










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.14 n.1 Lima ago. 2007
recursos costeros y su manejo
Michelle I. Graco1,2, Jesús Ledesma2, Georgina Flores2 y Margarita Girón2
1 Dirección de Investigaciones Oceanográficas-proyecto CENSOR, Instituto del Mar del Perú, Callao, Perú.
2 Dirección de Investigaciones Oceanográficas- Unidad de Investigaciones en oceanografía Química, Instituto del Mar del Perú, Callao, Perú.
Marco de referencia
Una breve introducción al Sistema de Surgencias de la Corriente de Humboldt frente a Perú
El Sistema de surgencias de la corriente de Humboldt, es considerado entre los sistemas más productivos del mundo, extendiéndose a lo largo del pacífico sur oriental frente a Chile y Perú. En particular las aguas costeras frente a Perú atraen un considerable interés por encontrarse entre las más fértiles y productivas del mundo, destacándose pesquerías, como la de anchoveta, que representan uno de los pilares de la economía del Perú.
Frente a Perú, la producción primaria alcanza valores entre 3 y 4 g C m-2 d-1 en la franja costera de 100 km (Calienes et al., 1985; Chavez et al., 1989). La alta productividad resulta de la gran disponibilidad de nutrientes y de factores forzantes favorables que se mantienen durante todo el año (Pocklington, 1981). Igualmente ciertas áreas altamente productivas coinciden con la presencia de una extensa plataforma, particularmente entre los 7º-10º S (Zuta & Guillén, 1970).
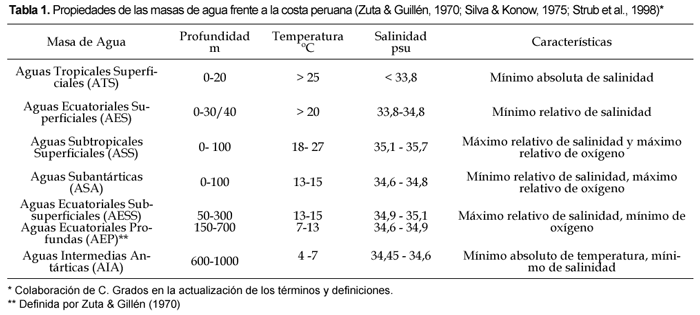
La disponibilidad de nutrientes frente a Perú es el resultado de eventos de surgencias costera, los cuales transportan desde profundidades someras (usualmente 50_100 m) aguas con un alto contenido de nitratos y bajo contenido de oxígeno disuelto (Zuta & Guillén, 1970; Pocklington, 1981). Importantes centros de surgencias se han identificado frente a Perú, a los 5º S (Paita), 6º S (Punta aguja), 9º S (Chimbote), 12º S (Callao) y 15º S (San Juan) (Zuta & Guillén, 1970; Rojas De Mendiola, 1981). Las surgencias pueden alimentarse de diferentes masas de agua (Zuta & Guillén, 1970): aguas relativamente oxigenadas de La Extensión Sur de la Corriente Subsuperficial de Cromwell que afloran al norte de la latitud 6º S; aguas deficientes en oxígeno de la Corriente Peruana Subsuperficial que va hacia el polo aflorando al norte de los 12º S y en ocasiones un poco más al sur, y finalmente de la mezcla de Aguas Templadas Subantárticas y Aguas Ecuatoriales Subsuperficiales que alimentan los afloramientos al sur de los 14º S (Zuta & Guillén, 1970).
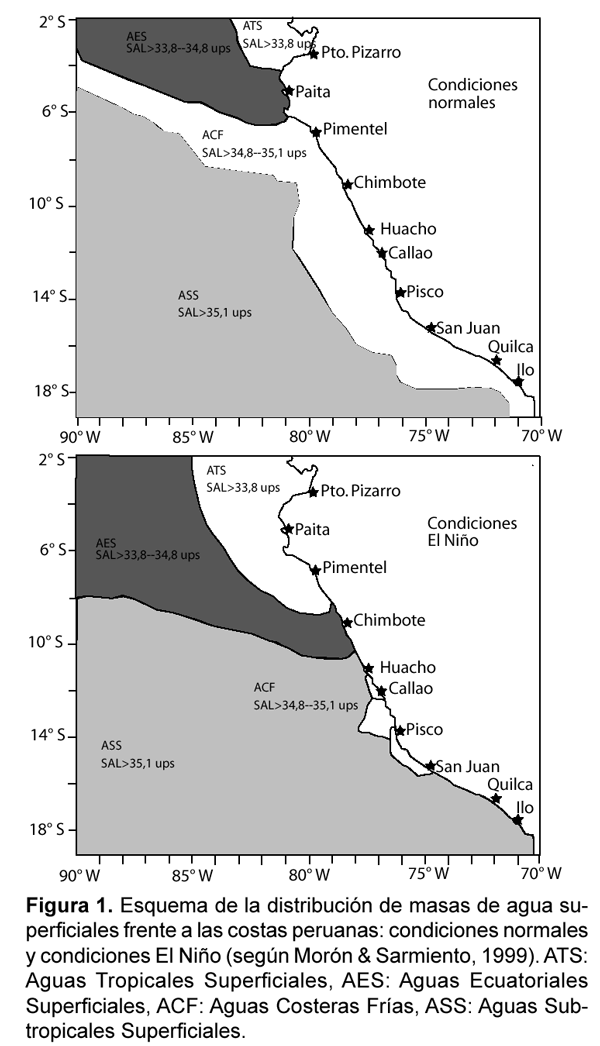
Aunque los eventos de surgencia ocurren durante todo el año frente a Perú, existe una alta variabilidad oceanográfica de origen local y/o remoto a diferentes escalas temporales (intraestacional, estacional, interanual y decadal). Esta variabilidad oceanográfica determina importantes fluctuaciones en el Frente Ecuatorial, en la Extensión Sur de la Corriente de Cromwell y en el afloramiento costero afectando la abundancia y distribución de los recursos biológicos. Algunos aspectos de la variabilidad estacional oceanográfica han sido discutidos por Zuta & Guillén (1970) y Wooster & Gilmartin (1961), donde se reconoce que a lo largo de la costa de Perú existe una mayor intensidad en la surgencia durante invierno y primavera, siendo más débil en verano y otoño. Igualmente en la región se observa una de las señales más intensas asociadas con el ciclo El Niño Oscilación del Sur (ENOS). Durante la fase cálida del ENOS, conocida como El Niño (EN), la distribución y extensión de las masas de agua varía (Tabla 1, Fig. 1, a y b), con desplazamientos hacia la costa de Aguas Subtropicales Superficiales y desplazamiento hacia el sur de Aguas Ecuatoriales Superficiales y Aguas Tropicales Superficiales las cuales se caracterizan por un bajo contenido de nutrientes. A nivel subsuperficial se intensifica temporalmente la Corriente Peruana Subsuperficial, elevándose la concentración de oxígeno disuelto (Guillén et al., 1985; Strub et al., 1998). Se reconocen igualmente los eventos La Niña (LN), sin embargo estos han sido menos descritos. Estos eventos presentan características opuestas a la de los Niños, observándose una intensificación en las aguas de afloramiento frías, ricas en nutrientes y una alta productividad (Chavez, 2005).
La gran producción biológica y la variabilidad de los procesos bio-oceanográficos del sistema de surgencias de la corriente de Humboldt frente a Perú han sido objeto de numerosos estudios y campañas oceanográficas tanto de carácter nacional como internacional. Algunas de estas campañas realizadas por el Instituto del Mar del Perú IMARPE (e.j., cruceros oceanográficos, cruceros de evaluación hidroacústica de recursos pelágicos, entre otros) se han mantenido a lo largo del tiempo con el fin de evaluar la abundancia y distribución de los recursos marinos y su interrelación con el ambiente en el cual se desarrollan, siendo este último aspecto clave para la comprensión del ecosistema y su funcionamiento.
El objetivo de este trabajo es presentar una revisión de las investigaciones en oceanografía química realizadas a lo largo de la costa de Perú para finalmente resaltar los desafíos futuros existentes en el área.
Revisión de las características químicas del sistema de surgencias de la corriente de Humboldt frente a Perú
Investigaciones y campañas oceanográficas en Perú
Diversos estudios y campañas oceanográficas, nacionales e internacionales, se han realizado a lo largo de la costa peruana y han permitido avanzar en el conocimiento de la oceanografía química del área. Entre los primeros trabajos oceanográficos realizados en Perú, en el año 1936 aparece un informe sobre el sistema de surgencias de la corriente de Humboldt frente a Perú en el cual se describe su productividad, distribución y variabilidad de nutrientes, particularmente fosfatos (Gunther, 1936). A fines de los años 50 y comienzos de los años 60, se realiza una importante campaña oceanográfica extranjera (STEP I) con el objeto de caracterizar la Contracorriente Subsuperficial de Perú-Chile (Wooster & Cromwell, 1958; Wooster, 1961; Wooster & Gilmartin, 1961; Wooster et al., 1965). Estos trabajos presentan mediciones de silicatos, nitratos y nitritos. La primera revisión de las condiciones químicas de las aguas costeras frente a Perú es el trabajo realizado por Zuta y Guillén (1970), en el cual se describe la distribución y variabilidad de las propiedades físicas, nutrientes y oxígeno a partir de datos obtenidos en diferentes cruceros realizados entre los años 1928 y 1967 (e.j., Carnegie VII, Antón Bruun, Crucero Unitas III) y entre los años 1961 y 1968 (e.j., "Bondy", "Unanue"). Entre los años 1976 y 1977 se realizó un importante esfuerzo de colección de datos en el marco del programa internacional CUEA "Coastal Upwelling Ecosystems Analysis". Este programa tuvo por objeto el estudio de aspectos físicos, químicos y biológicos de los ecosistemas de surgencias costeras (e.j., California, Óregon, NO de África y Perú). En el caso de Perú se realizaron expediciones (JOINT-II), a los 15º S (marzo-octubre, 1976 y marzo-mayo, 1977) que generaron diversas publicaciones (Fiadeiro & Strickland, 1968; Dugdale et al., 1977; Packard et al., 1978; Codispoti & Packard, 1980; Codispoti, 1981; Friederich & Codispoti, 1981; Guillén & Calienes, 1981; Packard et al., 1983; Codispoti & Christensen, 1985).
