolve some taxonomic questions in Ribes (Grossulariaceae) and Urtica (Urticaceae), both of which are typical and often abundant elements of the cloud forest vegetation and - unlike many other plant groups persist for a long time in former cloud forest areas, which have been cleared for agriculture. Both Urtica and Ribes are then found in hedges and along dry stone walls. Since both taxa are present throughout the year and often dominant or subdominant in the vegetation we would expect them to be very well documented in herbaria, i.e., we would think that their documentation would be a good indicator of how much we actually know about the flora of the relict forests. Conversely, a forest from which neither of the genera is documented can be considered as botanically virtually unknown. Both genera have not been taxonomically revised for a very long time the last monograph including South American Ribes is nearly 100 years old (Janczewski, 1907) and the last comprehensive treatment of Urtica is over 135 years old (Weddell, 1869). The Flora of Peru (1941) treatments for both taxa are best completely ignored.
Additionally, both genera have numerous widespread species and are thus useful for looking for phytogeographic relationships. The data are presented in the context of the phytogeography of the Amotape-Huancabamba Zone, so that distribution data will be given for all relevant taxa from the region. The following questions will be adressed:
- Which taxa of the respective groups are found in the relict forests of NW Peru?
- What are the phytogeographical affinities of the taxa associated with the relict forests?
- Are they mostly narrowly endemic, or are they mostly wide-ranging taxa?
- From which of the known forest fragments are Urtica and/or Ribes documented?

Materials and methods
Field studies were carried out in all Andean departments of Peru between 1997 and 2004. Additionally, herbarium material was revised in all major international herbaria (Ribes) or a selection of European herbaria and the local Peruvian herbaria (Urtica). All protologues and, where necessary, type elements were studied for identification.
Results
Urtica (Urticaceae)
Urtica in northern Peru
The genus Urtica is present with 6 native species in Peru (according to Brako & Zarucchi, 1993), since then one new species has been described from the relict forests in NW Peru (Prov. Contumazá, Depto. Cajamarca; Geltman, 1998). The Peruvian Urtica can be tentatively subdivided into two ecological groups, a high-Andean group of relatively small annual or perennial herbs (<<0.5 m, U. echinata, U. trichantha, U. flabellata) from paramo and puna habitats and a second group from intermediate elevations which is found in cloud forest and in places where cloud forest has been replaced by secondary vegetation due to anthropic activity. The latter taxa are all perennial, often suffrutescent or lianescent with at least basally woody shoots, have at least basally united stipules and are much larger (>1 m, sometimes >5 m) and include the taxa U. macbridei, U. ballotaefolia, U. peruviana, U. longispica. These species of Urtica are typically found along rivulets in forest, along the forest margins, on (natural and man-made) landslides and generally disturbed sites. This species group is widespread from Costa Rica to Patagonia and would be expected in all mesic forest remnants of NW Peru. However, the current literature gives very few data on the presence and taxonomy of these Urtica-species in the relict forests, and this article aims at giving some first data to close that gap. The crucial problem here is, however, that the systematics of Urtica in the Andes is very poorly resolved at present and both the names and the distribution data are highly ambiguous.
In reporting U. magellanica from Peru and the relict forests in the north Brako & Zarucchi (1993) follow Macbride (1941), which in turn gets his species definition from Weddell (1858, 1869). The Bolivian material previously attributed to U. magellanica has since been removed to a new species (U. pseudomagellanica, Geltman, 1998), but no new treatment has been proposed for the Peruvian material, although it definitly does not belong into U. magellanica, a south temperate species restricted to Argentina and Chile.
For the relict forests in NW Peru and for the Amotape-Huancabamba Zone there are few if any citations of Urtica and Brako & Zarucchi (1993) list not a single species/specimen for the departments of Piura, Lambayeque, Amazonas and La Libertad which is implausible given the universal presence of Urtica in Andean habitats. Recent field studies and herbarium studies yielded material of various species of Urtica from the relevant area and taxonomical interpretations are here offered for these.
Urtica macbridei was described from Huánuco and is apparently widespread on the E slope of the Andes, but absent from the western slope including the N Peruvian relict forests, where it is replaced by the very closely related U. longispica, which is widespread in Colombia and Ecuador and only known from the forests of Ayavaca (Depto. Piura) and San Miguel (Depto. Cajamarca), i.e., has a few isolated SW outposts in the relict forests of NW Peru. It is likely present also in most other relict forests, but I have not yet seen any specimens from there. Urtica longispica is replaced by U. peruviana south of the Río Jequetepeque in the Prov. Contumazá, a recently described species which is easily differentiated by the possession of strongly ramified (vs. unbranched) inflorescences. South of Contumazá both U. peruviana and U. longispica are absent and only widespread U. leptophylla is found. Other forest fragments in the Amotape-Huancabamba zone have so far not been systematically investigated with regards to their Urticas, but a collection trip in 2004 brought two new endemic species: A densely pubescent liana apparently restricted to the residual montane cloud forests on the east-facing slopes of the Marañon valley is described as Urtica urentivelutina and a very widespread species from areas with former forest cover and some relict forests in Otuzco is described as Urtica lalibertadensis. Thus the relict forests in the Amotape-Huancabamba Zone are home to two morphologically well-differentiated and very narrowly endemic species of Urtica, one in the relict forests on the Pacific slope and one in the relict forests in an interandean valley. These are the only narrowly endemic species of Urtica between Mexico and Central Chile.
Key to the rhizomatous species of Urtica with at least basally fused stipules in northern Peru
1. Inflorescences unisexual, male inflorescences usually much longer than the subtending leaves.
2. Plants finely and shortly pubescent with simple trichomes typically <0.5 mm long; stipules subglabrous, with scattered trichomes along the margin; perigon of female flowers without stinging hairs.
Urtica macbridei
2. Plants densely and long pubescent with simple trichomes typically 1 mm long; stipules completely pubescent; perigon of female flowers with stinging hairs.
Urtica urentivelutina
1*. Inflorescences androgynous, if lower ones male, then much shorter than the subtending leaves.
3. Inflorescences completely unbranched and continuously set with sessile glomerules, distinctly catkin-like, interspersed with small, hyaline bracts.
Urtica longispica
3*. Inflorescences at least marginally, with some glomerules over 5 mm, distinctly irregular and interrupted, without hyaline bracts.
4 Stem only with deflexed setae, otherwise glabrous; perigon (male and female) with stinging hairs.
Urtica lalibertadensis
4*. Stem with scattered, erect setae and additionally with numerous, usually appressed white trichomes; perigon (male and female) without stinging hairs.
5. Inflorescences laxly branched, with at least some branches of mature inflorescences 1015 mm long; lamina margin with 2030 teeth on each side.
Urtica peruviana
5*. Inflorescences densely branched, with at least branches of mature inflorescences ca. 5 mm long; lamina margin with <20 teeth on each side.
Urtica leptophylla
Urtica - formal taxonomy
Urtica macbridei Killip.
Journ. Wash. Acad. Sci 15: 49. 1925. - Type: Peru, Depto. Huánuco, Muña, 2400 m, Macbride 4271 (F, neg. nr. 69600!, US!).
Lianescent shrub 1,53 (4?) m, rarely erect subshrub from lignescent rhizome ca. 35 mm thick, stems erect or lax, with scattered stinging haris and densely covered with simple, often appressed trichomes ca. 0,20,4 mm long. Leaves opposite, interpetiolar stipules united in pairs but deeply incised for ca. 50% of their length, subcircular, ca. 58 mm long and wide, with conspicuous white cistoliths and with scattered, white simple trichomes ca. 0.50,8 mm along the margin only; petioles 46 (8) cm long, widely ovate to cordate, 10 x 8,5 12 x 9 cm, apex acuminate, base deeply cordate (sinus to 15 mm deep), margin coarsely simply serrate with 2030 teeth on each side, both surfaces finely pubescent and covered with scattered short stinging hairs (ca. 1 mm), cistoliths largely punctiform. Inflorescences unisexual, rarely a few male flowers in the proximal portion of female inflorescences, arising in pairs from the leaf axils, male inflorescences up to 15 cm long, female inflorescences up to 4 cm long, flowers in dense, subsessile ebracteose glomerules, female flowers ca. 1 mm long (including plumose stigma) with finely pubescent, esetulose perigon, male flowers with perigon ca. 1 mm long; fruit ovate, ca. 11,25 x 0,751 mm with very short apex, largely included in finely pubescent, esetulose perigon.
Specimens seen: Ecuador, Prov. unknown, Guataxi, Spruce 6536 (BM). Peru, [Depto. Huánuco, Muña?] Ruiz & Pavón s.n. (B!). Depto. Amazonas, Prov. Chachapoyas, Leimebamba, Laguna de los Condores, Pampa, 26003500 m, 15.05.2004, Schwarzer 05 (B, USM).
This species seems to be very widespread along the eastern slope of the Andes in Peru and Ecuador, but fails to penetrate into inner-andean valleys, where it is replaced by other species. The following species, U. urentivelutina, is apparently its closest relative, but is readily differentiated on the basis of its much denser indument and replaces U. macbridei in the seasonally dry forest fragments W of the Marañón.
