










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.14 no.2 Lima Dec. 2007
Jesús Izco1, Íñigo Pulgar1, Zhofre Aguirre2 y Fernando Santin2
1 Departamento de Botánica, Universidad de Santiago de Compostela. España.
2 Herbario Reinaldo Espinosa, Universidad Nacional. Loja. Ecuador
Resumen
El presente trabajo es un estudio de la flora de los páramos de pajonal seriales de distintos macizos montañosos de los Andes del Sur de Ecuador (provincias de Azuay y Loja) comprendidos entre los 2850 m y 3635 m. Fueron identificadas 43 familias de plantas vasculares, 120 géneros y 216 especies. La flora local es comparada con la de otros páramos de pajonal andinos; la riqueza florística (géneros y especies) es analizada por tramos altitudinales y se establece la flora característica de cada tramo del territorio estudiado. De forma complementaria, discutimos las influencias biogeográficas de la flora de los páramos de pajonal.
Palabras clave: Andes, Biodiversidad, Biogeografía, Páramo, Riqueza florística.
Abstract
A study of the flora of the bunchgrass paramo with antropic origin of different mountain ranges from the Andean areas in southern Ecuador (provinces of Azuay and Loja) was conducted. The study areas range between the 2850 m and 3635 m of altitude. The local flora is composed by 43 families, 120 genera and 216 species of vascular plants; this particular flora is compared with that of other Andean bunchgrass paramos, the floristic richness of genus and species is studied by altitudinal levels (100 m), and the characteristic flora of each level of the area is established. Finally the biogeographic aspects of the flora are discussed.
Keywords: Andes, Biodiversity, Biogeography, Bunchgrass Paramo, Floristic Richness.
Introducción
Pajonal es término de origen español derivado de paja, más concretamente de su aumentativo pajón, que se usa para designar las formaciones dominadas por gramíneas vivaces. En América del Sur en particular se refiere a las formaciones de gramíneas vivaceas amacolladas, propias de las altitudes medias y altas de los Andes. Da Lage et Métailié (2000) definen el término como formación gramínea de los Andes intertropicales que relevan a los páramos ecuatoriales de la Cordillera Oriental, en Perú y Bolivia.
Los páramos de pajonal forman parte de lo que genéricamente se denomina `páramo', con significado amplio de tierras altas, que en los Andes se sitúan entre los 10º N y los 8º S, a altitudes comprendidas entre los 3000 y los 4500 m (Chaverri y Cleef, 1997; Luteyn, 1999). Son numerosos los estudios previos sobre la tipología y las diferencias altitudinales de los páramos andinos, desde los trabajos iniciales de Cuatrecasas (1934) hasta los producidos en los últimos 25 años (Acosta, 1984; Luteyn, 1999; Rangel-Ch., 2000a; Salamanca et al., 2003; etc.). En este trabajo sólo se consideran los páramos de pajonal, formaciones herbáceas con dominancia clara de gramíneas amacolladas, presentes desde los 2850 m a los 3637 m de altitud en el área estudiada. Hay pocos datos monográficos sobre la flora de los páramos de pajonal ecuatorianos meridionales, que han recibido poca atención (Jørgensen et al., 1995). Los trabajos florísticos expresamente dedicados a este tipo de formaciones son escasos (Keating 1997, 1999; Cabrera et al., 2001; Quizhpe et al., 2002; Ulloa et al., 2004).
El área estudiada forma parte del reino Neotropical-Austroamericano, subreino Neotropical, Región Andina, según la clasificación inicial de Rivas-Martínez y Tovar (1983), modificada por Costa (2004).
En este trabajo se exponen los datos de las prospecciones realizadas en páramos de pajonal meridionales de Ecuador. Además de aportar nuevos datos sobre la flora de ese territorio se realiza un análisis preliminar sobre los grupos taxonómicos más ricos, de la riqueza florística por tramos altitudinales y una aproximación a las influencias corológicas de la flora identificada.
Material y métodos
Se han realizado dos campañas, en el mes de abril de 2004 (7 jornadas de campo) y durante el mes de junio de 2006 (5 jornadas de campo). El análisis se ha restringido a los páramos de pajonal, a formaciones con predominancia de gramíneas amacolladas, incluidos aquellos con una cobertura de especies leñosas inferior al 10%.