Una segunda revisión de los nutrientes, clorofila y productividad primaria en el área, aparece en los años 80, enfocada en la variabilidad espacio-temporal de las aguas costeras frente a Perú (Calienes et al., 1985). La serie de datos analizados en esta revisión corresponde al período 1964 y 1978, obtenidos en cruceros bio-oceanográficos periódicos nacionales asociados con el estudio del fenómeno del Niño (EUREKA Y EBCCEN) y con cruceros del programa CUEA. En la década de los 80 se realizaron varias campañas extranjeras, tal como NITROP-85 y PACIPROD, la primera asociada al estudio del ciclo del nitrógeno y la mínima de oxígeno (Codispoti et al., 1986; Codispoti et al., 1989; Ward et al., 1989a; Ward et al., 1989b; Lipschultz et al., 1990) y la segunda en la productividad y las surgencias frente a Perú (Minas et al., 1990; Copin-Montégut & Raimbault, 1994).
A nivel nacional, en el año 1986 IMARPE inicia un importante sistema de observación bio-oceanográfico mensual a lo largo de la costa de Perú conocido como Monitoreo Oceanográfico Pesquero en Áreas Seleccionadas o MOPAS. Estas observaciones, entre las que se encuentran mediciones de nutrientes y oxígeno en la columna de agua, se realizaron mensualmente en el área de Callao, Etapa I (Zuta, 1989) para luego extenderse al área norte como parte de las actividades del proyecto especial, Estudio Integral del fenómeno El Niño (PEEICON), y finalmente a las áreas de Tumbes, Paita, Chimbote e Ilo. Datos de las variables físicas y químicas obtenidas durante los MOPAS (años 1986 y 1991) se presentan en el atlas de R. Calienes (1992).
En los últimos años los estudios en oceanografía química se han enfocado en la comprensión de la productividad (Chavez, 1995; Bruland et al., 2005) y la abundancia y distribución de los recursos marinos (IMARPE, Cruceros Oceanográficos Regionales, Cruceros de estimación de la biomasa desovante de anchoveta etc), siendo el componente químico uno de los aspectos analizados (Ledesma & Sarmiento, 2000; Morón & Ledesma, 2001; Flores et al., 2004).
A continuación se realizará una síntesis de los principales hallazgos en oceanografía química para el Sistema de surgencias de la corriente de Humboldt frente a Perú a partir de los trabajos realizados y publicados en el marco de las expediciones y estudios expuestos anteriormente.
Distribución y variabilidad de Nutrientes
Los sistemas de surgencias se caracterizan por una alta concentración de nutrientes inorgánicos, tales como nitratos (NO3-), fosfatos (PO43-) y silicatos (SiO24+) los cuales se encuentran en exceso con respecto a las concentraciones requeridas para ser incorporados por los organismos. La concentración de estos nutrientes es uno de los factores que afecta el crecimiento de la comunidad marina, pero también lo son su distribución y variabilidad, objeto de estudio de la mayoría de los trabajos en el área.
Los rangos promedio superficiales de nutrientes publicados para la costa de Perú son: 0,2_4,0 μM PO42-; 0,0_35,0 μM NO3-; 0,0 _ 30,0 μM SiO24+ (Zuta & Guillén, 1970; Guillén & Izaguirre De Rondán, 1973; Calienes et al., 1985).
-Variabilidad costa-océano y latitudinal
Las mayores concentraciones de nutrientes frente a Perú se localizan cerca de la costa. Las altas concentraciones de nutrientes son resultado de lo aportado por las aguas de surgencias, el transporte de corrientes y la ocurrencia de procesos biogeoquímicos de regeneración y/o consumo de nutrientes en la capa eufótica o en el sedimento. Además el área se caracteriza por la presencia de altas concentraciones de nutrientes en forma de "lenguas" o plumas que se observan mar afuera a lo largo de la costa peruana (Calienes et al., 1985). Estas lenguas de aguas frías se explican por la dinámica oceanográfica del área, asociándolas con estructuras tipo remolinos (ciclónicas y anticiclónicas) y filamentos, limitadas horizontalmente a 50 km o menos (Strub et al., 1998). En el caso de los nitratos hasta las 50 millas de la costa presentan valores entre 20,0 μM y 0,5 μM , disminuyendo a casi cero en aguas oceánicas. Al igual que los silicatos en aguas recientemente advectadas a la superficie su concentración es alta, sin embargo los florecimientos fitoplanctónicos pueden agotarlos totalmente (Dugdale & Goering, 1967; Zuta & Guillén, 1970). Experimentos realizados durante los cruceros CUEA (1976-77), sobre consumo y regeneración de silicatos, muestran que la regeneración de este nutriente en las capas superficiales es mínima, siendo el primer nutriente que se agota, pudiendo ser limitantes para el fitoplancton (Nelson et al., 1981). En el caso de los nitratos, en aguas oceánicas se han reportado condiciones de altos nitratos y baja clorofila. Este exceso de nitrato con respecto a la asimilación por parte el fitoplancton ha sido explicado por efecto de pastoreo sobre el fitoplancton (Walsh, 1976). Recientemente investigaciones conducidas en aguas peruanas muestran que a lo largo de la costa sur de Perú y aguas afuera existen condiciones pobres en hierro bio-disponible, lo cual podría limitar la productividad a pesar de existir altas concentraciones de nitratos (Hutchins et al., 2002; Bruland et al., 2005).
Con respecto a la variabilidad latitudinal muchos trabajos indican que los nutrientes decrecen de norte a sur (Zuta & Guillén, 1970; Guillén & Izaguirre De Rondán, 1973; Codispoti, 1981). Al norte de los 15º S se destacan mayores concentraciones de nitratos (hasta 35,0 μM) y silicatos (hasta 30,0) que al sur (Guillén & Izaguirre De Rondán, 1973).
La distribución latitudinal de nutrientes está asociada con los distintos tipos de masas de aguas envueltos en el régimen de circulación del área. Los fosfatos y silicatos presentan valores más altos (> 2,5 mM y > 20,0 μM respectivamente) asociados con centros activos de surgencias, y los más bajos (< 1,0 μM y < 4,0 μM respectivamente) con Aguas Ecuatoriales Superficiales y Aguas Subtropicales Superficiales características del área norte (Zuta & Guillén, 1970; Guillén & Izaguirre De Rondán, 1973; Calienes & Guillén, 1981). La figura 2 representa la distribución promedio de nutrientes durante el verano y el invierno frente a la costa de Perú según Calienes et al.(1985). Esta figura indica, al igual que los trabajos precedentes, que las mayores concentraciones de nitratos, silicatos y fosfatos se asocian con los principales centros de surgencias, e igualmente sugieren la existencia de una variabilidad latitudinal. Esta variabilidad latitudinal es evidente en el caso de los nitratos que registran mayores concentraciones al norte de los 12º S (Fig. 2). Bajas concentraciones para los tres nutrientes se observan en el norte (> 4º S).
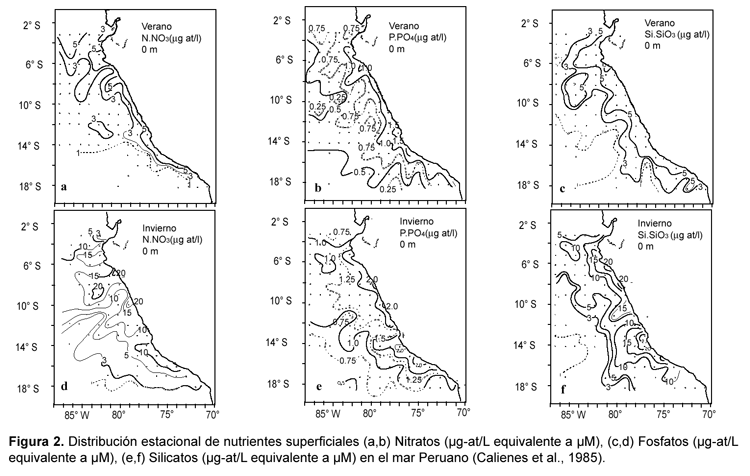
Con respecto a las diferencias latitudinales en la concentración de nitratos una de las explicaciones, tal como veremos en las secciones siguientes, podría asociarse con la existencia de procesos de reducción de nitrato en condiciones deficientes en oxígeno, condiciones que se intensifican al sur de los 10º S (Wooster et al., 1965; Fiadeiro & Strickland, 1968; Ward et al., 1989a).