Urtica urentivelutina Weigend, spec. nov.
Holotype: Peru, Depto. La Libertad, Prov. Sánchez Carrión, road Huamachuco-Chagual-Pataz, after Chugay and before reaching Aricapampa, in the village Molino Viejo, 3048 m, 20.04.2004, Weigend & Schwarzer 7907 (male, USM, iso: B, HUT).
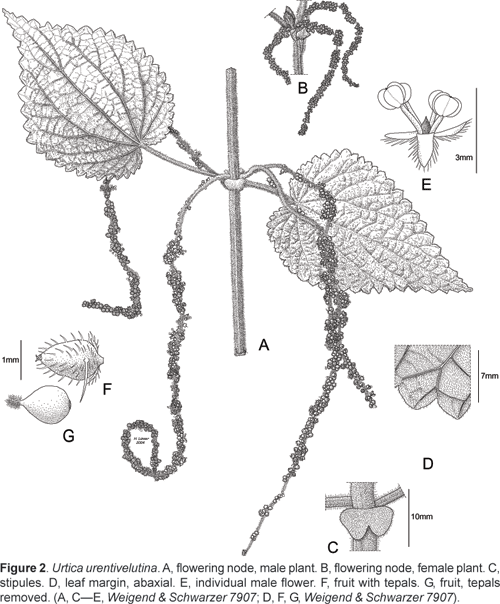
Urtica frutescens, scandens, perennis, monoica. Caulis robustus, stimulosus et densissime pilosus; paniculae unisexuales, masculi petiolo multo longiori, perigonium laciniis pilosis.
Liana 35 (6?) m tall from ligneous rhizome ca. 810 mm thick, stems lax, with scattered stinging hairs 1,5—2 mm long and very dense, white cover of simple trichomes 11,5 mm long. Leaves opposite, interpetiolar stipules united in pairs but deeply incised for ca. 3040% of their length, subcircular, ca. 810 mm long and wide, completely covered with white simple trichomes ca. 0,51 mm; petioles 24 (8) cm long, lamina widely ovate, 9 x 6 15 x 8 cm, apex acuminate, base cordate (sinus ca. 8 mm deep), margin coarsely simply serrate with 1825 teeth on each side, lamina strongly bullate, both surfaces with scattered stinging hairs ca. 1,5 mm long and completely covered with white, simple trichomes ca. 11,5 mm long, cistoliths largely punctiform. Inflorescences unisexual, male inflorescences ca. 1215 (18) cm long during anthesis, female inflorescences ca. 36 cm long, arising in pairs from the leaf axils, flowers in dense, subsessile glomerules, female flowers ca. 1 mm long (including plumose stigma), perigon lobes densely pubescent, with 13 (5) stinging hairs on each of the larger perigon lobes, male flowers with perigon ca. 1,5 mm long, perigon lobes densely pubescent, filaments ca. 3 mm long, anthers reniform, ca 1 mm in diameter; fruit ovate, ca. 1,5 x 1 mm with very short apex, largely included in pubescent perigon.
Specimens seen: Peru, Depto. La Libertad, Prov. Sánchez Carrión, road Huamachuco-Chagual-Pataz, after Chugay and before reaching Aricapampa, in the village Molino Viejo, 3048 m, 20.04.2004, Weigend & Schwarzer 7906 (female, USM, B, HUT).
This new species is closely allied to U. macbridei from the eastern slope of the Peruvian Andes, but clearly differs in its much denser and longer (11,5 vs. ca. 0,3 mm) indument, especially on the stipules (subglabrous in U. macbridei) and the presence of stinging hairs on the perigon of the female flowers. The leaves are very densely pubescent and also irrgularly bullate between the veins, which is a character not known from the other Peruvian species here discussed. The taxon is so far only known from a single collection near Molino Viejo just W of the Marañon rives. It is found in the remnants of cloud forest vegetation, which has here been largely destroyed. In indument it closely resembles Nasa urentivelutina, which is found in a perfectly analogous geographical position between Balsas and Celendín somewhat further North (Cajamarca, Prov. Celendín) and also is an atypically densely pubescent and stinging species closely allied to less pubescent species E of the Marañon, i.e., shows the same differentiation from its congeners as U. urentivelutina.
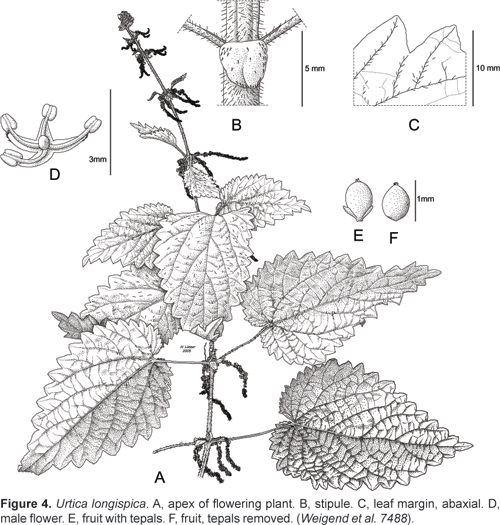
Urtica longispica Killip.
J. Wash. Acad. Sci. 19(10): 191. 1929. a» Urtica ballotaefolia Wedd. var. macrostachya Wedd., Monogr. Urticaceae: 74. 1856. Syntypes: Colombia, Prov. Cundinamarca, near Bogotá, San Antonio, Boqueron de la Mesa, Goudot 4 (P!), without precise locality, Triana 163 (P?, BM!). Lectotype: Colombia, Prov. Cundinamarca, near Bogotá, San Antonio, Boqueron de la Mesa, Goudot 4 (P!).
Erect subshrub 0,81,5 (2?) m, more rarely lianescent, from lignescent rhizome ca. 35 mm thick, stems erect or lax, with scattered stinging hairs ca 1,5 mm long and very short, appressed, simple trichomes ca. 0,1 mm long. Leaves opposite, interpetiolar stipules united in pairs but deeply incised for ca. 50% of their length, subcircular, ca. 36 mm long and wide, with conspicuous white cistoliths and with scattered, white simple trichomes ca. 0,10,3 mm along the margin only; petioles 24 (5) cm long, widely ovate to triangular-ovate, 10 x 5 12 x 6 cm, apex acuminate, base truncate to subcordate (sinus to 7 mm deep), margin coarsely simply, rarely doubly serrate with 1520 teeth on each side, both surfaces very finely pubescent to subglabrous and covered with scattered short stinging hairs (ca. 12,5 mm), cistoliths largely punctiform. Inflorescences androgynous, ca. 3040 mm long during anthesis, elongating to >12 cm in fruit, with male flowers in the proximal portion (510 mm) and female flowers in the distal (1025 mm) portion of inflorescences, arising in pairs from the leaf axils, flowers in dense, subsessile glomerules with tiny (to 1 mm), often at last marginally hyaline bracts, female flowers ca. 1 mm long (including plumose stigma), perigon lobes finely pubescent, without stinging hairs, male flowers with perigon ca. 1 mm long, perigon lobes finely pubescent; fruit ovate, ca. 11,25 x 0,751 mm with very short apex, largely included in pubescent perigon.
Specimens seen: Colombia, Depto. Caldas, above Salento, path from Romerales to Salneto, Amarguras, 3200 m, 29.09.1948, Hawkes 417 (BM). Ecuador, Prov. Pichincha, old road Chillagallo-San Juan Chiriboga Empalme, km 27, 00º17´ S, 78º38´ W, 3180 m, 23.02.1986, Zak 970 (GB, MO). Prov. Napo-Pastaza, ENE Cayambe, 3600 m, Cazalet & Pennington 5555 (B, NY). Prov. Bolívar; Parroquía Simiatug, Guaranda, 3200 m, 28.02.1982, Bravo-Falconi 331 (GB). Prov. Loja, Celica-Zapotillo road, km 5, 21002200 m, 23.02.1985, Harling & Andersson 22444 (GB). Peru, Depto Piura, Prov. Huancabamba, near Huancabamba, 19502150 m, 18.07.1994, Yarupaitán 1405 (B, USM). Depto. Cajamarca, Provincia Cajamarca, El Molino (San Pablo), 2340 m, «ishguin», Sagástegui et al. 7993 (HUT, MO). Prov. San Miguel, San Miguel to Agua Blanca, 07º02´ 51´´ S, 78º59´ 17´´ W, 2762 m, 01.05.2003, Weigend et al. 7485 (B, USM, M, HUT). Alrededores de El Tingo (Agua Blanca), 2950 m, 5.7.1986, «ishguin», Mostacero et al. 1323 (HUT, F, MO). Road San Miguel to Tongod (Santa Rosa-Hualgayoc), S 06º58´ 03,4´´, W 078º50´10,3´´, 2917 m, 02.05.2003, M. Weigend et al. 7488 (B, HUT, USM).
Urtica longispica was explicitly created as nomen novum for U. ballotaefolia var. macrostachya Wedd. by Killip, and thus is automatically homotypical. Both Geltman (1998) and Jørgensen and León (1999) consider U. longispica as a distinct species from U. ballotaefolia and the voucher cited Jørgensen and León (1999: Cazalet & Pennington 5555) is indeed clearly distinct and agrees perfectly with one type specimen in Paris (Goudot 4), which is here selected as the lectotype. The isosyntype at British Museum (Triana 163), however, is clearly identical to U. leptophylla (=U. ballotaefolia).