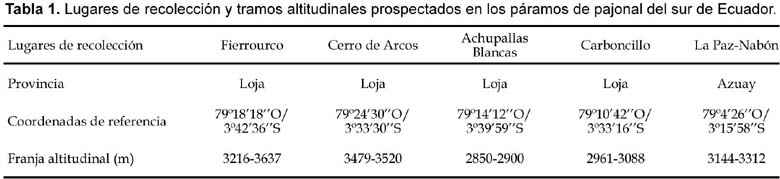
El área prospectada comprende la parte meridional de la provincia de Azuay y norte de la de Loja (Figura 1). Se han estudiado cinco territorios, que agrupamos bajo las denominaciones Fierrourco, Cerro de Arcos, Achupallas Blancas, Carboncillo y La Paz-Nabón. Los datos correspondientes a esos territorios se detallan en la Tabla 1.
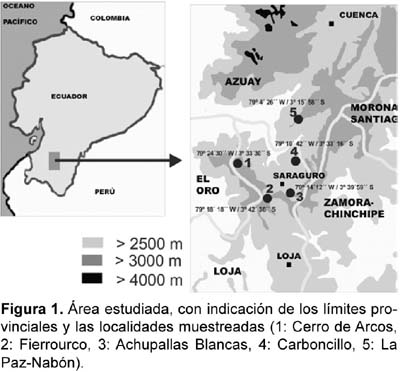
Las muestras se encuentran depositadas en el herbario de la Universidad de Santiago de Compostela (Herbario SANT) (España) y en el herbario Reinaldo Espinosa, de la Universidad Nacional de Loja (Herbario LOJA) (Ecuador). La nomenclatura empleada sigue básicamente el catálogo de plantas vasculares del Ecuador (Jørgensen y León-Yánez, 1999).
Resultados
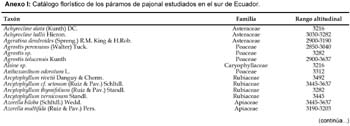
En el área estudiada, los páramos de pajonal comienzan a los 2800 m. Los más bajos son fragmentarios y se insertan entre formaciones arbustivas o arboladas, naturales o plantaciones de Pinus patula. A partir de los 3000 m los páramos de pajonal se extienden en superficies continuas, más o menos extensas, en el área de Carboncillo, otro tanto ocurre en las planicies onduladas de La Paz y La Jarata. En las otras localidades estudiadas cubren las cumbres de los macizos montañosos. La flora de los páramos de pajonal estudiados en el sur de Ecuador se compone de 43 familias, 120 géneros y 216 especies vasculares (Anexo I), con una distribución entre Anthophyta y Pteridophyta que se resume en la tabla 2. Destaca la alta proporción de helechos (10%), aunque no hay lagunas ni depresiones encharcadas en las áreas prospectadas.

Hay sustanciales variaciones de la riqueza florística por rangos y grupos sistemáticos. La curva de riqueza de géneros y de especies por familias responde a un modelo típico en L, con algunas irregularidades lógicas, de manera que unas pocas familias reúnen la máxima riqueza frente a muchas familias con representación progresivamente menor. Las diez más ricas en géneros, un cuarto del total, reúnen dos tercios de los géneros; otro tanto ocurre con las especies.
Las 10 familias más ricas en géneros, en secuencia progresiva descendente, son: Asteraceae, Poaceae, Cyperaceae, Ericaceae, Orchidaceae, Apiaceae, Caryophyllaceae, Gentianaceae, Scrophulariaceae, Lycopodiaceae (Fig. 2). Estas familias son igualmente muy ricas en especies; sin embargo, la ordenación por riqueza de especies es algo diferente. Además de la diferente ordenación por especies que se evidencia en la figura 2, es más llamativa la entrada de la Melastomataceae entre las diez familias más ricas, con sólo 2 géneros (Brachyotum y Miconia) pero con 12 especies, y las Rubiaceae (3 géneros y 8 especies).
