










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.12 no.3 Lima Oct./dic 2005
Rolando Aquino1, José Alvarez2 y Augusto Mulanovich
1. Instituto de Ciencias Biológicas Antonio Raimondi, Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos y Estación Experimental del IVITA-Iquitos.
2. Programa Biodiversidad, Instituto de Investigaciones de la Amazonía Peruana (IIAP), Iquitos.
Presentado: 20/10/2005
Aceptado: 22/12/2005
Resumen
Este trabajo contiene información del inventario, abundancia y estado de conservación de las especies de primates que habitan en los bosques de las Sierras de Contamana. Hemos registrado 87 grupos correspondientes a 11 especies. De ellas, Saguinus mystax Spix, Ateles chamek Humboldt, Cebus apella Linnaeus y Lagothrix poeppigii Schinz fueron las más avistadas con 14, 13, 13 y 11 grupos, respectivamente. El rango de variación del tamaño de grupo resultaron similares a los registrados para otras áreas, con excepción de A. chamek, cuyos grupos en la mayoría de los casos estuvieron compuestos por más de 16 individuos. En el área de estudio, las poblaciones más abundantes correspondieron al Cacajao calvus ucayalii Thomas (479 individuos/100 km), L. poeppigii (178,5 individuos/100 km) y A. chamek (148,2 individuos/100 km). los mayores avistamientos ocurrieron en los bosques de terraza alta (46%) y de colina baja (27%), constituyéndose así en los hábitats de mayor uso por los primates. Entre las especies registradas, C. calvus ucayalii y S. mystax aún no están protegidas en ninguna de las actuales áreas naturales protegidas del Perú.
Palabras claves: Primates, estado de conservación, Sierra del Divisor, Sierra de Contamana, biodiversidad, Perú .
Abstract
This paper provides information on the diversity, abundance and conservation status of primates species that inhabit the forests of the Sierras de Contamana. We recorded 87 groups corresponding to 11 species. Saguinus mystax Spix, Ateles chamek Humboldt, Cebus apella Linnaeus and Lagothrix poeppigii Schinz were the most frequently sighted with 14, 13, 13 and 11 groups, respectively. The variation of group size was similar to those of groups in other areas, except for A. chamek whose groups in most of the cases consisted of more than 16 individuals. In the study area, highest population were observed for Cacajao calvus ucayalii Thomas (479 individuals / 100 km), L. poeppigii (178,5 individuos/100 km) and A. chamek (148,2 individuos/100 km). Most sightings occurred in high terrace forests (46%) and low hill forests (27%), constituting the habitats most frequently used by primates. Among the recorded species, C. calvus ucayalii and S. mystax are not still protected in any of the current protected natural areas of Peru.
Keywords: Primate, conservation status, Sierra del Divisor, Sierra de Contamana, biodiversity, Peru.
Introducción
Las Sierra de Contamana forman parte de la Sierra del Divisor y están ubicadas en la margen derecha del río Ucayali, muy cerca de la frontera con Brasil. Políticamente pertenece a los Departamentos de Loreto y Ucayali. Para el Perú, este sector de la Amazonía es reconocida como una de las áreas con vacíos de información (Rodríguez,1997), de modo que los estudios que puedan realizarse adquieren especial importancia porque nos permitirán conocer la diversidad de este sector. Por otro lado, los estudios de primates en este sector amazónico nos dan la oportunidad de conocer los diversos tipos de hábitats en que viven; aunque algunas especies no están presentes en otras áreas de la región amazónica, pero igualmente se encuentran amenazados por las diversas actividades del hombre, entre ellas la l tala y extracción maderera, la quema con fines agrícolas y de ganadería y por ultimo la caza, muchas veces con fines comerciales.