-Distribución vertical de nutrientes en la columna de agua
La distribución vertical de fosfatos, silicatos y nitratos frente a las costas de Perú (Fig. 3) se caracteriza por una capa superficial, una capa de transición, una capa intermedia y una capa profunda (Guillén, 1966; Zuta & Guillén, 1970; Strickland & Parsons, 1972; Guillén & Izaguirre De Rondán, 1973).
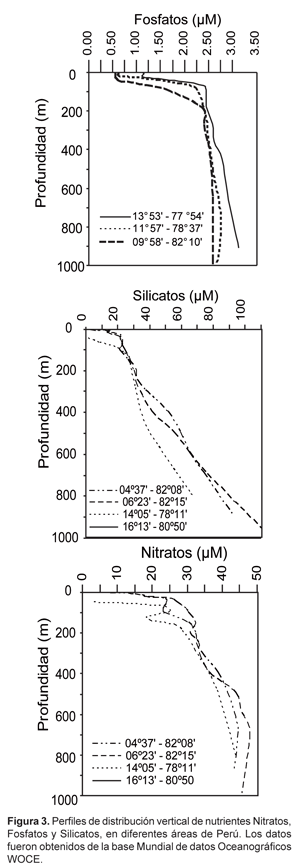
La capa superficial, de aproximadamente 50 m, presenta valores más bajos y relativamente uniformes con respecto a las capas más profundas (0,25 -1,2 μM PO43-; < 15,0 μM SiO24+ y < 15,0 μM NO3-). Mientras los nitratos y silicatos pueden agotarse durante los florecimientos fitoplanctónicos, el fósforo está presente siempre en exceso.
En la capa de transición, que coincide con la Termoclina Subsuperficial Permanente (TSP) y se encuentra por encima de los 100 m, encontramos un aumento rápido en los fosfatos de 1,0 a 2,0 μM y ocasionalmente de 0,5 a 2,5 μM. En el caso de los silicatos esta capa presenta valores entre 5,0 y 30,0 μM. Para los nitratos, esta capa está ligeramente sobre la capa de transición de silicatos, con concentraciones de 5,0 a 20,0 μM intensificándose en la costa.
La capa intermedia, es la capa en que los fosfatos alcanzan la máxima concentración con valores de 2,5 μM como se observa en los perfiles (Fig. 3) e incluso de 3,5 μM entre las profundidades de 500 y 1,200 m, por debajo de esnte se reducen. Valores superiores a 3,0 μM ocurren hacia la costa, y en las aguas de fondo en contacto con la plataforma continental. Para los silicatos en la capa intermedia tenemos valores entre 30,0 y 50,0 μM que se extienden hasta los 500 m y una capa más profunda con valores superiores a los 50 μM y de hasta 110 μM (Fig. 3). Los nitratos por su parte presentan por encima de los 600 m concentraciones de 20,0-45,0 μM y por debajo de los 600 m hasta aproximadamente los 1,300 m valores de nitratos de 40,0-55,0 μM.
La distribución vertical de los nutrientes resulta de los procesos físicos y biológicos que están ocurriendo, tales como circulación, presencia de diferentes masas de agua, eventos de surgencia, procesos biológicos (incorporación por fitoplancton, excreción) y biogeoquímicos (nitrato-reducción, remineralización). La variabilidad de estos procesos físicos, químicos y biológicos resulta en diferencias en los perfiles tales como los que se observan en la fig. 4 (secciones a diferentes latitudes de Perú). La capa superficial de la columna de agua, por ejemplo, se caracteriza por presentar máximos y mínimos que se asocian con los eventos de surgencias (aguas afloradas ricas en nutrientes) y/o la actividad de fotosíntesis (incorporación de nutrientes). En el caso de las capas intermedias, los altos nitratos evidencian el fuerte efecto de las Aguas Ecuatoriales Subsuperficiales o Aguas Ecuatoriales Profundas, particularmente entre 100 y 300 m (Zuta & Guillén, 1970). La disminución subsuperficial de los nitratos se asocia con procesos biogeoquímicos que se explican en detalle en las secciones siguientes. En el caso de los silicatos y fosfatos, los mayores valores que ocurren en aguas de fondo y en contacto con la plataforma se asocian con el transporte lateral, la interacción con el bentos y las altas tasas de remineralización de materia orgánica que allí ocurren.
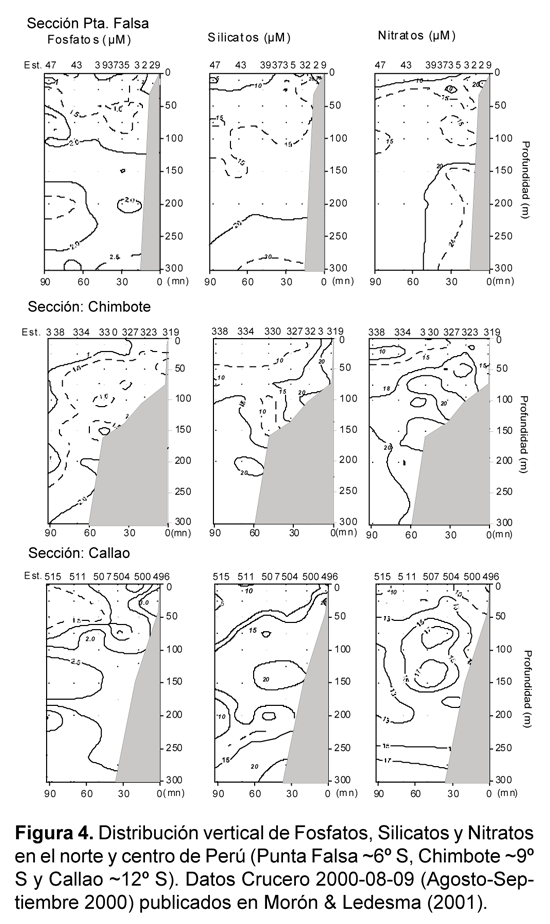
-Variabilidad temporal de nutrientes
Los procesos biogeoquímicos junto con la dinámica oceanográfica del área determinan no solo los cambios espaciales sino también la variabilidad temporal de los nutrientes, identificándose diferentes escalas de variabilidad temporal, tales como la intraestacional, estacional, interanual y decadal.
Pocos trabajos han sido publicados en relación a la variabilidad temporal, la mayoría de ellos asociados a la variabilidad estacional (Zuta & Guillén, 1970; Guillén et al., 1977; Calienes et al., 1985). En estos trabajos se analiza la variabilidad superficial de los nutrientes en relación a la dinámica oceanográfica y la distribución de masas de agua tal como muestra la fig. 2. En el caso de los fosfatos se observa que valores máximos (2,0 μM) ocurren en invierno y más bajos en verano (hasta de 0,25 μM). En verano la isolínea de fosfatos de 1,0 μM se extiende cerca a la costa, mientras en invierno se aleja, encontrándose concentraciones de 1,5 μM que se distribuyen más allá de las 200 millas (Fig. 2 c y d). En el área central de Perú (frente a Callao ~ 12º S), los cambios se producen por encima de los 50 m, la isolínea de fosfatos de 2,5 μM presenta grandes fluctuaciones estacionales, con un mayor acercamiento a la superficie en julio, septiembre y diciembre, y un mayor hundimiento en enero, febrero y de octubre a noviembre (Calienes et al., 1985). Igualmente en el norte (Punta falsa, ~ 6º S) se producen fluctuaciones en la isolínea de 2,0 μM con hundimiento en presencia de aguas de mayor salinidad y elevaciones en presencia de aguas con salinidades menores a 35,00 ups (Zuta & Guillén, 1970).
Los silicatos y los nitratos presentan distribuciones y variabilidad similares. Las mayores concentraciones de silicatos (25,0 μM) se observan en invierno y primavera y las menores en verano (1,0 μM, Fig. 2 e y f), con máximas concentraciones presentes en el área sur (~15º S) (Calienes et al., 1985). La variación de los silicatos está muy asociada con la variabilidad de las fases activas y pasivas de la surgencia y de los tipos de poblaciones fitoplanctónicas presentes (Zuta & Guillén, 1970). En el caso de los nitratos los valores máximos ocurren en invierno (20,0 μM, Fig. 2 a y b), mientras durante el verano los valores son próximos a cero (Calienes et al., 1985). La variabilidad de los nutrientes tiene un patrón temporal similar al de la temperatura, mostrando el rango anual de la concentración de nutrientes a una temperatura dada, límites definidos para cada estación del año. En resumen podemos decir que la variabilidad estacional de los nutrientes se asocia con la variabilidad en la intensidad de los eventos de surgencia, máximos en invierno y primavera, y más débiles en verano y otoño (Calienes et al., 1985).
Además de los cambios estacionales, hay cambios en la distribución de nutrientes influenciados por eventos como El Niño (EN) o La Niña (LN). Durante El Niño se ha reconocido que las aguas superficiales frente a Perú son alteradas por el flujo de aguas anormalmente cálidas y pobres en nutrientes que provienen del Ecuador. En estos períodos se observa el desplazamiento hacia el sur de Aguas Tropicales Superficiales (ATS) y Aguas Ecuatoriales Superficiales (AES) y hacia la costa de las Aguas Subtropicales Superficiales (ASS) (Fig. 1 b). Durante EN intenso el desplazamiento de las ATS y AES pueden llegar a la zona central de Perú (Guillén et al., 1985). Durante EN la termoclina y la nutriclina se profundizan, los eventos de surgencia se desarrollan en áreas muy reducidas y aunque los vientos favorables a las surgencias continúan, las aguas advectadas a la superficie presentan valores de nutrientes inferiores a los considerados en promedio para la región (Zuta & Guillén, 1970).