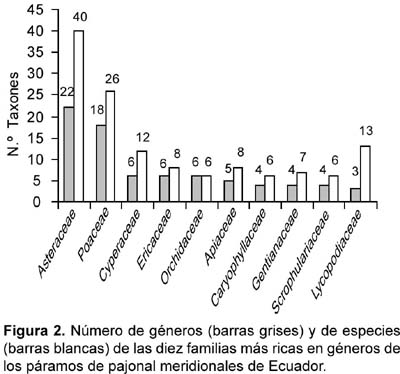
Por su riqueza en especies destacan los géneros Huperzia (7), Brachyotum (6), Hypericum (6), Miconia (7), Carex (5), Geranium (5) y Lycopodium (5). Estos 7 géneros reúnen el 18% de las especies, aunque su importancia por cobertura en el páramo de pajonal es pequeña. En la relación de géneros más ricos en especies resulta sorprendente la ausencia de géneros de las dos familias más numerosas en este rango, Asteraceae (40 especies) y Poaceae (26 especies), circunstancia probablemente relacionada con su mayor riqueza en géneros, con reparto de la diversidad específica. Llama la atención la alta riqueza específica de los géneros Huperzia y Lycopodium (Lycopodiaceae), que compiten en riqueza con las plantas con flores. A pesar de haber excluido del análisis los páramos de pajonal con cobertura leñosa superior al 10%, sorprende la riqueza de diversos géneros leñosos: Brachyotum (6 spp.), Hypericum (6), Miconia (7), Arcytophyllum (4), Baccharis (4) y Puya (4). A pesar de la frecuencia de especies leñosas no son determinantes en la morfología y estructura de los páramos de pajonal.
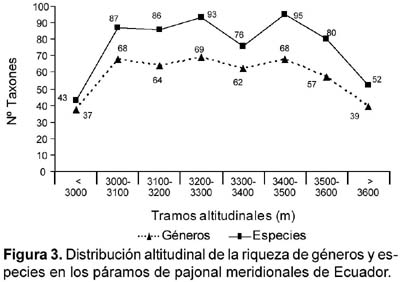
Como puede observarse en la figura 1, el análisis florístico abarca un gran tramo altitudinal, entre los 2850 y los 3637 m. Con objeto de discernir si hay patrones en la distribución altitudinal de la flora hemos realizado un análisis de riqueza por tramos de 100 m y una caracterización florística de los niveles más bajo y más alto. La curva de riqueza florística (géneros y especies), por tramos altitudinales (Fig. 3), tiene forma de meseta aplanada. Los valores más bajos de riqueza genérica se presentan en el tramo inferior (<3000 m) y superior (>3500 m), con valores de 37 y 39 géneros, respectivamente. La meseta de altitudes intermedias (3000—3500 m) dibuja una línea quebrada, con valores que van desde 62 a 69 géneros, según los distintos tramos. Las especies siguen el mismo modelo, con una curva paralela a la anterior, con valores más altos, como es lógico. En el tramo de alturas más bajas se contabilizan 43 especies y en el más alto 52; la meseta central de la curva presenta valores entre 76 y 95 especies, según los distintos tramos.
Aún no está bien determinado el rango altitudinal de las especies en los páramos de pajonal meridionales de Ecuador. Con objeto de aportar información en este sentido hemos analizado la distribución en altura de las especies del Anexo 1. Un grupo relativamente amplio (33 especies) está presente en tramos de más de 500 m (Tabla 3), entre los cuales Agrostis tolucensis, Calamagrostis bogotensis, Calamagrostis intermedia, Halenia weddeliana, Hieracium frigidum, Hypochaeris sessiliflora, Oreobolus goeppingeri, Oritrophium peruvianum, Pedicularis incurva, Rhynchospora cf. tenuis y Sisyrinchium tinctorium sobrepasan los 700 de desnivel.

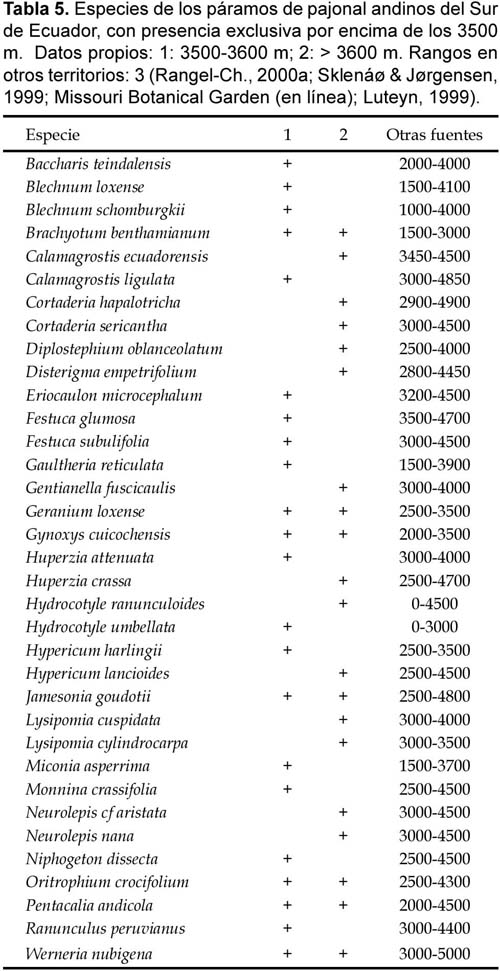
A partir de los datos propios se observan comportamientos altitudinales concretos. Una treintena de especies de los páramos de pajonal estudiados están restringidas a alturas por debajo de los 3100 m (Tabla 4), otras tantas son exclusivas de altitudes superiores a los 3500 m (Tabla 5).