Estudios previos en las Sierras de Contamana no existen. Las evaluaciones más cercanas sobre diversidad biológica fueron realizadas en noviembre del 2000 en el sector Contamana Aguas Calientes y luego en enero del 2001 en el sector Abujao Shesha (FPCN; CDC. 2001). En la primera dan cuenta del registro de 12 especies de primates, mientras que en la segunda de 13 especies. En ambos sectores destacaron los primates de tamaño mediano y pequeño.
En un esfuerzo por llenar este vacío de información para el área que nos ocupa, El Centro de Datos para la Conservación de la Universidad Nacional Agraria La Molina (CDC-UNALM) conjuntamente con PRO-NATURALEZA y The Nature Conservancy (TNC), dentro del proyecto «Apoyo a la Conservación de la Sierra del Divisor» y con el apoyo financiero de la Fundación Moore ejecutaron una Evaluación Ecológica Rápida (EER) en las Sierras de Contamana, donde tuvimos la oportunidad de participar. Uno de los componentes de la fauna mayor evaluada fueron los primates que se encuentran habitando los diferentes hábitats de las Sierras de Contamana. Un análisis del inventario y evaluación en lo que concierne a los primates es presentado aquí y está basado en el levantamiento de información directamente en el campo durante las incursiones a los bosques entre el 19 de octubre al 01 de noviembre del 2004.
Área de estudio
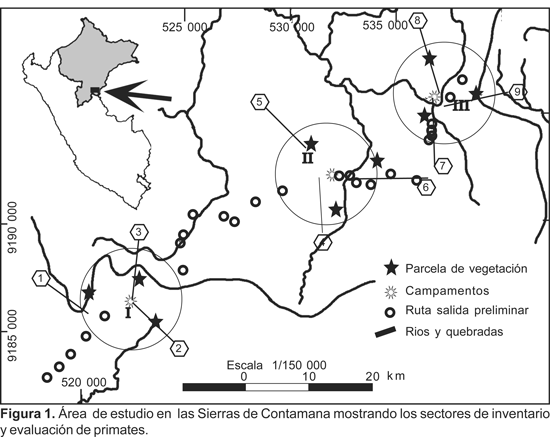
El área de estudio denominada Sierras de Contamana se encuentra ubicada al sureste de la ciudad de Contamana, aproximadamente a 16 km de longitud desde la orilla del río Ucayali (Fig. 1). El acceso a ella fue por la trocha carrozable abierta hace aproximadamente 15 años para la extracción de madera, cuyo punto de partida es el denominado Punto 8 de propiedad de MAPLE GAS.
En el área de estudio, el inventario y la evaluación fueron llevados a cabo en los siguientes tipos de bosques: a) bosque de terraza alta, b) bosque de colina alta con vegetación achaparrada, c) bosque de colina media con vegetación alta, d) bosque de colina media con vegetación achaparrada, e) bosque de colina baja, f) aguajal de altura y g) bosque secundario. De ellos, predominaron los bosques de terraza alta y de colina baja, en particular en los alrededores del Campamento II.
En los bosques de terraza alta y de colina baja, la cobertura vegetal estuvo conformada por árboles de fustes gruesos y de gran envergadura que alcanzaron entre 25 a 30 m de alto, con algunos emergentes arriba de los 35 m, muchos de ellos de copas amplias y con enmarañados de bejucos y lianas, asociados con palmeras como Scheelea sp., Astrocaryum sp., Attalea spp. Socratea sp. y Oenocarpus bataua Burret, entre otras. Entre los árboles, fue clara la predominancia de algunas sapotáceas, particularmente en el sector del campamento II. En términos generales, estos dos tipos de bosques parecieron altamente productivos.