The taxon is characterized by its unbranched, androgynous inflorescences and very dense, unbranched infructescences. The flowers are interspersed by tiny, hyaline bracts not found in any of the other species. The specimen Harling & Andersson 22444 is at first glance very aberrant, but probably belongs here: fruiting material from that population would be highly desirable. This species ranges from Colombia to Ecuador and has its southernmost populations in the relict forests of NW Peru. Apparently the southern limit of the species is marked by the Jequetepeque-valley, since it is abundant in the Prov. San Miguel, but replaced by U. peruviana just south of that valley in Contumazá.

Urtica leptophylla Kunth in Humb.
Bonpl. & Kunth, Nov. Gen. Sp. Pl. 2: 39. 1815. Types: [Colombia], Nova Granata, [Cauca]: Almaguer, Meneses, Humboldt & Bonpland s.n. (P-Bonpl., photo Field # 39594!), Bonpland 2438 (P!, 2 specimens). Lectotype (here designated): [Colombia], Nova Granata, [Cauca]: Almaguer, Meneses, Humboldt & Bonpland s.n. (P-Bonpl., photo Field # 39594!).
= Urtica ballotaefolia Wedd., Ann. Sci. Nat. Ser. 3, 18: 197. 1852. Type: [Colombia, Cundinamarca] near Bogotá, August 1844, Goudot 3 (P! Field Museum neg. nr. 40272!).
Erect perennial herb 0,61,5 (2) m tall, basally not or slightly lignified, from short, lignescent rhizome ca. 34 mm thick, stems stiffly erect, with numerous stinging hairs ca 11,5 mm long and very short, appressed, simple trichomes ca. 0,10,2 mm long. Leaves opposite, interpetiolar stipules united in pairs but deeply incised for ca. 5070% of their length, subcircular, ca. 48 mm long and wide, without conspicuous white cistoliths and with scattered, white simple trichomes ca. 0,10,3 mm over the entire surface; petioles (2,5)46 (8) cm long, lamina widely ovate, (6 x 3) 8 x 5 10 x 7 cm, apex acuminate, base truncate to subcordate (sinus to 6 mm deep), margin coarsely simply, rarely doubly serrate with 1520 teeth on each side, both surfaces very finely pubescent to subglabrous and covered with scattered short stinging hairs (ca. 12,5 mm), cistoliths largely punctiform. Inflorescences androgynous, lowest ones often pure male, upper one often pure female, ca. (25) 3060 mm long during anthesis, elongating to >12 cm in fruit, typically with male flowers in the proximal half and female flowers in the distal half, arising in pairs from the leaf axils, inflorescences irregularly interrupted, with flowers in ebracteose glomerules up to 7 mm long, female flowers ca. 1 mm long (including plumose stigma), perigon lobes finely pubescent, without stinging hairs, male flowers with perigon ca. 1 mm long, perigon lobes finely pubescent; fruit ovate, ca. 1,25 x 1 mm with very short apex, largely included in pubescent perigon.
Specimens seen: Colombia, Depto. Cundinamarca, Chiquinquirá, 2500 m, 17.02.1950, von Sneijdern 5823 (BM). Depto. Cundinamarca, Bogotá, 2660 m, anno 1854, Triana 874 (BM). Depto Cauca, Páramo de Ruiz, 22003000 m, Sept. 1883, Lehmann 3159 (BM). - Ecuador, Prov. Riobamba, Riobamba, 3000 m, Dez. 1932, Rimbach s.n. (B). Prov. Cañar, Ingapirca, 3200 m, 02º33´S, 78º52´ W, 3200 m, Løjtnant et al. 14871 (GB). Peru, Depto Lima, Prov. Yauyos, Cruz Pampa, 2950 m, 07.01.1952, Cerrate 1112 (B, USM). Depto Lima, Prov. Huarochiri, Huaquicha, above Surco, 2900 m, 09.03.1974, Cerrate 5811 (B, USM). Near Santiago, 3600 m, 16.05.1953, Cerrate 1926 (B, USM). Depto Lima, Cruz Pampa above Supe, 2950 m, 07.01.1952, Cerrate 1112 (B, USM). - Depto. Ancash, Prov. Carhuaz, without precise locality or elevation, 10.03.1964, Wright 4308 (B, USM). Depto. Ancash, Prov. Huaraz, Río Grande/Río Chacchan, towards Pariacoto, 18L 0200571 UTM 8942645, 3000 m, 16.05.2003, Weigend et al. 7698 (B, M, USM). Prov. Huaylas, between Pamparomas and Laguna Negra Huacanan, just before Karka, 12.10.2002, Weigend et al. 7407 (B, HUT, M, USM). Just above Pamparomas on road to Caraz, 18L 0173333 UTM 8994296, 3133 m, 14.05.2003, Weigend et al. 7682 (B, M, USM). Between Pamparomas and Pampap, 09º03´S, 77º58´ W, 2850 m, 26.09.2001, Weigend & Skrabal 5895 (B, HUT, M, USM).
The descriptions, the material I have seen and cultivated, and the type specimens (photographs) I have revised all agree with the judgement, that Geltmann (1998) gives in passing: The taxa U. ballotaefolia and U. leptophylla are indistinguishable. Weddell (1858, 1869) and Macbride (1941) consider U. leptophylla as a synonym of U. magellanica, which, however, is not present in Colombia (nor anywhere north of Chile), whereas W.C. Burger (Fieldiana 40. 1977) first recongized that U. leptophylla is a valid taxon and that both U. nicaraguensis and U. copeyana (from Costa Rica) are synonymous with that taxon. The Central American material differs superficially from the Andean material in shorter inflorescences and smaller leaves, but Burger´s judgement will probably turn out to be correct once more material is revised. The Peruvian material is very clearly conspecific with U. leptophylla/U. ballotaefolia. U. leptophylla will remain the oldest name for the taxon even if it should turn out that one of the Chilean species is conspecific, since the relevant names are all younger (e.g., U. pseudodioica, U. buchtienii, Taylor, 2003).
The name Urtica magellanica is often erroneously employed for introduced, weedy U. urens from Europe, but also for at least three different native species, most commonly for U. leptophylla, the most widespread and abundant Urtica species in Peru (Ancash to Arequipa). Of all Central and Northern Andean taxa U. leptophylla is most similar to the north-temperate U. dioica-complex and many reports of (presumably introduced) U. dioica from South America probably refer to this taxon. It is particularly close to variable (monoecious) U. dioica subsp. gracilis from North America (Woodland, 1982) and future studies will be required to establish taxon limits.
Urtica leptophylla is present both north and south of the Amotape-Huancabamba Zone, but is apparently absent from the Amotape-Huancabamba Zone and its relict forests, where it is replaced by endemic taxa such as U. urentivelutina, U. lalibertadensis and U. peruviana.
Urtica lalibertadensis Weigend, spec. nov.
Holotype: Peru, Depto. La Libertad, Prov. Pataz, between Huancaspata and Tayabamba, S 08º18,053´, W 077º17,032´, 3071 m, 24.04.2004, Weigend & Schwarzer 8018 (USM, iso: B, HUT).
Urtica herbacea, perennis, monoica. Caulis robustus, stimulosus, glabrescens; paniculae androgynae, petiolo multo longiorae, perigonium laciniis valde hispidis.
Lianescent subshrub or erect perennial herb 0,61,5 (3) m tall, basally slightly lignified, from lignescent rhizome ca. 35 mm thick, stems erect, with numerous deflexed stinging hairs ca 1.52 mm long, otherwise glabrous. Leaves opposite, interpetiolar stipules united in pairs but deeply incised for ca. 5070% of their length, subcircular, ca. 610 mm long and wide, without conspicuous cistoliths and with scattered, white simple trichomes ca. 0,10,3 mm only along the margins; petioles (2,5)46 (8) cm long, lamina widely ovate, (6 x 3) 8 x 4 14 x 9,5 cm, apex acuminate, base subcordate to cordate (sinus to 13 mm deep), margin coarsely simply, rarely doubly serrate with (20)2533 teeth on each side, adaxial surface subglabrous with scattered short stinging hairs (ca. 12,5 mm), abaxial surface with scattered pubescence on the veins and with scattered stinging hairs. Inflorescences androgynous, lowest ones often pure male, upper one often pure female, ca. (25) 50100 mm long during anthesis, elongating to >12 cm in fruit, typically with male flowers in the proximal half and female flowers in the distal half, arising in pairs from the leaf axils, inflorescences irregularly interrupted, with flowers in ebracteose glomerules up to 5 mm long, female flowers ca. 1 mm long (including plumose stigma), perigon lobes finely pubescent, with (1) 35 stinging hairs each, male flowers with perigon ca. 1 mm long, perigon lobes finely pubescent, often with individual stinging hair; fruit ovate, ca. 1,251,5 x 1 mm with very short apex, largely included perigon in pubescent.