Durante EN de 1972-73 se detectaron altas concentraciones de silicatos cerca de la costa, asociadas a una pobre productividad (Rojas De Mendiola, 1969). Estudios realizados durante 1987 muestran como las Aguas Costeras Frías fueron desplazadas por aguas calientes. Durante el verano las AES caracterizadas por una baja concentración de nutrientes (0,5-0,7 μM PO43-; 10,7 _ 0,8 μM NO3- y 4,0 _ 5,0 μM SiO24+) se extienden hasta los 8º30' S. Igualmente frente a los 7º_8º S aparecen las ASS que también poseen un contenido bajo de nutrientes (<0,5 μM PO43-, <5 μM NO3- y SiO24+) (Guillén et al., 1989).
Con respecto a la distribución de nitratos, la capa de 5_10 μM normalmente asociada a la nutriclina, resulta útil para seguir el desarrollo de los eventos EN. Durante EN 1982 y 1987 esta capa se profundizó hasta los 80 m comparado con lo observado en períodos neutros del ciclo ENOS en que se presenta más superficial (Calienes et al., 1985; Guillén et al., 1985; Guillén et al., 1989). Durante EN el nitrato aparece como el factor limitante para la productividad, mientras el silicato es limitante durante los períodos neutros (Guillén & Izaguirre De Rondán, 1973).
Durante EN 1997-98 las concentraciones de nutrientes, nitratos, silicatos y fosfatos decrecieron significativamente, particularmente entre los meses de primavera y verano con respecto a los valores que se observaron durante años que no presentaron condición EN (Purca, 2005; Flores et al., 2006; Graco et al., 2006) tal como lo ejemplifica la figura 5 para los fosfatos y silicatos.
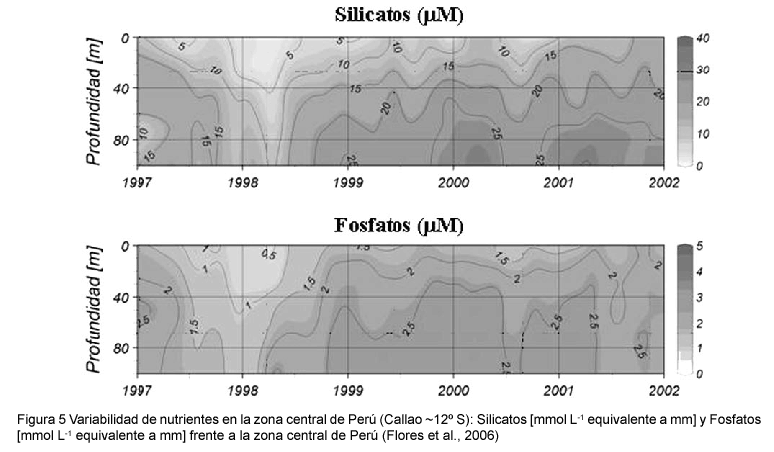
Los cambios decadales son cada vez más reconocidos como fuentes importantes de variabilidad en la estructura atmosférica y oceánica (Miller et al., 1994; Field & Baumgartner, 2000). La variabilidad decadal puede modular la frecuencia, intensidad y ocurrencia del ciclo ENOS (Wang & Picaut, 2004). Respecto al sistema de surgencias de la Corriente de Humboldt, se ha postulado para los últimos 45 años la ocurrencia de un "régimen de anchoveta" o "La Vieja", desde el inicio de las pesquerías hasta inicios de los 70's, y desde el inicio de los 90's hasta la actualidad; separados por un "régimen de sardina" o "El Viejo" desde mediados de los 70's hasta inicios de los 90's (Chavez et al., 2003; Alheit & Ñiquen, 2004). El Viejo es comparable en sus manifestaciones frente al Pacífico oriental a un Niño persistente en la escala decadal, afectando la temperatura superficial del mar y la profundidad de la termoclina. Durante `La Vieja' en cambio la termoclina es más somera, y en consecuencia se espera un mayor aporte de nutrientes e incremento de la producción primaria (Chavez et al., 2003; Chavez, 2005). En el caso de los nutrientes aun no se ha explorado este tipo de variabilidad, siendo necesario el análisis retrospectivo de la información histórica de la región.
-Relación entre nutrientes
Además de la concentración y distribución de los nutrientes en el agua de mar, es importante la relación que existe entre ellos, tal como la proporción entre Nitrógeno/Fósforo/Sílice (N/P/Si). El fitoplancton, considerado el más abundante de los productores marinos, constituye una fuente dominante de materia orgánica, con una proporción de Nitrógeno/Fósforo de 16/1 (proporción Redfield C/N/P 106/16/1). Las desviaciones de estas proporciones pueden tener diferentes orígenes, una materia orgánica con una proporción diferente de N/P, procesos de remineralización distintos de los aeróbicos, y/o la presencia de nutrientes preformados que caracterizan una masa de agua y pueden acumularse durante su transporte (Libes, 1992). En consecuencia, la proporción entre nutrientes es un indicador de procesos físicos y biogeoquímicos que están ocurriendo en el ambiente con implicancias en las comunidades fitoplanctónicas que se desarrollan (diatomeas versus dinoflagelados) y en la productividad del sistema.
Para el Océano Pacífico, Fiadeiro & Strickland (1968) encontraron una proporción atómica entre el N/P de 13/1 en aguas de fondo y algo menor en aguas someras. Frente a Perú Strickland (1970) encontró una relación N/P/Si de 19,2/2,6/35,0 y a los 1000 m de 18,5/2,9/35,0. En el trabajo de Guillén y colaboradores (1977) observaron valores de N/P/Si de 11,0/1,0/9,0 en la capa de 0 _ 25 m dentro de las 50 millas de la costa y más allá de las 55 millas de 13,0/1,0/9,0. Los mayores valores de N/P (> 14,0) se asocian con aguas transportadas por la Contra Corriente Ecuatorial Sur, y los menores (< 10,0) con las Aguas Subantártica que fluyen hacia el norte. Valores más bajos (< 8,0) se encuentran al sur del área de surgencia de los 15º S.
En la superficie de la Corriente Costera Peruana la proporción de N/P es en promedio de 9,0 (21,5/0,2) (Guillén & Izaguirre De Rondán, 1973), similar a la proporción encontrada frente a California, otra área caracterizada por eventos de surgencia (Park, 1967). En el caso de N/Si las aguas superficiales peruanas muestran una proporción de 1,7 (Guillén & Izaguirre De Rondán, 1973; Guillén et al., 1977), y de 1,0 (Calienes et al., 1985). Con respecto a la relación entre Si/P no es lineal, y la proporción se incrementa con la profundidad.
La proporción entre nutrientes indica que entre fosfatos, nitrato y silicatos, los silicatos parecen ser el nutriente limitante para el crecimiento fitoplanctónico. Los nitratos, presentan proporciones bajas con respecto a los fosfatos, lo cual evidencia no sólo la incorporación por fitoplancton sino además la existencia de una significativa remoción de nitratos en el ambiente.
Oxígeno y Zonas de Mínimo de Oxígeno frente a Perú
-Distribución y variabilidad del oxígeno disuelto
La distribución y concentración de oxígeno en los sistemas de surgencias presentan dos particularidades, las capas superficiales muestran altas concentraciones de oxígeno, mientras las subsuperficiales y profundas pueden tener valores cercanos a cero (Fig. 6 a). En efecto, frente a las costas peruanas las condiciones de oxígeno en las aguas superficiales son elevadas, con valores máximos en verano y otoño. Valores mayores a 7 mL L-1 se encuentran en las aguas frías de latitudes altas y menores a 5 mL L-1 en las regiones calientes próximas al ecuador (Zuta & Guillén, 1970). Se puede decir que la concentración de oxígeno aumenta al alejarse de la costa, valores mínimos (hasta 2 mL L-1) se encuentran en las áreas de afloramiento, mientras los máximos ocurren en las áreas de intensa fotosíntesis. Condiciones de sub-saturación de oxígeno se han observado superficialmente, lo que podría indicar una actividad fotosintética reducida y/o un mínimo de oxigenación biológica (Minas et al., 1990).
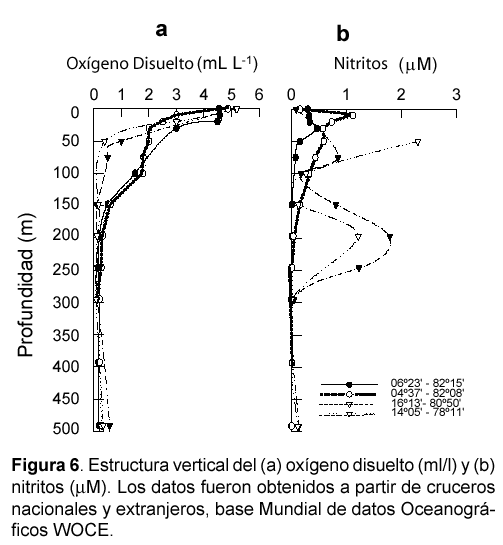
Zuta y Guillén (1970) describen en detalle la distribución vertical del oxígeno frente a Perú (Fig. 6 a). Existe una capa superficial de alto oxígeno por debajo de la cual aparece una capa de discontinuidad, conocida como oxiclina, en la cual el oxígeno cae bruscamente hasta el valor de 1,0 mL L-1 e incluso más bajo. Esta capa persiste durante todo el año, y se relaciona en el norte con la termoclina superficial permanente, y al sur de los 6º S con la termoclina subsuperficial permanente. Generalmente al sur de los 14º S se asocia además con la haloclina. Por debajo de la oxiclina se encuentra una capa de concentraciones bajas de oxígeno (< 0,5 mL L-1, Fig. 6). Esta capa limita la conocida Zona de Mínima de Oxígeno (ZMO), la cual describiremos en detalle en la sección siguiente. Por debajo de la ZMO y en relación con las Aguas Antárticas Intermedias (> 500m), el oxígeno incrementa sus concentraciones.