En los bosques de colina alta con vegetación acahaparrada y colina media igualmente con vegetación achaparrada, la formación vegetal predominante fue semejante a un varillal seco y estuvo poblado por una mezcla de árboles y palmeras. Los árboles se caracterizaron por su porte bajo, entre 15 a 20 m de alto con algunos emergentes que no superaban los 25 metros, caducifolios y hojas xeromórficas; sotobosque en algunos casos denso y poblado por helechos y otras plantas herbáceas y en otros casi exclusivamente por palmeras de Attalea spp. Suelo arcillo arenoso a completamente arenoso, cubierto en la mayoría de las veces por un colchón de hojarascas. Este tipo de bosque nos pareció muy pobre en cuanto a producción de frutos.
En los bosques de colina media con vegetación alta, la formación vegetal predominante fue de tipo varillal alto poblado por árboles entre 20 a 25 m, con algunos emergentes hasta 30 m; la mayoría de ellos de fuste delgado y casi recto, en algunos casos bastante tupidos; sotobosque abierto mayormente poblado por Attalea spp., suelo de tipo areno-arcilloso y cubierto por hojarascas.
El aguajal de altura escasamente representado por una faja angosta entre los campamentos II y III, estuvo poblado por una mezcla de árboles y palmeras, con predominancia de Mauritia flexuosa L.f., Iriartea sp., Euterpe oleraceae Mart y Ficus spp.
El Bosque secundario estuvo presente únicamente en las antiguas trochas carrozables y viales para la extracción de madera. Aquí, la cobertura vegetal estuvo dominado mayormente por Cecropia spp., Guazuma sp., Muntingia sp. y Pourouma spp.
En términos generales los bosques en el área de estudio presentaron un aspecto ligeramente alterado, particularmente en el sector del campamento II. Asimismo, la fauna silvestre salvo excepciones nos pareció diversa y relativamente abundante.
Material y métodos
El inventario y evaluación de los primates se hicieron simultáneamente a los mamíferos mayores. Para este propósito hicimos uso de los nueve transectos abiertos en los alrededores de los tres campamentos y la trocha de acceso entre el campamento I al campamento III. Cada uno de los transectos medía entre 4 a 5 km de longitud e incluía diversos tipos de bosques y formaciones vegetales, diferenciados mediante el uso de mapas de imagen satelital y fisiográfico Datum WCS84-Proyección UTM18s. Los avistamientos incluyeron la visualización directa e indirecta (audición de vocalizaciones).
Los avistamientos fueron realizados durante las horas de mayor actividad de locomoción y alimentación, es decir, desde las 06:00 a 11:00 h y 14:00 a 17:00 h (hora local). El corto tiempo previsto para las evaluaciones en cada uno de los campamentos no permitió mayores exploraciones por los transectos y la trocha de acceso, limitándonos a un solo recorrido de ida y vuelta. Por otro lado, debido al agotamiento físico por la caminata del día no se hicieron avistamientos nocturnos. No obstante, el mono de habitos nocturnos Aotus nancymae Hershkovitz fue observado hasta en dos oportunidades en circunstancias diferentes. Las trochas fueron recorridas a una velocidad máxima de 1 km/ h, con paradas cada cierto trecho para auscultar desde el piso hasta lo alto del dosel con el objetivo de detectar cualquier movimiento o ruido. Cada vez que hubo avistamiento procedimos a anotar la especie, tipo de hábitat, número de individuos, distancia perpendicular de la trocha al primer animal avistado, altura en el estrato y la actividad al momento de su detección. Debido a la escasa cobertura de los censos, los avistamientos fueron bastante limitados por lo que no nos permitió estimar densidades, en su lugar, hemos desarrollado un criterio de abundancia (número de grupos e individuos por cada 100 km de longitud recorrida). Cuando los registros son insuficientes, este método de trabajo resulta apropiado y los resultados bastante confiables. Durante los censos fueron recorridos en total 114 km; de ellos, 46 km fueron cubiertos en el campamento I, 40 km en el campamento II y 28 km en el campamento III.
Resultados
Como resultado del inventario y la evaluación rápida logramos registrar 87 grupos de primates pertenecientes a 11 especies. De ellos, 82 fueron observados directamente, y solamente 5 a través de las vocalizaciones y correspondieron a Ateles chamek (un grupo) y Alouatta seniculus Linnaeus (cuatro grupos).