Specimens seen: Peru, Depto La Libertad, Prov. Sánchez Carrión, road Huamachuco to Cahgual, before Chugay and Aricapampa, S 07º48,626´, W 077º52,879´, 3005 m, 20.04.2004, Weigend & Schwarzer 7896 (B, HUT, USM). Prov. Otuzco, road Otuzco to Agallpampa, Motil, 19.04.2004, Weigend & Schwarzer 7886 (B, HUT, USM). Otuzco, 2900 m, 23.11.1973, López & Sagástegui 8051 (HUT). Debajo de Shitahuara (al Norte de Salpo), 3020 m, 1.4.1999, Leiva & Guevara 2274 (HAO, HUT, F). San Vicente-Chota (Otuzco), 3100 m, 6.12.1997, M. Rodríguez E. 356 (HUT). San Pedro (Motil, Otuzco), 2980 m, 10.5.1997, M. Rodríguez E. 18 (HUT). Distrito Agallpampa (Caserío La Florida), 2800 m, 4.4.2003, «ortiga negra», Medina 52 (HUT, HAO). Road Otuzco to Agallpampa, Motil, S 07º59,120´, W 078º 32,364´, 19.04.2004, Weigend & Schwarzer 7885 (B, HUT, USM). Prov. Santiago de Chuco: Santiago de Chuco, 2600 m, 20.6.1954, «ortiga de león», López 2340 (HUT). Prov. Pataz, road Chagual-Buldibuyo-Tayabamba, between Parcoy and Buldibuyo, S 08º07,162´, W 077º25,016´, 3564 m, 22.04.2004, Weigend & Schwarzer 7944 (B, HUT, USM). Between Parcoy and Buldibuyo, 5 km S of Parcoy, 22.04.2004, Weigend & Schwarzer 7932 (B, HUT, USM). Road Buldibuyo to Zona Maderas, Pass height towards Zona Maderas, S 08º05,106´, W 077º16,731´, 3775 m, 23.04.2004, Weigend & Schwarzer 7990 (B, HUT, USM). Depto. Ancash, Prov. Huaylas, Cordillera Negra, road (Caraz to) Huaylas to Jimbe, San Toribio, S 08º51,411´, W 077º55,098´, 3143 m, 29.04.2004, Weigend & Schwarzer 8070 (B, HUT, USM).
This new taxon is now known from La Libertad and neighbouring Ancash and is evidently closely allied to U. leptostachya. It differs from the latter in the often subscandent habit and the deflexed stinging hairs on the otherwise glabrous stem and most importantly by the presence of numerous stinging hairs on the perigon of the female flowers (and fruits) and individual stinging hairs on the perigon of the male flower. It is an extremely abundant species and its distribution probably marks all the areas with former forst cover it is often found in residual stands of Alnus acuminata. Its distribution area includes the region where U. urentivelutina is found, but that taxon is restricted to a seasonally dry forest fragment overlooking the Marañon river and differs in its very dense and soft indument and the distribution of male and female flowers in different inflorescences.
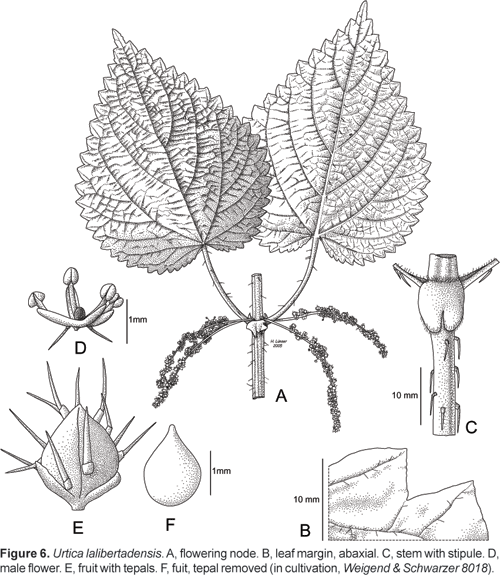
Urtica lalibertadensis replaces widespread U. leptophylla all across the Andes from the western slope to the eastern slope along the southern margin of the Amotape-Huancabamba Zone.
Urtica peruviana Geltman
Novon 8: 15. 1998. Type: Peru, Depto Cajamarca, Prov. Contumazá, El Tunel between Cascas & Contumazá, 2700 m, Sagástegui et al. 12630 (holo MO!, iso: HUA, HUT!).
Lianescent shrub 1.52 (3) m, more rarely erect, from ligneous rhizome to 10 mm thick, stems erect or lax, with scattered stinging hairs ca 1.5 mm long and very short, appressed, simple trichomes ca. 0.1 mm long. Leaves opposite, interpetiolar stipules united in pairs but deeply incised for ca. 50% of their length, subcircular, ca. 47 mm long and wide, with conspicuous white cistoliths and pubescent white simple trichomes ca. 0,10,5 mm; petioles 24 cm long, lamina widely ovate, 6 x 3 13 x 10 cm, apex acuminate, base rounded to subcordate (sinus to 5 mm deep), margin coarsely simply, rarely doubly serrate with 2030 teeth on each side, both surfaces very finely pubescent to subglabrous and covered with scattered short stinging hairs (ca. 12,5 mm), cistoliths largely punctiform. Inflorescences androgynous [with male flowers in the proximal portion (530 mm) and female flowers in the distal portion of inflorescences], or largely unisexual (lower ones largely or completely male, upper ones largely or completely female), ca. 3080 mm long during anthesis, elongating to >12 cm in fruit, arising in pairs from the leaf axils, inflorescences branched with some ebracteose glomerules scattered on lax branches up to 15 mm long, female flowers ca. 1 mm long (including plumose stigma), perigon lobes finely pubescent, without stinging hairs, male flowers with perigon ca. 1 mm long, perigon lobes finely pubescent; fruit ovate, ca. 1,51,7 x 1,31,7 mm with very short apex, largely included in pubescent perigon.
Specimens seen: Depto. Cajamarca, Contumazá, Cascas to Contumazá, near Tunel, 2700 m, 20.02.2002, Dostert & Rodriguez R. 2002/03 (BSB, M, HUT). 7,5 km from Contumazá on road to Guzmango, 27002800 m, 11.05.2003, Weigend et al. 7625 (BSB, M, USM, HUT). La Herilla (Guzmango), 2800 m, 2.04.1981, Sagástegui et al. 9704 (HUT, MO). Alrededores de Guzmango, 2700 m, 2.04.1981, Sagástegui et al. 9646 (HUT, MO). Alrededores de Guzmango, 2400 m, 21.04.1984, Sagástegui 11413 (HUT, MO). Arriba de Lledén, 2500-3000 m, 28.06.1983, Sagástegui et al. 10833 (HUT). Alrededores de Guzmango, 2500 m, 22.05.1978, Sagástegui & Mostacero 9132 (HUT, MO). Santiago, Guzmango, 2660 m, 30.05.1959, Sagástegui & Samamé 2935 (HUT, NY). Dto. San Benito, caserío La Montaña, 2440-2680 m, M. Binder et al. 1999/31 (HUT).
As noted by Geltman (1998), the species resembles U. leptophylla, U. longispica and U. macbridei at first glance, but it is the only species with distinct inflorescence branches and differs from U. longispica and U. macbridei in its larger fruits and from U. macbridei by its androgynous (not unisexual) inflorescencens. It is thus a well-characterized species. So far the species is only known from the forests of the Prov. Contumazá and it is replaced by U. lalibertadensis (see below) in the southern neighbouring Prov. Otuzco and by U. longispica in the northern neighbouring Prov. San Miguel de Pallaques.
Ribes andicola group (Grossulariaceae)
The Ribes andicola group in northern Peru
Ribes usually forms the upper margin of the relict forests where these forests grade into the jalca/paramo vegetation and especially in moderately disturbed habitats can form a broad transition zone between cloud forest and jalca which largely consists of a single species of Ribes. Also, in areas where the degradation of the forest vegetation has far progressed, Ribes (and Urtica and Hesperomeles, Rosaceae) are often the last witnesses of former forest cover and constitute the small hedges (cercos) between the fields.
The genus Ribes has numerous species in Peru, but only one species group has is so far known from the NW Peruvian relict forests. According to the existing taxonomy (Janczewski, 1907; Freire Fierro, 2004) these specimens would be placed into Ribes andicola Jancz., a somewhat heterogenous species group from Venezuela, Colombia and Ecuador. Ribes andicola has not been previously reported from Peru (Brako & Zarucchi, 1993). However, both from the upper margins of cloud forests in the Amotape-Huancabamba Zone and from hedges and forests remnants of formerly forested areas a large number of collections are now know which roughly correspond to the Ecuadorean species R. austroecuadorense and R. andicola. A closer examination shows that all these specimens from relict forests and montane forest of the eastern slope are actually specifically distinct from R. andicola, and only two of them can be tenatively placed in R. austroecuadorense, whereas the remaining material is here placed into three new species, one widespread in relict forests and forest remnants from Piura to Cajamarca (R. colandina), another apparently endemic to Contumazá (R. contumazensis) and a third one endemic to the subparamo of the eastern slope of the Andes (R. sanchezii).
Key to the species of the Ribes andicola group in northern Peru
1. Inflorescences erect.
Ribes austroecuadorense
1*. Inflorescences pendulous.