En estos momentos no es posible un análisis corológico preciso de la flora de los páramos de pajonal estudiados, pero se pueden observar tendencias. Con base en la obra de Takhtajan (1986) se aprecia que las familias mejor representadas son cosmopolitas, frecuentemente con óptimo holártico, como Asteraceae, Poaceae, Cyperaceae, Orchidaceae y Lycopodiaceae, más las minoritarias Iridaceae, Violaceae, Gentianaceae, Plantaginaceae, etc. Un segundo grupo es de óptimo tropical-subtropical, aunque tenga representantes en territorios extratropicales, como Melastomataceae, Bromeliaceae, Blechnaceae, Clusiaceae, Cunoniaceae, Dennstaedtiaceae, Myrtaceae, Myrsinaceae, Eriocaulaceae, Proteaceae, Xyridaceae, etc.
Entre los géneros se repite el componente tropical-subtropical, con especial significado de las orquidáceas (Cyrtochilum, Elleanthus, Gomphichis, Habenaria, Oncidium, Stelis) y bromeliáceas (Puya). Los representantes de estas dos familias son terrícolas, dada la estructura de los páramos de pajonal.
Muy pocas especies de los páramos de pajonal alcanzan áreas extracontinentales, Lycopodium clavatum, Oxalis tuberosa (introducida en Europa), Anthoxanthum odoratum (europea introducida en América).
Discusión
En el estudio se incluyen las formaciones de La Paz, La Jarata y los páramos por encima de Nabón, a ambos lados de la carretera Panamericana. Aunque la población de Nabón se sitúa en niveles inferiores, como indican Luteyn & Gavilanes (1999), en la parte alta existen extensos páramos de pajonal.
En ningún caso se alcanza el nivel de superpáramo. El límite inferior de los páramos de pajonal identificado por nosotros coincide con el señalado inicialmente por Acosta (1984) y reiterado por autores posteriores (Sierra et al., 1999; Luteyn, 1999; Quizhpe et al., 2002). Para López (2002) el páramo de Loja comienza un poco más arriba. Por otro lado, Acosta (1984) indica un inicio ligeramente inferior de los páramos meridionales con respecto a los del norte y centro de Ecuador.
No es fácil hacer comparaciones de riqueza florística con otros territorios u otras formaciones análogas. Las dificultades vienen por escasez de datos y, sobre todo, por ausencia de criterios homogéneos en la prospección, con garantía para una correcta confrontación. A pesar de ello, es posible enfrentar datos y obtener algunos resultados.
En comparación con otros estudios sobre los páramos, los páramos de pajonal meridionales estudiados son más pobres. Según Jørgensen (1992), citado por Sklenáø & Jørgensen (1999) el conjunto de la flora paramuna ecuatoriana por encima de los 4000 m comprende 200 géneros y 508 especies. Nuestros datos están también por debajo de la flora paramuna colombiana (3100—4000 m), páramos de Chingaza (76 familias, 274 géneros y 534 especies), asímismo de los páramos de Sumapaz (77 familias, 251 géneros y 614 especies) (Rangel-Ch., 2000b). La riqueza florística es también superior en cualquiera de los tramos de vegetación paramuna colombiana, salvo las especies del piso Altoandino de Sumapaz. Sin duda, ello tiene que ver con el número de hábitats de los estudios colombianos, que incluyen páramos de pajonal y otras formaciones. Los datos propios muestran una riqueza florística análoga a la de otros espacios próximos como los páramos de Cajamuna (Parque Nacional Podocarpus) (50 familias, 81 géneros, 130 especies) (Keating, 1997), flora paramuna del Parque Nacional Podocarpus (61 familias, 93 géneros y 221 especies (Cabrera et al., 2001).