-Zona de Mínimo de Oxígeno (ZMO) frente a Perú
Las ZMOs, son zonas del océano persistentes que se distribuyen a profundidades intermedias y se caracterizan por presentar condiciones de muy bajo oxígeno (< 0,5 mL L-1) (Anderson et al., 1982; Kamykowski & Zentara, 1990). Estas ZMOs se desarrollan cuando hay aguas pobres en oxígeno y una alta demanda de oxígeno en combinación con una pobre ventilación (Wyrtki, 1962; Kamykowski & Zentara, 1990). Un largo período de residencia y una intensa actividad biogeoquímica determinan en estas áreas una alta tasa de consumo de oxígeno, llegando a valores cercanos a cero. La importancia de cada uno de los procesos de ventilación, circulación y productividad en la formación, mantenimiento y evolución de las ZMOs está en debate (Minas et al., 1990; Pennington et al., In press) y es objeto de estudio de diferentes grupos de investigación.
Una ZMO bien desarrollada se localiza frente a Perú donde comparada con otros sistemas es mucho más intensa y somera (Helly & Levin, 2004). A partir de estudios, y datos de oxígeno obtenidos en los diferentes cruceros y actividades realizadas frente a Perú, tanto a nivel nacional (IMARPE) como internacional (Wooster & Cromwell, 1958; Wooster & Gilmartin, 1961; Zuta & Guillén, 1970; Codispoti & Packard, 1980) se observa que:
• La capa deficiente en oxígeno (<0,5 mL L-1), conocida como ZMO, se encuentra a partir de los 30-50 m intersectando la capa eufótica y siendo una de las ZMO más someras del planeta (Helly & Levin, 2004).
• La figura 6 muestra la distribución de la ZMO con una mínima absoluta o núcleo localizado entre los 50 y 450 _ 500 m con valores predominantemente menores a 0,25 mL L-1. La isolínea de 0,25 mL L-1 de oxígeno aparece más superficial cerca de la costa y hacia el sur. En el norte (5_6º S) se presenta por debajo de los 100 _ 200 m, mientras a partir de los 12º S predomina por debajo de los 50 m. Valores menores a 0,1 mL L-1 aparecen a los 15º S a partir de los 150 _ 200 m cerca de la costa y se profundiza mar adentro (>300 m) (Minas et al., 1990).
• La ZMO se asocia con la contracorriente de Chile-Perú y las AESS o Aguas Ecuatoriales Subsuperficiales definidas en el Pacífico Oriental Sur frente a Chile (Morales et al., 1999) y denominadas para Perú como AEP o Aguas Ecuatoriales Profundas por Zuta y Guillén (1970). Estas masas de agua pobres en oxígeno se caracterizan además por altas salinidades, bajas temperaturas y altos nitratos.
Con respecto a la variabilidad de la ZMO en el sistema de surgencias de la corriente de Humboldt, aún se conoce muy poco. Existen algunas observaciones a escala estacional, sin embargo la mayor parte de los estudios de la ZMO se han enfocado principalmente en los cambios que ocurren durante el ciclo ENOS. En el Pacífico Oriental Sur frente a Chile se ha observado durante eventos El Niño (EN) una significativa profundización del límite superior de la mínima de oxígeno, incrementando en la costa la concentración de oxígeno en los 100 m indicando su alejamiento de la costa (Morales et al., 1999). Trabajos realizados frente a Perú, en la zona de Paita (5º S) y San José (6º 45' S) muestran como la distribución de la mínima de oxígeno es modulada por los eventos EN o La Niña (LN), los cuales modifican la intensidad de la Corriente Peruana Subsuperficial y aumentan el oxígeno en la columna de agua (Guillén et al., 1985). Por ejemplo, durante EN 1997_1998, la zona norte de Perú mostró la depresión en la isoterma de 15º C por debajo de los 100 m y la intrusión de AES, oxigenándose gran parte de la columna de agua (Gutiérrez et al., 2005; Ledesma et al., 2006). La zona central de Perú (12º S), igualmente mostró cambios en la ZMO durante EN 1997_1998, tal como lo muestra la figura 7 a y b, profundizándose y distribuyéndose más allá de los 100 m y observándose aguas costeras más oxígenadas a diferencia de lo observado durante períodos no El Niño en que estas aguas se distribuyen a partir de los 30_50 m (Graco et al., 2006).
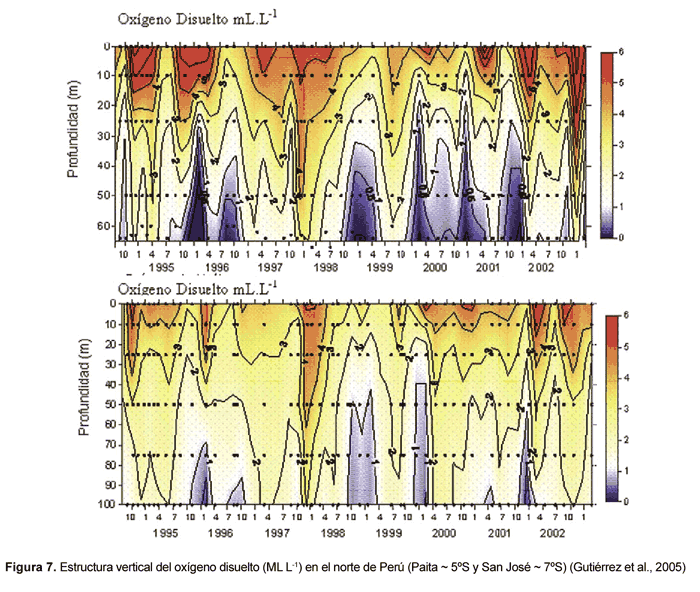
Durante LN 1998-99, por su parte, en la región norte de Perú la columna de agua se caracterizó por la dominancia de aguas frías y pobres en oxígeno, manifestándose una mínima de oxígeno significativamente más somera que en períodos neutros (Gutiérrez et al., 2005; Ledesma et al., 2006). Existen trabajos recientes que muestran cambios en la ZMO, particularmente cambios en su borde superior y en la intensidad de las concentraciones de oxígeno frente a Perú a escalas estacional e intraestacional asociada a forzamiento remoto (Graco et al., 2006; Gutiérrez et al., 2006).
Con respecto a los cambios latitudinales en la distribución de la ZMO, las condiciones de mínima de oxígeno se encuentran a partir de los 2_3º S (Wooster & Gilmartin, 1961). La mínima de oxígeno en el norte y hasta lo 9_11º S de Perú, se profundiza a unos 100_200 m (Fig. 6 a), por efecto del control que ejerce el flujo de aguas oxigenadas cercanas a la costa asociadas a la extensión Sur de la Corriente de Cromwell, mientras hacia el Sur se superficializa y se intensifica (Fiadeiro & Strickland, 1968; Codispoti & Packard, 1980).
Implicancias biogeoquímicas de la ZMO frente a Perú
La presencia de una ZMO tiene un impacto significativo desde el punto de vista ambiental, ecológico y económico. En efecto, estas áreas constituyen ambientes muy particulares donde se desarrollan comunidades peculiares tanto pelágicas como bentónicas. Estudios en el norte de Chile muestran la ocurrencia de una comunidad de cianobacterias y bacterias en condiciones de bajo oxígeno (Ulloa, com. pers.). En el caso del bentos la coexistencia de bajo oxígeno y altos nitratos favorece el desarrollo de tapices de bacterias gigantes, e.j., Thioploca, cuyas biomasas pueden superar las de otros organismos bentónicos (Jørgensen & Gallardo, 1999). La coexistencia de bajo oxígeno y alta materia orgánica también modula la estructura y abundancia de las comunidades bentónicas asociadas (Arntz et al., 1991; Gutiérrez et al., 2005; Gutiérrez et al., 2006).
Desde el punto de vista biogeoquímico existen algunos trabajos en el Pacífico Sur Oriental que evalúan el impacto de estas áreas en el ciclo del carbono, en las tasas de remineralización e intercambio de CO2 (Paulmier, 2005; Chavez, 2005), y en el ciclo del nitrógeno (Brandhorst, 1959; Fiadeiro & Strickland, 1968; Cline & Richards, 1972; Codispoti & Richards, 1976). Asociado con el ciclo del nitrógeno en ZMO procesos como la desnitrificación y nitrato reducción aparecen como claves siendo los nitritos un importante trazador en estas áreas, tal como se expondrá a continuación.