2. Leaves and inflorescences exclusively with stalked, glandular trichomes, without simple, white trichomes.
Ribes contumazensis
2*. Leaves and inflorescences with usually many simple white trichomes and additionally with few to many stalked glandular trichomes.
3*. Adaxial leaf surface shiny dark green, abaxial surface with simple, white trichomes only along the veins, lamina subentire with one very small lobe c. 1/3rd from base; fruits strongly resupinate.
Ribes sanchezii
3*. Ad- and abaxial leaf surface matt green and (mostly densely) pubescent from simple, white trichomes, lamina subentire to five-libed, fruits not resupinate.
4. Leaves typically 1530 x 1520 mm, entire or unevenly three-lobed, with lateral lobes absent or much smaller than central lobe, calyx dark red, but petals orange-yellow, fruits yellow.
Ribes andicola
4*. Leaves typically 3590 x 30100 mm, three- to five-lobed, with lateral lobes well developed, calyx and petals dark red, fruits dark red to black.
Ribes colandina
Formal taxonomy
Ribes andicola Jancz.
Bull. Int. Acad. Sci. Cracovie, Cl. Sci. Math. 10: 760. 1905. Lectoype (designated in Freire Fierro 2004: 56): Ecuador, Pichincha, Jameson 69 (AAU photo ex K, K).
See Freire Fierro 2004 for description and illustration.
Distribution: Venezuela (Mérida, Trujillo), Colombia (Valle de Cauca, Cauca), Ecuador (Pichincha, Chimborazo, Cotopaxi, Tungurahua, Bolívar, Napo, Azuay).
A somewhat variable species with varying degrees of pubescence on its leaves. The new species described below is at first glance quite similar, but differs in consistently black (versus yellow) fruits and distinctly larger and wider leaves and longer inflorescences. Ribes andicola appears to be absent from the extreme S of Ecuador, where it is replaced by R. austroecuadorense (see below), and consequently also from Peru. Our northernmost collection of the R. andicola group (Piura Huancabamba: Weigend & Dostert 98/271) has the black fruits typical of R. colandina, not the yellow fruits typical of R. andicola.
Ribes austroecuadorense Freire Fierro
Novon 8: 356. 1998. Holotype: Ecuador, Prov. Loja, Parque Nacional de Podocarpus, E of Nudo de Cajanuma, S 04º05´, W 79º10´, 31003200 m, Øllgard 90339 (QCA, isotypes: AAU, QCNE).
See Freire Fierro 2004 for description and illustration.
Distribution: Ecuador (Loja, Azuay), Peru: Depto Piura, Prov. Huancabamba, km 3842 on road from Huancabamba to (Sondor) Tabaconas, mixed jalca and forest remnants, 26002700 m, 18.5.1998, Weigend & Dostert 98/211 (M, USM). Depto Cajamarca, Prov. Chota, Miracosta, between Miracosta and Pampa del Lirio, 3150 m, 11.11.2000, Sánchez Vega et al. 10300 (B, CPUN).
The determination of these two specimens as R. austroecuadorense is still somewhat tenuous the specimen from Piura is sterile, but agrees well with the leaf characters described in Freire Fierro (2004). The Cajamarca-specimen is a male plant in full flower and appears to have the typical erect (not pendulous) inflorescences of R. austroecuadorense. Cleary, both localities should be revisited at other times of the year to clarify the identity of these specimens.
Ribes colandina Weigend, spec. nov.
Holotype: PERU, Depto. La Libertad, Prov. Bolívar, Bolívar, path to Laguna Shopol, 32803600 m, S 10º17´ 57´´, W 078º02´ 50´´, 18.10.2000, Weigend et al. 2000/823 (holotype: USM; isotypes: HUT, M, B).

Ribes fruticosa, usque ad 4 m alta, dioica; folia magna, triloba vel sub-qinqueloba, utrinque latere densissime pubescentes; racemis pendulis (10) 3050-floris, floribus vineis, pubescentibus, baccis nigris, sparse pilosis.
Dioecious shrub (1) 1,54 m tall; very densely to moderately tomentose from simple, curly trichomes 12 mm long and with scattered subsessile glands, especially on young shoots and the abaxial leaf surface. Leaves deciduous, petiole 1535 mm long, 1 mm wide; stipules well differentiated, united with petiole for 610 mm, stipulate region 35 mm wide, free apex of stipules not differentiated, distal margin deeply laciniate with plumose setae up to 2 mm long; lamina subcoriaceous, widely ovate to subcircular; 3590 x 30100 mm, distinctly three- to five-lobed with central lobe slightly (1,52 x) longer than lateral lobes, lobes triangular-ovate, free portion of central lobe up to 50 x 40 mm, margin irregularely serrate to lobulate; lobules up to 9 x 14 mm, with up to 4 serrations per lobule, base rounded to deeply cordate (sinus to 15 mm deep); adaxial surface subglabrous with scattered simple hairs 0,11 mm long, eglandular, abaxial surface densely to very densely pubescent (often appearing snow-white) with simple hairs up to 2 mm long, without or with scattered sessile glands. Inflorescences terminal on short lateral shoots (brachyblasts); racemes pendent with (10) 3050 flowers and 4080 (120) mm long, peduncle 510 (15) mm long; densely pubescent with numerous simple hairs 1 mm long, eglandular or with scattered sessile glands; pedicels c. 1 mm long, c. 24 mm apart in open flowers; bracts 46 x 1 (2) mm, narrowly ovate, bracteoles 1 x 0,5 mm, narrowly ovate to linear, persistent. Flowers narrowly cyathiform, calyx and corolla very dark red; 4 x 35 mm; covered with simple hairs 0.2 mm long, ovarian portion glandular; ovary conical, 1 x 1 mm; hypanthium 12 mm long; calyx lobes ovate acuminate, 2 x 1,5 mm, erect or half-spreading; petals inserted c. 1 mm from base of hypanthium, 1 x 0.8 mm, narrowly ovate; filaments inserted c. 0,5 mm from base of hypanthium; filaments and anthers 0,3 (female) to 0,5 mm (male) long; style apically bifid, 1 mm long. Fruit spherical, 812 mm in diameter, with scattered shortly stalked glands, pendulous, black.
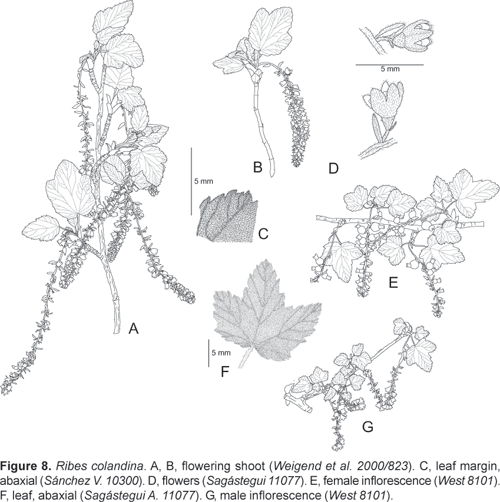
Peru: Depto. Piura. Prov. Ayabaca: Frías, Las Pircas, 3000 m, 17.10.1988, Diaz & Vasquez 3003 (MO). Prov. Huancabamba: Huancabamba 29003100 m, 20.5.1998, Weigend & Dostert 98/271 (M). Depto Lambayeque. Prov. Ferreñafe, Sinchihual, Incahuasi, 3400 m, 3.10.1985, Llatas Quiroz 1294 (F); around Incahuasi, 2750 m, 13.9.1985, Sagástegui et al. 12875 (F). Depto. Amazonas. Prov. Chachapoyas footpath from Granada to Siete Lagunas, 3600 m, 4.12.1999, T. Hofreiter & Th. Franke 1/172 (B, HUT, M, USM). Depto. Cajamarca: Prov. Contumazá, Road Contumazá to Guzmango, 5-10 km W of Contumazá, 2600 m, 20.6.1998, Weigend et al. 98/564 (M). 5 km from Contumazá on road to Cascas, 26002700 m, 10.05.2003, Weigend et al. 7607 (B, HUT, M, USM). 22 km from Contumazá on road to Guzmango, above Cruz Grande, 30003600 m, 11.05.2003, Weigend et al. 7627 (B, HUT, M, USM), same locality, Weigend et al. 7638 (B, HUT, M, USM). 7.5 km from Contumazá on road to Guzmango, above Cruz Grande, 2700-2800 m, 11.05.2003, Weigend et al. 7624 (B, HUT, M, USM). - Prov. Cajamarca, between Hualgayoc and Bambamarca, 26503550 m, 16.11.1948, R. Scolnik 1349 (NY). W of Cajamarca, road to Cumbemayo, 3350 m, 24.9.1991, Sánchez Vega 5817 (F); Huacataz, 20 km NE of Cajamarca, 3000 m, 21.11.1981, Sánchez Vega & Medina 2687 (USM, F). Cerro Sexcemayo, W of Cajamarca, 3430 m, 2.4.1991, Vega 5426 (F). Surroundings of Cajamarca, 27003700 m, 5.6.1986, Becker et al. 1339 (USM). - Prov. Celendin, near Agua Colorada on road Cajamarca to Celendin, 3324 m, S 06º58'19, W 078º11'16, 15.10.2000, Weigend et al. 2000/760 (M, B, USM, HUT). 