La riqueza de géneros y especies por familia refleja una cierta coincidencia en las familias más ricas de distintos páramos y pajonales andinos (Tabla 6). Las diez familias más ricas de esos territorios coinciden en buena parte. En un total de seis territorios comparados el conjunto de familias más ricas es de poco más de 20. Asteraceae y Poaceae ocupan por lo general los primeros lugares, en concordancia con la dominancia de estas familias en las altas montañas tropicales (Smith & Cleef, 1988). Sin embargo, una buena radiografía de la situación debe trabajar con cifras relativas de las familias, para evitar la sobre-representación de las familias más ricas en términos absolutos, aunque no es posible por falta de datos de referencia. Por otro lado, las discrepancias entre los distintos territorios seguramente tienen más que ver con las diferencias ecológicas de las comunidades prospectadas (rangos altitudinales, tipos de vegetación, etc.) que con un significado corológico. A pesar de todo resulta evidente la mayor semejanza entre las localidades ecuatorianas y colombianas que entre estas y las peruanas estudiadas por Yarupaitán & Albán (2003) y Flores et al. (2005). A ello contribuye también que la flora de las lagunas de Pomacocha y Habascocha (Junín, Perú) corresponde a un medio higroturboso de altura (4350—4450 m).
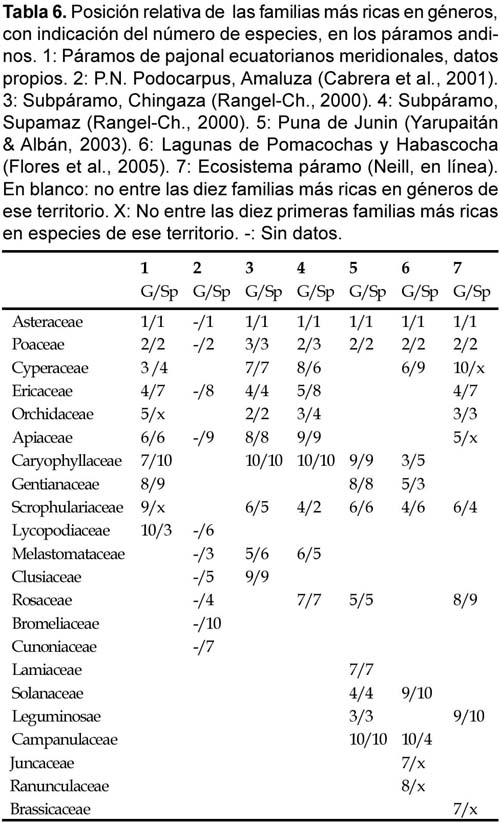
La comparación de las floras de los distintos territorios de la tabla 6 muestra diferencias notables, probablemente como resultado de sus distintas condiciones ambientales (posición en la cordillera oriental u occidental, inclusión de formaciones leñosas o herbáceas, tramo altitudinal, etc.) o, por lo menos, influenciado por ellas. Así, la flora de las lagunas de Pomacochas y Habascocha es rica en juncáceas y ranunculáceas, ligadas a medios húmedos. En los páramos de pajonales meridionales de Ecuador, las Asteraceae y Poaceae son las familias más ricas en géneros, como es habitual en los páramos, le siguen un grupo de familias (Ericaceae, Orchidaceae, Cyperaceae, Gentianaceae, Scrophulariaceae, Apiaceae) que ocupan también lugares relevantes en otras localidades paramunas. Los páramos de pajonal estudiados se desvían algo de las listas de familias paramunas más ricas por la riqueza de licopodiáceas.
Resulta evidente que la distribución altitudinal de la flora de los páramos de pajonal estudiados no refleja con precisión los rangos absolutos de las especies. La amplitud vertical de la flora altoandina ecuatoriana es muy grande, sobre un total de 138 especies que habitan por encima de los 4000 m, más de la mitad tiene una amplitud de más de 1300 m y un tercio más supera los 1000 m de desnivel (Sklenáø & Jørgensen, 1999). Los datos colombianos de Chingaza y Sumapaz (Rangel-Ch., 2000) y de los páramo altos ecuatorianos (Sklenáø y Jørgensen, 1999) incluidos en las tablas 4 y 5 ponen en evidencia que algunas de las plantas restringidas a niveles inferiores (<3100 m) en los páramos de pajonal estudiados alcanzan cotas superiores a los 4000 m en otras localidades, entre ellas Baccharis tricuneata y Gaultheria erecta. Otras parecen más ajustadas a niveles relativamente bajos: Myrcianthes myrsinoides, Stellaria recurvata, Viola arguta. También hay distorsiones en las plantas de tramos altos; una buena parte de las que están presentes en niveles por encima de los 3500 m en los páramos de pajonal sudecuatorianos habitan también tramos altos en los páramos de Colombia: Eriocaulon microcephalum, Festuca subulifolia, Jamesonia goudotii, Neurolepis nana; otras descienden muy por debajo de ese nivel: Blechnum schomburgkii, Cortaderia hapalotricha, Disterigma empetrifolium, Hypericum lancioides.