-ZMO y máximos de nitritos
Los nitritos (NO2-) representan un estado de oxidación intermedio entre el amonio (NH4+) y el nitrato (NO3-), y su presencia refleja un estado inestable en el sistema. Los nitritos pueden ser producidos y/o excretados por el fitoplancton, bacterias yo utilizados por diferentes microorganismos a diferentes profundidades. En la mayor parte de las regiones del océano abierto, concentraciones muy bajas de nitrito se detectan, tanto en las aguas superficiales como en las aguas profundas. Sin embargo en los sistemas de surgencias, y en particular frente a Perú una de las peculiaridades que se observa es la presencia de altas concentraciones subsuperficiales de nitritos.
Dos máximos de nitritos que se observan en la columna de agua, uno que se encuentra cerca del piso de la capa superficial y se denomina máximo primario de nitritos (MPN), y otro por debajo del MPN denominado máximo secundario de nitritos (MSN) (Fig. 6). Este último se observó por primera vez en el año 1937 (Gilson, 1937) en el mar de Arabia, con concentraciones de hasta 7,4 μM. Posteriormente, Brandhorst (1959) lo reconoció en el Pacífico Tropical, México, Norte de Chile y Perú.
Frente a Perú, al norte de los 6º S, tal como muestra la Fig. 6 b, aparece sólo el MPN ubicándose en la base de la termoclina superficial estacional (~50 m), entre los 10 y 60 m de profundidad y asociado con las aguas templadas subantárticas (Fiadeiro & Strickland, 1968). Este MPN presenta valores promedio de 3,5 _ 0,1 μM. Al sur de los 10_12º S y distribuyéndose hasta el norte de Chile (25º S) se han observado los dos máximos, el MPN y el MSN (Fig. 6 b). El MPN entre 10 y 50 m y El MSN que se extiende verticalmente entre los 50 _ 100 m cerca de la costa, y a más de 200 m a aproximadamente 10 millas. El MSN se asocia con las Aguas Ecuatoriales Subsuperficiales y presenta concentraciones entre 1,0 y 5,5 μM (Wooster et al., 1965; Zuta & Guillén, 1970), con un máximo de hasta 9,0 μM mar afuera entre los 12º y 15º S (Codispoti & Packard, 1980). A los 15º S el MPN ocurre entre los 20 y 50 m, y el MSN se presenta entre los 75 y 350 m (Packard et al., 1978).
El MSN se observa frecuentemente asociado a concentraciones de oxígeno inferiores a 0,2 mL L-1 (Fiadeiro & Strickland, 1968), es decir con la mínima de oxígeno absoluta tal como lo muestra la figura 6 a y b. Sin embargo, a pesar de que las concentraciones bajas de oxígeno aparecen como un prerrequisito para la presencia de esta acumulación de nitritos, su distribución no se extiende más allá de los 9º S, a pesar de que mínimas de oxígeno se encuentran hasta los 2_3º S (Wooster & Gilmartin, 1961). El límite norte en la distribución del MSN puede ser explicado por el control que ejerce el flujo de aguas oxigenadas (i.e., Cromwell) (Fiadeiro & Strickland, 1968; Codispoti & Packard, 1980) que determina la profundización de la mínima de oxígeno a unos 100 _ 200 m figura 6 a y b
El límite sur del MSN está dado por la posición de la corriente oceánica de Perú que se aleja de la costa a los 25º S. Las altas concentraciones de nitrito presentes en el sur de Perú y norte de Chile pueden asociarse al transporte y flujo subsuperficial hacia el polo de la Contracorriente de Chile-Perú (Codispoti & Packard, 1980).
-Origen de los máximos de nitritos
El MPN se ha atribuido a la oxidación de amonio durante la descomposición de la materia orgánica, y también a la liberación extracelular de nitrito durante la asimilación de nitrato por el fitoplancton (Richards, 1965; Spencer, 1975). La falta de requerimientos para la fotosíntesis (e.j., luz, niveles de ferredoxina) y exceso de nitratos determina que el fitoplancton excrete nitrito y cese el crecimiento (Morris, 1974).
En el caso del MSN, la acumulación de nitritos que se observa se asocia con el proceso de reducción de nitratos, que es el primer paso de la desnitrificación y que permite la transformación de nitratos en nitrógeno gaseoso. El proceso de reducción de nitratos puede ser inhibido por la presencia de oxígeno, limitado por la disponibilidad de nitrato y en mayor proporción por la disponibilidad de bacterias desnitrificantes (Payne, 1973). Frente a Perú una importante actividad de nitrato reductasa se encontró en el núcleo del MSN, coincidente con un déficit de nitrato de 7,0 μM y condiciones de oxígeno menores a 0,2 mL L-1 (Packard et al., 1978). Un importante hallazgo en la ZMO frente a Perú, fue la existencia de actividad nitrificante (oxidación de amonio) en condiciones deficientes en oxígeno (< 2,0 mL L-1) considerado un dominio de las bacterias desnitrificantes(Ward et al., 1989a; Ward et al., 1989b). En consecuencia los nitritos podrían acumularse por procesos de oxidación de amonio, siendo máxima esta actividad en los bordes de la ZMO y mínima en el núcleo. Losa procesos de nitrificación y desnitrificación parecen coexisten en el área existiendo un importante acoplamiento entre ellos, dando lugar a la pérdida de nitrógeno pero también pudiendo contribuir con acumulación de nitritos.
-Variabilidad temporal de las concentraciones máximas de nitrito
Pocos estudios existen sobre la variabilidad temporal de los nitritos. Se ha observado que los bordes espaciales pueden ser desplazados temporalmente durante eventos EN. Posterior al evento EN de 1972 la zona costera presentó concentraciones de nitritos mayores a 7,0 μM, incremento que podría asociarse con cambios en procesos de desnitrificación (Codispoti & Christensen, 1985; Codispoti et al., 1986). Durante las fases frías igualmente se han observado ciertas anomalías. Durante 1974-75 entre los 4º y 10º S, el MSN se observó a una profundidad de 100 m, y concentraciones de hasta 11,0 μM en condiciones ambientales bien reducidas, con una alta producción de sulfuros (HS2) (Dugdale et al., 1977). Esta condición puede considerarse transitoria y fue observada mar adentro en los 50 m, coincidiendo con una zona de remoción completa de nitratos en prácticamente toda la columna de agua. Igualmente, luego de EN de 1982-1983, concentraciones de nitritos mayores a 1,0 μM fueron encontradas frente a Perú coincidiendo con la distribución de la ZMO. Entre los 7º y 10º S fueron inusualmente altos los nitritos, registrándose concentraciones de hasta 23,0 μM. este incremento en nitrito sugiere una situación fría posterior al evento cálido y un nuevo incremento en las tasas de desnitrificación (Codispoti et al., 1986).
Estudios recientes muestran significativos cambios interanuales en las concentraciones de nitritos, tales como los observados durante EN 1997-1998 los cuales se correlacionan con cambios en la distribución vertical de la ZMO, particularmente la posición de su borde superior (Graco et al., 2006).
Conclusiones
Los diferentes estudios realizados por investigadores nacionales y extranjeros en el Sistema de Surgencias de la Corriente de Humboldt frente a Perú han contribuido sustancialmente en la comprensión de los procesos de la surgencia costera. Desde el punto de vista de la oceanografía química y biogeoquímica han permitido reconocer un sistema caracterizado por una alta variabilidad espacial y temporal en sus parámetros hidroquímicos tales como nutrientes (nitratos, fosfatos y silicatos) presentes en altas concentraciones en el área. Además se reconoce en el área la presencia de una Zona de Mínima de Oxígeno (ZMO) caracterizada por aguas deficientes en oxígeno (< 0,5 mL L-1) que se distribuyen según la latitud a partir de los 50 o 200 m, y en ocasiones interceptan la plataforma. Las condiciones de altos nutrientes, alta productividad y bajo oxígeno por su parte favorecen en el área procesos como la nitrato reducción el cual se evidencia a partir de la acumulación de nitritos subsuperficiales y por la pérdida de nitrógeno generando importantes déficit en los nitratos y modificaciones en las proporciones de C/N/P en el área. Los diversos cruceros y monitoreos bio-oceanográficos realizados frente a Perú han permitido describir algunas escalas de variabilidad oceanográfica existente. Se observa la importancia de la dinámica oceanográfica (distribución de masas de agua) y la intensidad de los eventos de surgencias a escala estacional, como también la influencia del ciclo ENOS, con sus fases cálidas (EN) y frías (LN) a escala interanual. Se reconoce la existencia de otras escalas de variabilidad (intraestacionales-ondas atrapadas e interdecadales), sin embargo aún no se ha profundizado en estos aspectos. Los estudios realizados en el área han abordado diferentes temáticas además del estudio de los parámetros hidroquímicos, como son el CO2; hierro y productividad, ciclo del nitrógeno, abriendo numerosos interrogantes que constituyen un desafío para esta área de investigación.
Las características hidroquímicas del área de surgencias de la corriente de Humboldt frente a Perú y su variabilidad son aspectos claves para comprender las fluctuaciones en la productividad, en la distribución y abundancia de los recursos marinos y en general de los componentes biológicos del sistema. El desafío actual es avanzar en la comprensión de uno de los ecosistemas más productivos del mundo, estudiando el ambiente químico, buscando determinar el tipo de relaciones que existen entre las condiciones químicas, tales como nutrientes u oxígeno y los distintos componentes biológicos del sistema. En este sentido, la profundización en los análisis retrospectivos de datos químicos es un aspecto clave, como también el análisis integrado y multidisciplinario de datos y procesos físicos, químicos y biológicos.