64 km E of Cajamarca on the road to Celendin, 3500 m, 5.2.1997-2.4.1997, Weigend et all. 97/372 (F, M). Ca. 10 km before Celendin on road from Cajamarca, 3100 m, 06°55'11"S, 78°11'23"W, 16.10.2000, Weigend et al. 2000/779 (B, HUT) - Prov. Chota, Cangana above Chota, 2700 m, 6.10.1993, Llatas Quiroz et all. 9396 (USM). Miracosta, between Miracosta and Pampa del Lirio, 3150 m, 11.11.2000, Sánchez Vega et al. 10300 (CPUN, B). - Prov. San Miguel de Pallaques, Agua Blanca to Prado-San Miguel, S 07º00'57,0", W 079º02'31,2", 2900 m, 11.05.2001, Henning & Schneider 194 (HUT, BSB, M, USM). Above Agua Blanca, Cerro Quillón, 33203500 m, S 07º01'54", W 079º04'10", 14.10.2000, Weigend et al. 2000/739 (HUT, B, M, USM). Road San Miguel to Tongod, S 06º55'09", W 078º48"22", 3434 m, 02.05.2003, Weigend et al. 7495 (B, USM, HUT, M). Cerro Minis, Distrito Unión Agua Blanca, 2980 m, 16.02.2000, Rodríguez et al. 2288 (F, HUT, M, NY). El Tingo, Dist. Unión Agua Blanca, 3200 m, 17.02.2000, Rodríguez et al. 2310 (F, HUT, MO, NY). El Tingo (camino Taulis), Dist. Unión Agua Blanca, 3000-3250 m, 18.02.2000, Rodríguez et al. 2359 (F, HUT, M, NY), 2367 (HUT, NY). - Prov. San Marcos, below of Totorilla, Río Shitamalca, 3450 m, 31.10.1992, Vega & Torres 6415 (F, MO). Depto. La Libertad: Prov. Otuzco, Chota between Yamoabamba und Shorey, 2900 m, 15.11.1983, Sagástegui et al. 11077 (NY). Prov. Bolívar, Balsas to Bolívar, 72 km from Balsas, 3600 m, S 07º04' 16", W 077º50'05", 17.10.2000, Weigend et al. 2000/820 (HUT, M, USM, B). - Prov. Pataz, road Huamachuco-Chagual-Buldibuyo, between Parcoy and Buldibuyo, 5 km S of Parcoy, 22.04.2004, Weigend & Schwarzer 7934 (B, HUT, USM). Prov. Huamachuco, outskirts of town, 26.11.1936, West 8101 (HUH, UC), same locality, Weigend et al. 97/253 (F, M).
This new species replaces R. andicola just south of the Ecuadorean border and appears to be very widespreach in Piura, Lambayeque, Cajamarca and La Libertad. A close search in Ancash yielded not a single specimen of this taxon, it is there replaced by R. viscosum. The material here referred to this new species is slightly heterogenous and includes some very small leaved specimens from the southern part of the range (e.g., West 8101). Also, documentation is highly unsatisfactory from Piura and Lambayeque and many apparent distribution gaps in areas of known subparamo vegetation show a considerable degree of undercollection. Ribes colandina differs from Ribes andicola in uniformly dark red flowers (petals red, not orange-yellow as in R. andicola), black instead of yellow fruits and wider, more deeply lobed leaves.
Etymology: the specific epithet is an anagramm of the epithet of its apparent closest relative, R. andicola.
Ribes sanchezii Weigend, spec. nov.
Holotype: Peru, Depto. Amazonas, Prov. Chachapoyas, Balsas road to Leymebamba, just below Abra Callacalla, 3559 m, S 06º43' 49", W 077º53'18", 19.10.2000, M. Weigend et al. 2000/862 (M, B, BM, USM, HUT).

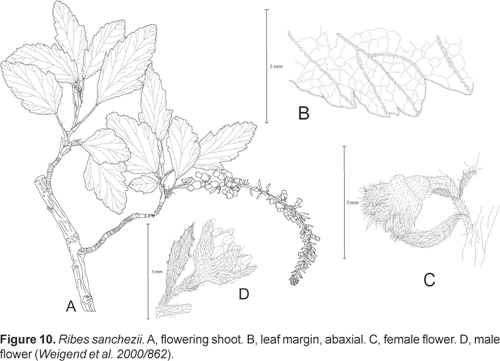
Ribes fruticosa, usque ad 4 m alta, dioica; folia magna, sub-triloba, supra obscure virides, glabrescentes, infra sparse pubescentes; racemis 3050-floris, floribus vineis, pubescentibus et sparse glandulosibus.
Dioecious shrub (1) 1,54 m tall; shoots and adaxial leaf surfaces sparsely pubescent to glabrous, abaxial leaf surfaces, inflorescence axis, petioles moderately pubescent from simple, trichomes c. 1 mm long, leaves deciduous, petiole 1525 mm long, 1 mm wide; stipules well differentiated, united with petiole for 610 mm, stipulate region 35 mm wide, free apex of stipules not differentiated, distal margin deeply laciniate with plumose setae up to 2 mm long; lamina subcoriaceous, ovate to elliptical; 4065 x 3050 mm, subentire with one very small lobe c. 1/3rd from base, lateral lobes up to 510 x 10 mm, central lobe much (45 x) longer than lateral lobes, lobes ovate, margin irregularely lobulate; lobules up to 10 x 15 mm, with up to 4 serrations per lobule, base cuneate, rarely nearly truncate; adaxial surface subglabrous, dark green and shiny, with very few simple hairs 0,10,5 mm long towards the margin, eglandular, abaxial surface densely short pubescent only on primary and secondary veins with with simple hairs up to 1 mm long, eglandular. Inflorescences terminal on short lateral shoots (brachyblasts); racemes pendent with 3050 flowers and 80130 mm long, peduncle 510 (15) mm long; densely pubescent with numerous simple hairs 1 mm long, eglandular or with scattered stalked glands c. 05. mm long; pedicels c. 1 mm long, c. 24 mm apart in open flowers; bracts 46 x 1 (2) mm, narrowly ovate, bracteoles 1 x 0.5 mm, narrowly ovate to linear, persistent. Flowers narrowly cyathiform, calyx and corolla dark red; 5 x 5 mm; covered with simple hairs 0.2 mm long, ovary with 15 stalked glands and very few simple trichomes; ovary conical, 1 x 1 mm; hypanthium 12 mm long; calyx lobes ovate acuminate, 2 x 1.5 mm, erect or half-spreading; petals inserted c. 1 mm from base of hypanthium, 1 x 0,8 mm, narrowly ovate; filaments inserted c. 0,5 mm from base of hypanthium; filaments and anthers 0,3 (female) to 0,5 mm (male) long; style apically bifid, 1 mm long. Fruit strongly resupinate, otherwise unknown.
Peru: Depto. La Libertad, Prov. Bolívar, between Sant Luisa and Uchumarca turnoff, S 07º04', W 077º49', 3700 m, 11.11.2001, Sánchez Vega et al. 11192 (CPUN, B).
This new species of Ribes is very clearly distinct form both R. andicola and R. colandina because of its ovate to elliptical leaves with a very poorly developed lateral lobe and particularly its aberrant indument: Both other taxa have leaves with pubescence (albeit in varying densities) on both the adaxial and abaxial surface and the adaxial leaf surface is matt green, whereas R. sanchezii a shiny dark green upper leaf surface and pubescence abaxially restricted to the primary and secondary veins. Ribes sanchezii also has strongly resupinate fruits, whereas the fruits of R. andicola and R. colandina are pendulous. It replaces widespread R. colandina on the wetter eastern slopes of the Cordillera Oriental and forms dense stands at the upper limit of the cloud forest at the Abra CallaCalla, where it is one of the most abundant shrub species in the vegetation.
Etymology: Named in honour of the distinguished Peruvian botanist Isidoro Sánchez Vega of Cajamarca, who made the second known collection of this taxon.
Ribes contumazensis Weigend, spec. nov.
Holotype: Peru, Depto. Cajamarca, Prov. Contumazá: Between Magdalena and Contumazá, 3100 m, Dec. 1916, Weberbauer 7221 (HUH!, isotype: US!).
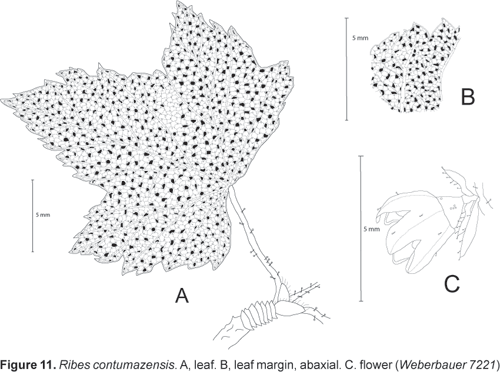
Ribes fruticosa, usque ad 2 m alta, dioica; folia parva, triloba vel sub-quinqueloba, utrinque latere dense glandulosa; racemis 3050-floris, floribus luteo-rubris, glandulosibus.