En el sector del Campamento I logramos observar 29 grupos que correspondieron a nueve especies, (Tabla 1).
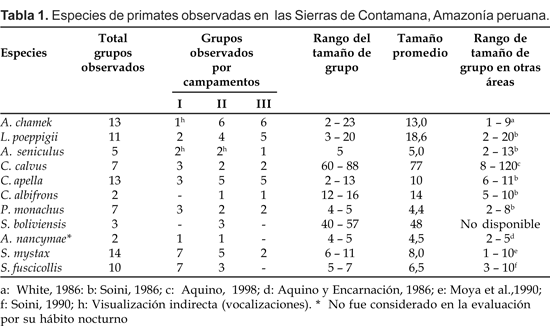
En este sector los primates más avistados fueron Saguinus mystax y Saguinus fuscicollis Spix, cuyos grupos en conjunto representaron cerca del 50% del total. No obstante, debemos resaltar la presencia de Ateles chamek (un grupo), Lagothrix poeppigii (dos grupos), Alouatta seniculus (dos grupos) y Cacajao calvus (tres grupos). En los registros se incluye un grupo de Aotus nancymae, observado circunstancialmente al oscurecer en momentos que se desplazaban a través del bosque secundario al borde de la trocha carrozable muy cerca del campamento I.
En el sector del Campamento II se observaron 34 grupos pertenecientes a 11 especies, constituyéndose así en el sector de mayor registro tanto de grupos como de especies (Tabla 1). A diferencia del anterior, los primates considerados como de tamaño grande fueron los más avistados, particularmente A. chamek, y L. poeppigii con seis y cuatro grupos, respectivamente. Entre los primates de tamaño pequeño, otro grupo de A. nancymae logramos observar en bosque de terraza alta en su respectivo árbol de dormir.
En el sector del Campamento III logramos avistar 24 grupos que correspondieron a ocho especies (Tabla 1), constituyéndose así en el sector de más bajo registro tanto en grupos como en especies. Sin embargo, como en el caso anterior los primates de tamaño grande como A. chamek (seis grupos), L. poeppigi (cinco grupos) y C. calvus (dos grupos) fueron los más avistados, que en conjunto representaron más del 50% de los avistamientos. Lo contrario ocurrió con las especies de tamaño pequeño que a diferencia de S. mystax, las demás estuvieron ausentes.
Con excepción de Alouatta seniculus y Saimiri boliviensis Hershkovitz, en las demás especies el tamaño promedio de grupo fue calculado de grupos donde los conteos fueron completos. El rango de variación en el tamaño de grupo y el tamaño promedio para las diferentes especies están contenidas en la Tabla 1. En A. seniculus, el tamaño promedio fue asumido del único grupo observado directamente; mientras que en S. boliviensis, el rango de variación y el tamaño promedio es un estimado, puesto que en ninguno de los tres grupos observados fue posible el conteo completo.
En el área de estudio las especies más abundantes fueron C. calvus, L. poeppigii y A. chamek (Tabla 2).
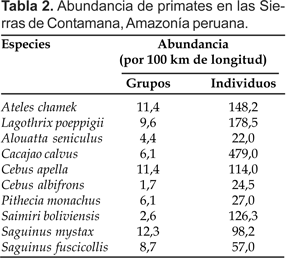
Entre los primates de tamaño grande, la única con menor abundancia fue A. seniculus, especie que tiene mayor predisposición por los bosques inundables antes que los bosques de altura.
Los primates fueron registrados haciendo uso de los diferentes tipos de bosques diferenciados para el área de estudio; sin embargo, los mayores avistamientos ocurrieron en los bosques de terraza alta (46%) y bosques de colina baja (27%) (Tabla 3).