Agradecimientos
Agradecemos los valiosos comentarios y correcciones del Dr. D. Gutiérrez. Un reconocimiento especial a los técnicos Miguel Sarmiento y Carlos Robles de la Unidad de Investigaciones en Oceanografía Química (IMARPE), y a todos aquellos cuya labor a través de los años nos permite avanzar en el conocimiento de la oceanografía química. Agradecemos al Instituto del Mar del Perú (IMARPE). Este trabajo fue realizado en el marco del proyecto europeo CENSOR (Climate variability and El Nino Southern Oscillation: Impacts for natural resources and management, contract 511071) y es la publicacion CENSOR 0089.
Literatura citada
Alheit, J. & C. Ñiquen. 2004. Regime shifts in the Humboldt Current Ecosystem. Progress in Oceanography (60): 201-222.
Anderson, J.J.; A. Okubo; A.S. Robbins & A. Richards. 1982. A model for nitrite and nitrates distributions in oceanic oxygen minimum zones. Deep-Sea Research (29): 1113-1140.
Arntz, W.; J. Tarazona; V.A. Gallardo; L.A. Flores & H. Salzwedel. 1991. Benthos communities in oxygen deficient shelf and upper slope areas of the Peruvian and Chilean pacific coast, and changes caused by El Niño. Geological Society: 131-154.
Brandhorst, W. 1959. Nitrification and denitrification in the eastern tropical North Pacific. J. Cons. int. Explor. Mer (25): 3-20.
Bruland, K.W.; E.L. Rue; G.J. Smith & G.R. Ditullio. 2005. Iron, macronutrients and diatom blooms in the Perú upwelling regime: brown and blue waters of Perú. Marine Chemistry (93): 81-103.
Calienes, R. & O. Guillén. 1981. Masas de agua y producción primaria en el Perú. Bol. Inst. Mar Perú (Vol. Extraordinario ICANE): 155-163.
Calienes, R.; O. Guillén & N. Lostaunau. 1985. Variabilidad espacio-temporal de clorofila, produccion primaria y nutrientes frente a la costa peruana. Bol. Inst. Mar Perú (10): 6-12.
Calienes, R. 1992. Proyecto MOPAS: Monitoreo Oceanográfico pesquero en áreas seleccionadas. Inf. Inst. Mar Perú (102): 167.
Chavez, F.P.; R.T. Barber & M.P. Sanderson. 1989. The potential Primary Production of the Peruvian Upwelling System. Instituto del Mar del Peru (IMARPE), Deutshe Gesellschafr fur Technische Zusammenarbeit (GTZ) and International Center for Living Aquatic Resources Mangement (ICLARM).
Chavez, F.P. 1995. A comparison of ship and satellite chlorophyll from California and Peru. Journal of Geophysical Research (100): 24,855 - 824, 862.
Chavez, F.P.; J. Ryan; S.E. Liuch-Cota & C. Ñiquen. 2003. Climate, fish, ocean productivity, and atmospheric carbon dioxide. Science (299): 217-221.
Chavez, F.P. 2005. Biological consequences of interannual to multidecadal variability. En A. Robinson & K. Brink [eds.], The Sea. Harvard University Press, Texas A. & M.Cambridge. p. 643-679.
Cline, J.D. & A. Richards. 1972. Oxygen deficient conditions and nitrate reduction in the eastern tropical North Pacific Ocean. Limnology and Oceanography (17): 885-900.
Codispoti, L.A. & A. Richards. 1976. An analysis of the horizontal regime of denitrification in the eastern tropical North Pacific. Limnology and Oceanography (21 (3)): 379-388.
Codispoti, L.A. & T.T. Packard. 1980. Denitrification rates in the eastern tropical South Pacific. Journal of Marine Research (38): 453-477.
Codispoti, L.A. 1981. Temporal nutrient variability in three different upwelling regions. En F. A. Richards [ed.], Coastal Upwelling. Coastal and estuarine Science 1. American Geophysical Union.Washington. p. 209-220.
Codispoti, L.A. & J.P. Christensen. 1985. Nitrification, denitrification and nitrous oxide cycling in the eastern tropical south Pacific Ocean. Marine Chemistry (16): 277-300.
Codispoti, L.A.; T.T. Friederich; T.T. Packard; H.E. Glover; P.J. Kelly; R.W. Spinrad; R.T. Barber; W. Elkins; B.B. Ward; F. Lipschultz & N. Lostanau. 1986. High nitrite levels off northern Perú. A signal of inestability in the marine denitrification rate. Science (233): 1200-1202.
Codispoti, L.A.; R.T. Barber & G.E. Friederich. 1989. Do nitrogen transformation in the poleward undercurrent off Perú and Chile a globally significant influence? En R. L. Smith & R. T. Barber [eds.], Coastal and estuarine studies. Poleward flows along Eastern Ocean Boundaries. Coastal and Estuarine Studies. Springerp. 280-310.
Copin-Montégut, C. & P. Raimbault. 1994. The Peruvian upwelling near 15o C in August 1986. Results of continuous meausurements of physical and chamical properties between 0 and 200 m depth. Deep-Sea Research Part I (41): 439-467.
Dugdale, J.J. & J.J. Goering. 1967. Uptake of new and regenerated forms of nitrogen in primary productivity. Limnol. Oceanogr. (12): 196-206.
Dugdale, R.C.; J.J. Goering; R.T. Barber; R.L. Smith & T.T. Packard. 1977. Denitrification and hydrogen sulfide in the Peru upwelling region during 1976. Deep-Sea Res. (24): 601-608.
Fiadeiro, M. & J.D. Strickland. 1968. Nitrate reduction and the ocurrence of a deep nitrite maximum in the ocean off the west coast of South America. Journal of Marine Research (26): 187-201.
Field, D.B. & T.R. Baumgartner. 2000. A 900 year stable isotope record of inter-decadal and centennial change from the California Current. Paleoceanography (15): 695-708.
Flores, G.; C. Robles & M. Sarmiento. 2004. Condiciones hidroquímicas frente a la costa norte-centro del Perú durante agosto-septiembre 2001. Inf. Inst. Mar Perú (32): 127-134.
Flores, G.; M. Graco; M. Bouchon; E. Delgado; M. Girón; J. Ledesma & L. Pizarro. 2006. Impact of the oceanographic variability in the phytoplankton community and the anchovy vertical distribution in front of Callao (12º S) Perú during 1997-2000. Extended Abstracts of the International Conference The Humboldt Current System Conference: Climate, ocean dynamics ecosystem processes and fisheries. Lima, Peru. November 27-December 1. p. 135-136.
Friederich, G.E. & L.A. Codispoti. 1981. The effects of mixing and regeneration on the nutrient content of upwelling waters off Perú. En F. A. Richards [ed.], Coastal Upwelling. Coastal and estuarine sciences 1. American Geophysical Union.Washington, D.C., p. 221-227.
Gilson, H.C. 1937. The nitrogen cycle. Chem. Phys. Invest. (Sci. Rep. Murray exped., 2): 21-81.
Graco, M.; G. Flores; J. Ledesma; S. Purca; D. Gutiérrez & M. Girón. 2006. Biogeochemical variability in the oxygen minimum zone of the upwelling Humboldt system off central Perú. Extended Abstracts of the International Conference The Humboldt Current System Conference: Climate, ocean dynamics ecosystem processes and fisheries. Lima, Peru. November 27-December 1. p. 32-33.
Guillén & R. Calienes. 1981. Upwelling off Chimbote. En F. A. Richards [ed.], Coastal Upwelling. Coastal and estuarine sciences 1. American Geophysical Union.Washington, D.C., p. 312-326.
Guillén, O. 1966. Variación de los fosfatos en el Callao como medida de la producción primaria., Memoria el I Seminario Latino-Americano sobre el océano pacífico oriental. p. 192-198.
Guillén, O. & R. Izaguirre De Rondán. 1973. Nutrients in the Perú coastal current. En R. Fraser [ed.], Oceanography of the South Pacific 1972. National comission for UNESCO.Wellington. New Zeland. p. 397-418.
Guillén, O.; R. Calienes & R. Rondan De. 1977. Medio ambiente y producción primaria frente al área Pimentel-Chimbote. Bol. Inst. Mar Perú (3): 107-159.
Guillén, O.G.; E.A. Cárcamo & R. Calienes. 1989. Oxígeno disuelto, nutrientes y clorofila frente a la costa peruana durante el Niño 1987. Pacífico Sur (Número Especial): 83-94.
Guillén, O.N.; N. Lostaunau & M. Jacinto. 1985. Características de El Fenómeno El Niño 1982-1983. En W. Arnzt; A. Landa &J. Tarazona [eds.], El Niño y su impacto en la fauna marina. Bol. Inst. Mar. Perú-Callao. p. 11-20.
Gunther, E.R. 1936. A report on oceanographical investigations i the Perú coastal current. Discovery Report (13): 107-276.
Gutiérrez, D.; K. Aronés; F. Chang; L. Quipúzcoa & P. Villanueva. 2005. Impacto de la variación oceanográfica estacional e inter-anual sobre los ensambles de microfitoplancton, mesozooplancton, ictioplancton y macrozoobentos de dos áreas costeras del norte del Perú entre 1994 y 2002. Bol. Inst. Mar Perú (22): 3-60.