Dioecious shrub 12 (4?) m tall; shoots and adaxial leaf surfaces covered only with scattered stalked glands < 0,5 mm long. Leaves deciduous, petiole 1015 mm long, 1 mm wide; stipules well differentiated, united with petiole for 34 mm, stipulate region 34 mm wide, free apex of stipules not differentiated or triangular, glandular-serrate; lamina subcoriaceous, subcircular; 2530 x 2530 mm, with one distinct lobe on each side, lateral lobes up to 510 x 10 mm, central lobe much (1,52 x) longer than lateral lobes, lobes ovate, margin irregularely lobulate; lobules up to 5 x 5 mm, with up to 4 serrations per lobule, subcordate, rarely nearly truncate; adaxial surface subglabrous, eglandular, abaxial with scattered stalked glands especially on primary and secondary veins. Inflorescences terminal on short lateral shoots (brachyblasts); racemes pendent with 1525 flowers and 3050 mm long, peduncle 510 (15) mm long; with scattered stalked glands, pedicels c. 1 mm long, c. 24 mm apart in open flowers; bracts 1,52 x 1 mm, narrowly ovate, bracteoles 1 x 0,5 mm, narrowly ovate to linear, glabrous, glandular-serrate. Flowers narrowly cyathiform, brownish yellow; 5 x 5 mm; covered with scattered glandular trichomes, ovary with 15 stalked glands and very individual few simple trichomes (< 5 per flower); ovary conical, 1 x 1 mm; hypanthium 12 mm long; calyx lobes ovate acuminate, 2 x 1,5 mm, erect or half-spreading; petals inserted c. 1 mm from base of hypanthium, 1 x 0,8 mm, narrowly ovate; filaments inserted ca. 0,5 mm from base of hypanthium; filaments and anthers 0,3 (female) to 0,5 mm (male) long; style apically bifid, 1 mm long. Fruit unknown.
Only known from type collection.
In spite of the fact that a lot of collections are generally known from the Contumazá area, not least because Abundio Sagástegui Alva of Trujillo (HAO) has worked there for a very long time, the only collection we have of this taxon is by Weberbauer. I hestitated for a long time before recognizing this species as distinct, but the fact that it is completely glabrous apart form the stalked glands clearly separates it from R. colandina from the same region. Additional collections of this taxon are clearly desirable.
Conclusions
Both groups here studied, the Ribes andicola group and the rhizomatous species of Urtica are present with often narrowly endemic species in the Amotape-Huancabamba Zone and more especially in the relict forests of NW Peru. Three new species of Ribes and two new species of Urtica are described, three of these new species are found in the relict forests, and two of them are endemic to them. Additionally, two taxa previously only known from Ecuador, Ribes austroecuadorense and Urtica longispica are reported from the forest fragments of NW Peru (Table 1). While this in total signifies a considerable increase in known diversity for the region in general and these forests in particular, a close examination reveals that our knowledge of the area is still grossly insufficient.
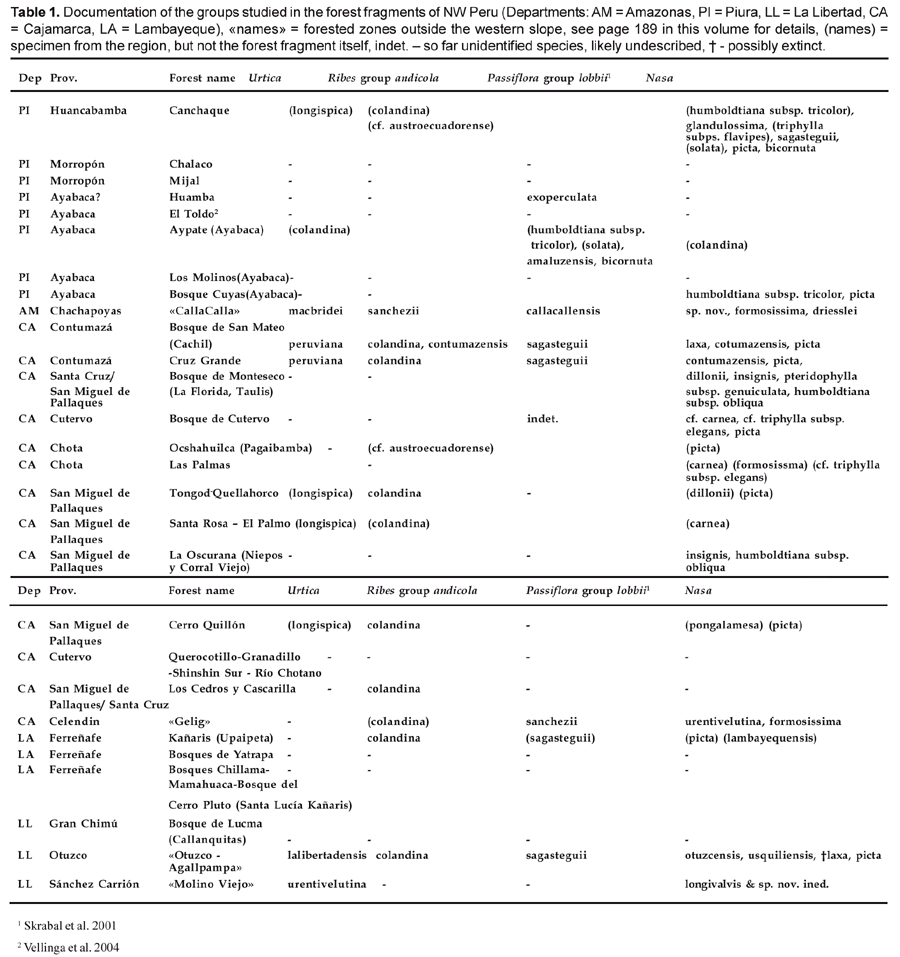
State of our knowledge
Both groups studied in Ribes and Urtica can be considered as indicative of humid conditions and can be expected to be present in any montane forest fragment in this region. However, if we summarize the locality data, we find that we still lack any collections of these two plant groups from a large number of known forest fragments. Table 1 summarizes the number taxa known from individual forest fragments in four plant groups: Urtica and the Ribes andicola group (see above), the Passiflora lobbii group (Skrabal et al., 2001) and Nasa (Dostert & Weigend, 1999; Weigend, 2000, 2002, 2004, 2005; Weigend & Rodrgiuez, 2001, 2003; Weigend et al., 1998, 2003). All four groups can be expected to be present in every major relict forest with at least one species. There are more than 20 forest fragments recognized here, but there are only 2 forest fragments (San Mateo and Cruz Grande in Contumazá) and a «region» (Otuzco-Agallpampa) from which representatives of all four groups are currently documented, another five regions have at least three of the four groups documented (Canchaque, Kañaris, Cerro Quillón and Tongod). Both Urtica and Passiflora are currently known from less than half of the forest fragments. Moreover, moderately large (< 3000 ha), but well explored relict forests have 47 taxa of Nasa (Monteseco, Canchaque), whereas the huge relict forests of Kañaris (ca. 14000 ha) has only two species documented. It is immediately apparent that the documentation of the flora of these forests is extremely incomplete. New taxa are still expected in this region. A typical example is a new species of Ribes of the Ribes macrobotrys group, which is so far only known from a sterile specimen [Depto. Cajamarca, Prov. San Miguel, Cerro Quillón, El Tingo, Agua Blanca, 3445 m, 17.02.2000, E. Rodriguez R. et al. 2333 (HUT, NY)].
Under-documentation of the Passiflora lobbii group is probably due to the fact that they are near-ubiquitous, but difficult-to-find plants, which moreover flower only part of the year and are not usually collected in the sterile stage (even though they can be determined in the vegetative state). Similarly, Ribes has a very short flowering period at the end of the dry season (usually October) and is not usually collected in fruit or in the sterile state. Urtica is present all around the year, but is often not collected for other, obvious reasons.
Indicators of former montane forests
Both groups studied in Ribes and Urtica indicate sites with humid conditions with former forest cover and can be considered as important markers for forests which have been completely destroyed. They persist in secondary vegetation for a much longer time, than most forest taxa, e.g. in Loasaceae, Orchidaceae etc. Many of the collections of Urtica and Ribes from outside present-day forest thus indicate the former presence of such forests. Particularly striking cases are the stands of U. lalibertadensis which, together with the abundance of scattered trees of Alnus acuminata, seem to indicate that montane forests once covered large parts of the the Provinces Otuzco and Pataz, where there are now barely any recognizable remnants left. Similarly, the near universal presence of Ribes colandina, Urtica leptostachya and scattered trees of Alnus acuminata and Hesperomeles in hedges and gullies strongly indicate that the Province San Miguel was at some stage largely covered with montane forests, of which now only a few fragments remain. The large communities of Ribes colandina above Guzmango (Prov. Contumazá) or near Celendín (Prov. Celendín), together with other floristic elements, clearly show that these areas once bore montane forest, where there is now degraded scrubland. The present habitat of U. urentivelutina is entirely secondary (in hedges) and there is possibly no undisturbed forest left in the area.