Discusión
Especies no observadas
Además de las especies registradas, tomando en cuenta los antecedentes de colectas de pieles de mamíferos en esta parte de la Amazonía peruana y que datan desde los inicios del siglo XX; se esperaba el registro de Saimiri sciureus Linnaeus, Callimico goeldii Thomas y Callicebus cupreus Spix, pero tales acontecimientos no ocurrieron. En referencia a S. sciureus, la especie fue reportada para el área de Contamana Aguas Calientes y el sector Abujao Shesha (FPCN; CDC, 2001), Al respecto, Hershkovitz (1984) hace mención de los «Cerros de Contamana» como parte de su área de distribución geográfica. En cuanto a C. goeldii, existe el antecedente de colecta de un espécimen por L.W Hendei en 1927 en la Cordillera Azul, próximo a las cabeceras del río Contaya, afluente izquierdo del río Tapiche, entre las coordenadas 7º14' S y 74º34' W, y aproximadamente a 624 msnm (Hershkovitz, 1977). Por otro lado, en el Parque Nacional Serra do Divisor en Brasil, en el límite fronterizo con el Perú y un poco más al sur de las Sierras de Contamana recientemente fueron observados dos grupos de C. goeldii en asociación con Saguinus imperator (Aparecida de Oliveira et al., 2003). Para el caso de C. cupreus, también existen antecedentes de colecta en Cerro azul y Cashiboya (Hershkovitz, 1990). Recientemente para el área de Abujao Shesha hacen mención de la presencia de Callicebus sin especificar a la especie (FCN; CDC, 2001).
Considerando los antecedentes mencionados, es muy posible que estas especies están presentes en las Sierras de Contamana y la ausencia de registros durante el inventario por ahora asumimos que se debió al tiempo de permanencia que no fue lo suficiente como para indagar en otros hábitats y en alturas mayores a los 600 m, de modo que la confirmación solo podrá ser despejada en futuros inventarios y evaluaciones.
Tamaño de grupo
Salvo excepciones, el rango de variación del tamaño de grupo obtenido para las especies estuvo dentro o muy cercano de los estimados para otras áreas (Tabla 1). En A. chamek, los grupos en la mayoría de los casos fueron superiores a los 16 individuos, lo cual contrasta con los registrados para otras áreas de la Amazonía peruana. Al respecto, solamente en una oportunidad fue observado un grupo con 16 individuos en la Reserva Comunal Tamshiyacu Tahuayo (Aquino, obs. pers.), mientras que en el Parque Nacional Manú el máximo tamaño de grupo fue de nueve individuos (White, 1986), aún cuando en este último caso posiblemente se trataron de subgrupos, puesto que en los Ateles es común la formación de subgrupos e incluso el cambio en la composición del grupo a lo largo del día (Van Roosmalen, 1985; Mcfarland Symington, 1990; Chapman, 1990). En referencia a C. calvus, los grupos estuvieron compuestos por más de 60 individuos y como en el caso anterior podrían incluso haberse tratado de subgrupos, si consideramos los antecedentes de grupos de hasta 120 individuos para otras áreas (Aquino, 1998).
Hábitats preferenciales
La presencia y mayor abundancia de especies consideradas de tamaño grande es un buen indicador de la óptima calidad de un bosque primario como el de las Sierras de Contamana. Sin embargo, en el área hemos notado cierta predisposición de los primates y de otros componentes de la fauna por los bosques de terraza alta y colina baja, el cual podría estar en estrecha relación con la composición florística. En estos dos tipos de bosques fue notoria la existencia de una vegetación heterogénea donde los árboles eran tupidos, erguidos y con amplias copas, algunos de ellos incluso superaban los 35 m de alto. En ambos, la producción de frutos nos pareció alta en comparación a otros tipos de bosques, donde predominaron especies de las familias Sapotaceae y Arecaceae.