Gutiérrez, D.; E. Enríquez; S. Purca; J. Pasapera; L. Quipúzcoa; G. Flores & M. Graco. 2006. Remotely-driven temporal variations of the subsurface dissolved oxygen content and responses of the continental shelf benthic subsystem off central Perú. Extended Abstracts of the International Conference The Humboldt Current System Conference: Climate, ocean dynamics ecosystem processes and fisheries. Lima, Perú. November 27- December 1. p. 206-208.
Helly, J.J. & L.A. Levin. 2004. Global distribution of naturally ocurring marine hypoxia on continental margins. Deep-Sea Research Part I (51): 1159-1168.
Hutchins, D.A.; C.E. Hare; R.S. Weaver; Y. Zhang; G.F. Firme; G.R. Ditullio; M.B. Alm; S.F. Riseman; J.M. Maucher; M. Geesey; et al. 2002. Phytoplankton iron limitation in the Humboldt Current and Perú upwelling. Limnol. Oceanogr. (47): 997-1011.
Jørgensen, B.B. & V.A. Gallardo. 1999. Thioploca spp.: Filamentous sulfur bacteria with nitrate vacuoles. FEMS Microbiology Ecology (28): 301-313.
Kamykowski, D. & S.-J. Zentara. 1990. Hypoxia in the world ocean as recorded in the historical data set. Deep-Sea Research (37): 1861-1874.
Ledesma, J. & M. Sarmiento. 2000. Condiciones hidroquímicas del mar peruano a fines del invierno 1999, en un área seleccionada (7º - 9ºS). Inf. Inst. Mar Perú (154): 65 - 76.
Ledesma, J.; G. Flores & M. Graco. 2006. Chemical Shift characteristics in the sea Along the peruvian Coast, during El Niño 1997-98. Extended Abstracts of the International Conference The Humboldt Current System Conference: Climate, ocean dynamics ecosystem processes and fisheries. Lima, Perú. November 27- December 1. p.
Libes, S.M. 1992. An introduction to Marine Biogeochemistry. John Wiley & Sons, Inc., Nueva York, 734 p.
Lipschultz, F.; S.C. Wofsy; B.B. Ward; L.A. Codispoti; G. Friedrich & W. Elkins. 1990. Bacterial transformation of inorganic nitrogen in the oxygen-deficient waters of the eastern tropical south Pacific ocean. Deep-Sea Research II (37): 1513-1541.
Miller, A.; D.R. Cayan; T.P. Barnett; N.E. Graham & J. Oberhunter. 1994. Interdecadal variability of the pacific Ocean: model response to observed heat flux and wind stress anomalies. Climate dynamics (9): 287-302.
Minas, H.J.; B. Coste; M. Minas & P. Raimbault. 1990. Conditions hydrologiques, chimiques et production primaire dans les upwellings du Pérou et des iles Galapagos, en régime d`hiver austral (capagne Paciprod). Oceanologica Acta (10): 383-391.
Morales, C.; S. Hormazabal & J.L. Blanco. 1999. Interannual variability in the mesoscale distribution of the depth of the upper boundary of the oxygen minimum layer off northen Chile (18-24S): Implications for the pelagic system and biogeochemical cycling. Journal of Marine Research (57): 909-932.
Morón, O. & M. Sarmiento. 1999. Detección de especies e invertebrados indicadores del fenómeno El Niño y otros trastornos con el medio ambiente. Internal report. IMARPE. p. 6
Morón, O. & J. Ledesma. 2001. Aspectos químicos del mar peruano en agosto y setiembre 2000. Inf. Inst. Mar Perú (162): 43 - 50.
Morris, I. 1974. Nitrogen assimilation and protein synthesis. En W. D. P. Stewart [ed.], Algal Physiology and Biochemistry. University of California Press.Berkeley. p. 583-609.
Nelson, D.M.; J.J. Goering & D.W. Boisseau. 1981. Consumption and regeneration of Silicic Acid in three coastal upwelling systems. En F. A. Richards [ed.], Coastal Upwelling. Coastal and estuarine Science 1. American Geophysical Union.Washington. p. 242-256.
Packard, T.T.; J.J. Dugdale; J.J. Goering & R.T. Barber. 1978. Nitrate reductase activity in the subsurface waters of the Perú current. Journal of Marine Research (36): 59-76.
Packard, T.T.; P.C. Garfield & L.A. Codispoti. 1983. Oxygen consumption and denitrification below the peruvian upwelling. En E. Suess & J. Thiede [eds.], Coastal Upwelling. Plenum Press.NY., p. 147-173.
Park, K. 1967. Nutrient regeneration and preformed nutrients off Oregon. Limnol. Oceanogr. (12): 353-357.
Paulmier, A. 2005. Zones de minimum d`Oxygene (OMZs) de l`océan moderne. PhD. Pierre et marie Curie (Paris VI).Paris. pp 248.
Payne, W.J. 1973. Reduction of nitrogenous oxides by microorganisms. Bacteriological Reviews (37): 409-452.
Pennington, J.T.; K.L. Mahoney; V.S. Kuwahara; D.D. Kolber; R. Calienes & F.P. Chavez. In press. Primary Production in the Eastern Tropical Pacific: A Review. Progress In Oceanography, Volume 69, Issues 2-4, May-June 2006, Pp 285-317.
Pocklington, R. 1981. Organic matter in upwelling off Northern Perú, November 1977. Bol. Inst. Mar Perú (Vol. extraordinario ICANE): 7-14.
Purca, S. 2005. Variabilidad temporal de baja frecuencia en el Ecosistema de la Corriente Humboldt frente a Perú. Tesis de Doctorado.Universidad de Concepción.Concepción, Chile. 32 pp. Anexos 37pp.
Richards, F.A. 1965. Anoxic basins and fjords. En J. P. Riley & G. Skirrow [eds.], Chemical Oceanography, Vol 1. Academic Pressp. 611-645.
Rojas De Mendiola, B. 1969. Contenido estomacal de anchoveta en cuatro áreas de la costa peruana., Informe 27. Inst. Mar Perú. 31 p.
Rojas De Mendiola, B. 1981. Seasonal phytoplankton distribution along the peruvian coast. En F. A. Richards [ed.], Coastal Upwelling. Coastal and estuarine Science 1. American Geophysical Union.Washington. p. 348-356.
Silva, N. & D. Konow. 1975. Contribución al conocimiento de las masas de agua en el Pacífico Sudoriental. Expedición Krill. Crucero 3-4, Julio-Agosto 1974. Revista de la Comisión Permanente del Pacífico Sur (3): 63-75.
Spencer, C.P. 1975. The micronutrient elements. En J. P. Riley & G. Skirrow [eds.], Chemical Oceanography. Academic Press.New York. p. 245-300.
Strickland, J. & T. Parsons. 1972. A practical handbook of sea water analysis. Fisheries Research Board of Canada. 310 pp.
Strickland, J.D. 1970. Part IV Research on the marine planktonic food web at the institute of marine resources., IMR Report 70-5 p.
Strub, P.T.; J.M. Mesías; V. Montecino; J. Rutlant & S. Salinas. 1998. Coastal ocean circulation off western South America. Coastal Segment (6, E). En A. R. Robinson & K. H. Brink [eds.], The Sea. Wiley.New York. p. 273-313.
Walsh, J.J. 1976. Herbivory as a factor in patterns of nutrients utilization in the sea. Limnol. Oceanogr. (21): 1-13.
Wang, C. & J. Picaut. 2004. Understanding ENSO physics- A review. En S. Wang; P. Xie &J. A. Carton [eds.], Earth`s Climate: The Ocean-Atmosphere Interaction. AGU Geophysical Monograp Seriesp. 1-19 p.
Ward, B.B.; H.E. Glover & F. Lipschultz. 1989a. Chemoautotrophic activity and nitrification in the oxygen minimum zone off Perú. Deep sea research (36): 1031-1051.
Ward, B.B.; K.A. Kilpatrick; E. Renger & R.W. Eppley. 1989b. Biological nitrogen cycling in the nitracline. Limnology and Oceanography (34): 493-513.
Wooster, W.S. & T. Cromwell. 1958. An oceanographic descripcion of the eastern tropical Pacific. Bull. Scripss Instn. Oceanogr. (7): 169-282.
Wooster, W.S. 1961. Step I Expedition, 15 September to 14 December 1960. Univ. Calif. Scripps Inst. Oceanogr. Rep. 61-9, 50 pp. p. 50
Wooster, W.S. & M. Gilmartin. 1961. The Peru-Chile Undercurrent. Journal of Marine Research (19): 97-122.
Wooster, W.S.; T.J. Chow & I. Barret. 1965. Nitrite distribution in the Perú current Waters. Journal of Marine Research (23): 210-221.
Wyrtki, K. 1962. The oxygen minima relation to ocean circulation. Deep sea research (9): 11-23.
Zuta, S. & O.G. Guillén. 1970. Oceanografía de las aguas costeras del Perú. Bol. Inst. Mar Perú (2): 157-324.
Zuta, S. 1989. Sumario de las condiciones dominantes en el verano (enero-marzo) de 1989. Informativo Oceanográfico Vol. 4 Nº 1. IMARPE. p. 15
Correspondencia
Dirección de Investigaciones Oceanográficas-proyecto CENSOR, Instituto del Mar del Perú, P.O. Box 22, Callao, Perú.
Email Michelle Graco: mgraco@imarpe.gob.pe
Presentado: 15/03/2006
Aceptado: 18/12/2006