The value of these indicator species in reconstructing the ancient forests of NW Peru is underscored by the fact that there are some historical collections of more sensitive forest plants from areas which are now no longer suitable for their existence: Narrowly endemic Nasa dilloniana, currently known only from the Bosque de Monteseco, is also known from a historical collection (pre-1900) from Santa Rosa near San Miguel de Pallaques, where the species is now apparently extint and no undisturbed forest remains, U. longispica is still abundant in the area.
Relationships between the relict forests
At present, none of the plant groups studied here (Ribes, Urtica) or elsewhere (Nasa, Passiflora, see above) is documented comprehensively from the relict forests and the surrounding areas. However, on the basis of what data we have, we can already draw some conclusions. There is apparently a relatively close relationship between the forest fragments of San Miguel de Pallaques and Santa Cruz, with the presence (current or historical) of Nasa dilloniana, N. humboldtiana subsp. obliqua and N. insignis in the Bosque de Oscurana, near San Miguel and in Monte Seco. This complex of forest remnants also presents the southernmost known localities for Urtica longispica, which is otherwise known from Piura (and Ecuador and Colombia). This San Miguel/Monte Seco complex apparently represents the southern end of the western Andean forest elements and its closest relationships at species and species group level appear to be towards Ecuadorean taxa. The forests of Lambayeque and Piura are at present so poorly known, that little can be said about them, but what taxa are known from the area (U. longispica, N. sagasteguii, N. glandulosissima) appear to confirm this view. Similarly, the apparent presence of Ribes austroecuadorense in Chota and Huancabamba also ties this region to Ecuador.
Both in Nasa and in Urtica the species are replaced by a new set of taxa in crossing the Jequetepeque river. Contumazá (San Mateo and Guzmango) have many local endemics (N. contumazensis, U. peruviana, R. contumazensis), but also share some elements with Otuzco (N. laxa).
Very little is currently known for the plant groups here studied in either Cutervo or Kañaris and this is possibly the region which is in the most urgent need of study. We expect a relatively close floristic relationship between these areas, and we also expect that the actual diversity in these two areas, especially in the huge forests of Kañaris, is very high.
Apart from these tentative relationships the most striking phenomenon is the transandean distribution of the few more widespread taxa: R. colandina and U. lalibertadensis are widespread from the W slope of the Cordillera Occidental to the west-facing slopes of the mountains overlooking the Marañón (the northermost part of the Cordillera Central) and are replaced by local endemics (e.g. U. urentivelutina) on the drier east-facing slopes of the mountains overlooking the Marañón and by other species on the very wet eastern (amazonian) slope of the Cordillera Central (U. macbridei, R. sanchezii and species from other groups of Ribes).
Outlook
The data presently available on the flora of the relict forests of NW Peru are highly unsatisfactory. There are probably still a very considerable number of undescribed species to be discovered. Documentation of the biodiversity in the form of herbarium collection is currently fragmentary at best.
The former extent of the montane forests can probably be inferred from the current distribution of tenacious members of the secondary vegetation such as Urtica, Ribes and Alnus. Their present distribution strongly indicates that the former extent of these forests was very much larger than at present. Large parts of the Prov. Otuzco (La Libertad) - currently without any coherent forest cover - were probably completely and coherently forested. The relict forests of the provinces Santa Cruz and San Miguel de Pallaques (Tongo, Oscurana, Santa Cruz, Cerro Quillón, etc.) probably had a more or less coherent forest belt, possibly with interspersed paramo islands, not _ as now _ largely open vegetation with forest fragments and relict species in quebradas and hedges. Similarly, the forests of Guzmango and Contumazá were probably more or less coherent and there are at least close floristic links to the Otuzco region, but little is currently known about the intervening region. A detailed mapping of the distribution of the Ribes andicola groups, of Urtica and Alnus for the Amotape-Hunacabamba Zione could probably give a detailed picture about the former overall extent of montane forests in the region.
Literature cited
Berger, A. 1924: A taxonomic review of currants and gooseberries. - New York Agric. Exp. Sta. Techn. Bull. 109: 3-118. [ Links ]
Brako, L. & J.L. Zarucchi 1993. Catalogue of the Flowering Plants and Gymnosperms of Peru. Monogr. Syst. Bot. Missouri Bot. Gard. 45.
Britton, N. L. & Brown, A. 1913: An illustrated Flora of the Northern United States and Canada 3: 236-241. - New York.
Dostert, N. & Weigend, M. 1999: A synopsis of the Nasa triphylla complex (Loasaceae), including some new species and subspecies. Harvard Papers in Botany 4: 439-467.
Freire F., A. 1998: New species of Ribes (Grossulariaceae) from South America. - Novon 8: 354-358.
Freire Fierro, A. 2004. 75B. Grossulariaceae. In: G. Harling & L. Andersson, Flora of Ecuador 73: 41-66.
Geltman, D.V. - 1998 - New species and new combinations of Urtica (Urticaceae) in South America. Novon 8: 15-17.
Janczewski, E. 1907: Monographie de Grosseillier. - Mem. Soc. Phys. Genève 35: 199-517.
Jørgensen, P.M. & Y. León, S.: Checklist of the Vascular Plants of Ecuador. Monogr. Syst. Bot. Missouri Bot. Gard. 75.
Koepcke, H. W. 1961: Synökologische Studien an der Westeseite der peruanischen Anden. Bonner Geogr. Abh. 29: 1-320.
Lingt-ti, Lu 1995: A study on the genus Ribes L. in China. - Acta Phytotax. Sin. 33: 58-75.
Macbride, J. F. 1941: Saxifragaceae, Flora of Peru 3(2). - Publ. Field Mus. Nat. Hist., Bot. Ser. 13: 1015-1025.
Moerman, D.E. 1998: Native American ethnobotany. - Portland/Oregon: Timber Press.
Ragonese, A. E. & Martínez C., R. 1947: Plantas indígenas de la Argentina con frutos comestibles. - Revista Invest. Agríc. 1: 147-216.
Skrabal, J., Tillich H.-J., Weigend, M. 2001: A revision of the Passiflora lobbii group (Passifloraceae), including some new species and subspecies. Harvard Papers in Botany 6: 309-339.
Taylor, Ch. 2003: Urticaceae. In: C. Marticorena & R. Rodrogiuez: Flora de Chile 2(2): 44-61.
Vellinga, Willem-Pier, Jeremy N. M. Flanagan & Todd R. Mark 2004: New and interesting records of birds from Ayabaca province, Piura, north-west Peru. Bull. B. O.C. 124(2): 124-142.
Weddell, H.A. 1856: Monographie de la famille des urticacées. Arch. mus. hist. nat. Paris 9: 1-592.
Weddell, H.A. 1869: Urtica. - In: Candolle, A.D. de: Prodr. Syst. Nat. 16/1: 39-67.
Weigend, M. & Binder, M., in prep.: Three new species of Ribes (Grossulariaceae) from Central and South America. - Syst. Bot., submitted 15.12.00.
Weigend, M. & E. Rodriguez R. 2003: A revision of the the Nasa stuebeliana group [Nasa ser. Saccatae (Urb. & Gilg) Weigend, Loasaceae] with notes on morphology, ecology, and distribution. Botanische Jahrbücher für Systematik und Pflanzengeographie 124: 345-382.
Weigend, M. & E. Rodriguez R., 2001: Nasa picta Hook.f. subsp. pamparomasii (Loasaceae), a new subspecies of Nasa picta from Ancash, Peru. Arnaldoa 7(1-2): 19-26.
Weigend, M. 2000: A revision of the Peruvian species of Nasa ser. Alatae in Peru. Nordic Journal of Botany 20: 15-32.
Weigend, M. 2002: Observations on the Biogeography of the Amotape-Huancabamba Zone in Northern Peru. - In: K. Young, C. Ulloa U., J. L. Luteyn & S. Knapp, Plant Evolution and Endemism in Andean South America. Botanical Review 68: 38-54.
Weigend, M. 2004: Additional Observations on the Biogeography of the Amotape-Huancabamba Zone in Northern Peru: Defining the South-Eastern limits. Rev. Peru. Biol. 11(2): 127-134.
Weigend, M. Henning, T. & Schneider, C. 2003: Notes on the Systematics, Morphology, Distribution and Pollination of Nasa Ser. Carunculatae (Loasaceae subfam. Loasoideae). Systematic Botany 29: 765-781.
Weigend, M., 2004: Four New Species of Nasa ser. Alatae (Loasaceae) in the Amotape-Huancabamba Zone of Peru. Novon 14: 134-146.
Weigend, M., E. Rodriguez R. & N. Dostert 1998: Nasa insignis y Nasa glandulosissima, dos especies nuevas de Nasa con hojas peltadas del norte del Perú. Arnaldoa 5: 151-157.
Woodland, D. W. - 1982 - Biosystematics of the perennial North American Taxa of Urtica. II. Taxonomy. Syst. Bot. 7(3): 282-290.
Correspondencia
2 Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos. Aptdo. 14-0434, Lima 14, PERÚ
3 Herbarium Truxillense (HUT), Universidad Nacional de Trujillo, Jr. San Martín 392, Trujillo, PERÚ