Especies amenazadas y no protegidas
De las 11 especies registradas habitando en las Sierras de Contamana, C. calvus, A. chamek, y L. poeppigii están clasificadas en situación vulnerable (VU) en la última lista de especies amenazadas para el Perú (D.S. No. 034-2004-AG); con posibilidades de incremento a cuatro si en el futuro se confirma la presencia de C. goeldii. De ellas, las dos primeras están contenidas en el CITES 2003 en los apéndices I y II, respectivamente. Además, en el área se encuentran habitando dos especies que aún no están protegidas en ninguna de las actuales áreas naturales protegidas del Perú. C. calvus con distribución en la margen derecha de los ríos Amazonas y Ucayali, desde la frontera con Colombia y Brasil en el norte hasta el río Sheshea en el sur (Aquino, 1988). Con excepción de la cuenca del río Yavarí, en el nor-oriente peruano esta especie es muy difícil de observar debido a la alta presión de caza al cual fue sometida en estos últimos años. Como consecuencia de la caza, los grupos en el nor-oriente, salvo excepciones no son muy grandes y oscilan entre 30 a 45 individuos (Aquino, 1998), lo cual contrasta con los observados en este estudio, donde el tamaño fluctuó entre 60 a más de 88 individuos. Su relativa mayor abundancia en esta parte de la Amazonía peruana podría tener su explicación en la baja o casi nula presión de caza y también quizá con factores ecológicos relacionado con los tipos de hábitats, recursos alimenticios y competidores, entre otros. S. mystax, con distribución similar a C. calvus es otra de las especies aún no protegidas; este primate es bastante común en esta parte de la Amazonía, posiblemente porque todavía no existe ningún tipo de presión de caza como viene ocurriendo en ciertos sectores cercanos a los grandes asentamientos humanos del nor-oriente peruano, donde incluso son capturados con trampa artesanal para el consumo de subsistencia, habiendo incluso ocasionado la extinción local en algunas cuencas cercanas a la ciudad de Iquitos.
Amenazas relevantes
Las amenazas para la supervivencia de las poblaciones de primates que habitan en las Sierras de Contamana por el momento son mínimas y están orientadas principalmente a la deforestación y la caza. En un enfoque global, el bosque primario se encuentra casi intacto; no obstante, hemos notado pequeñas alteraciones del bosque en el sector del campamento I como consecuencia de la agresiva extracción selectiva en el pasado de especies maderables, mientras que en el sector del campamento III encontramos un grupo de personas dedicados a la extracción de madera en tablones, cuyo impacto por el momento podríamos considerarlo como insignificante, pero que a mediano plazo y con el ingreso de un mayor número de madereros ilegales las poblaciones de primates y de otros componentes de la fauna silvestre podrían sufrir serias alteraciones, al extremo incluso de producirse fraccionamientos de sus hábitats y con ello el aislamiento de poblaciones como ya se advierten en otras partes de la Amazonía peruana. En cuanto a la caza, esta actividad mayormente está concentrada en el sector del campamento I, donde incluso contabilizamos hasta siete trochas en uso y en cada una de ellas fueron hallados varios casquillos de cartuchos, por lo que en este sector lo más probable es que existe alta presión de caza hacia los animales de mayor biomasa como los ungulados y primates de tamaño grande. Un buen indicador sería la escasa presencia de A. chamek, especie considerada como altamente susceptible a la presión de caza. En los otros dos sectores, las actividades de caza nos parecieron mínimas y muy ocasionales, por lo que en términos generales esta actividad por el momento no estaría afectando a las poblaciones de primates. Sin embargo, el impacto de la caza a mediano plazo podría ser mayor, particularmente en poblaciones de primates de tamaño grande, quienes han demostrado alta susceptibilidad a la caza.
Agradecimientos
Nuestro agradecimiento al Centro de Datos para la Conservación de la Universidad Nacional Agraria La Molina (CDC-UNALM), PRO-NATURALEZA y The Nature Conservancy (TNC) por habernos permitido participar en la Evaluación Ecológica Rápida (EER) en las Sierras de Contamana dentro del proyecto «Apoyo a la Conservación de la Sierra del Divisor», bajo el auspicio financiero de la Fundación Moore.
Literatura citada
Aparecida de Oliveira, M.; A. Lopez y J. A. Rehg. 2003. Observation of Callimico goeldii with Saguinus imperator in the Serra do Divisor National Park, Acre, Brasil. Neotrop. Primates 11 (3): 181 183. [ Links ]
Aquino, R. 1988. Preliminary survey on the population densities of Cacajao calvus ucayalii. Primate Conservation 9: 24 26.
Aquino, R. y F. Encarnación. 1990. Population structure of Aotus nancymai (Cebidae: Primates) in Peruvian amazon lowland forest. Am. J. Primatol. 11: 1 7.
Aquino, R. 1998. Some observation on the ecology of Cacajao calvus ucayalii in the Peruvian Amazon. Primate Conservation 18: 21 - 24.
Chapman, C. 1990. Association patterns of spider monkeys: the influence of ecology and sex on social organization. Behav. Ecol. Sociobiol. 26: 409 414.
FPCN; CDC. 2001. Evaluación ecológica rápida de las Sierras de Contamana y El Divisor. Lima. Informe final. 74 pag., más anexos.
Hershkovitz, P. 1977. Living new world monkeys (Platyrrhini), with a introduction to primates. Vol. 1. The University of Chicago Press, London. 1,117 pp.
Hershkovitz, P. 1984. Taxonomy of squirrel monkeys genus Saimiri (Cebidae, Platyrrhini): A preliminary report with descriptions of a hitherto unnamed form. Am. J. Primatol. 7: 155 210.
Herskovitz, P. 1990. Titis, new world monkeys of the genus Callicebus (Cebidae, Platyrrhini): A preliminary taxonomic review. Fieldiana, Zoology, New Series 55: 1 - 109.
Mcfarland Symington, M. 1990. Fission-fusion social organization in Ateles and Pan. Int. J. Primatol. 11 47 61.
Moya, L.; L. Verdi; G. Bocanegra y J. Rimachi. 1990. Análisis poblacional de Saguinus mystax (Spix 1823) (Callitrichidae) en la cuenca del río Yarapa, Loreto, Perú. En: La Primatología en el Perú.: Investigaciones primatológicas (1973 1985). Proyecto Peruano de Primatología (ed), pp . 80 -95. Lima, Perú.
Rodríguez, L. 1997. Diversidad biológica del Perú. Zonas prioritarias para su conservación, Proyecto FANPE,GTZ-INRENA. Lima, Industrial Payros S.A.
Soini, P. 1986. A synecological study of a primate community in the Pacaya Samiria National Reserve, Peru. Primate Conservation 7: 63 71.
Soini, P.; 1990. Ecología y dinámica poblacional de pichico común Saguinus fuscicollis (Callitrichidae, Primates). En: La Primatología en el Perú.: Investigaciones primatológicas (1973 1985). Proyecto Peruano de Primatología (ed), pp. 202 253. Lima, Perú
Van Roosmalen, M. G. M. 1985. Habitat preferences,diet,feeding strategy and social organization of the black spider monkey (Ateles p. paniscus Linnaeus 1758) in Surinam. Acta Amazonica 15 (supl.): 1 238.
White, F. 1986. Census and preliminary observations on the ecology of black-faced black spider monkey (Ateles paniscus chamek) in Manu National Park. Am. J. Primatol. 11: 125 132
Correspondencia:
1. Estación Experimental del IVITA-Iquitos, Perú. P.O. Box 575, Iquitos, Perú. Telefax 65-265510.
3. Fco. Tudela y Varela 215, Miraflores, Lima
E-mail Rolando Aquino: raquinoy2005@yahoo.es