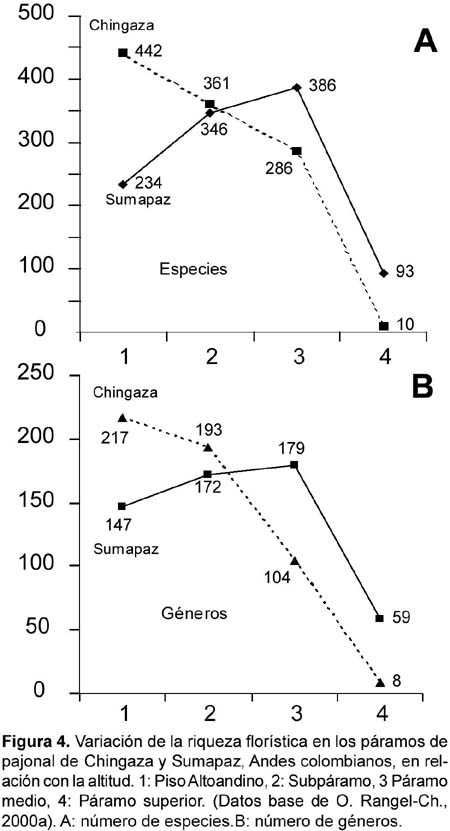
La riqueza específica de los Andes ecuatorianos presenta su máximo a los 2700—2800 m y decrece a partir de este límite (Jørgensen et al., 1995), aunque puede haber razones ajenas a la altitud. En la distribución altitudinal de la riqueza de géneros y de especies en los páramos de pajonal ecuatorianos meridionales, no tienen fácil explicación las bajas cifras de géneros y especies por debajo de los 3000 m. El tramo es más amplio que otros, abarca 150 m de altitud, y se inserta en niveles globalmente más ricos. Es posible que el fenómeno se deba a una subrepresentación de muestreo, es decir, el número de especies inventariadas está fuertemente correlacionado con el número de muestras recolectadas (Sklenáø & Jørgensen, 1999). La caída en el tramo más alto (>3600 m) tiene explicación en su menor amplitud y un empobrecimiento de hábitats. El modelo de los páramos de pajonal de Azuay y Loja es semejante al de los páramos de Sumapaz (Colombia), pero discrepa del modelo de los páramos de Chingaza, con caídas de flora continuas al ascender en altitud (Rangel-Ch., 2000) (Figura 4). Keating (1999) ha estudiado la riqueza florística en relación con la variación altitudinal y aporta como resultado un marcado incremento de especies relacionado con el descenso en altura, sobre todo de las herbáceas, con inflexión alrededor de los 3500 m.
La flora andina a partir de los 3000 m circula facilmente a lo largo de la cordillera. El 72 % de las especies ecuatorianas por encima de esas altitudes es transandina (Balslev, 1988), más una importante proporción de endemismos locales. De los siete elementos florísticos que Cleef (1979) considera en los Andes Colombianos Orientales, tres son claramente reconocibles en los páramos de pajonal meridionales de Ecuador: holártico, tropical (Reino Neotropical-Austroamericano), austral (Reino Antárctico) (Figura 5). Una gran parte de géneros tiene su origen o su óptimo en el mundo Holártico, como es habitual en las altas montañas tropicales (Luteyn, 1999). Los representantes del elemento austral tienen apenas una presencia testimonial.
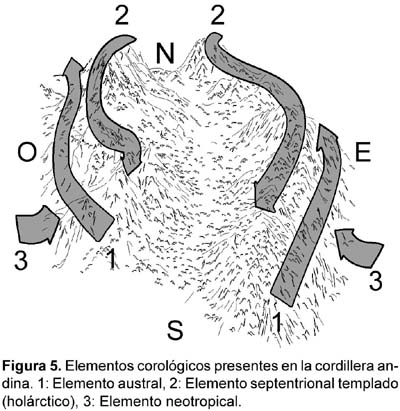
Los representantes neotropicales de los paramos de pajonal estudiados provienen de los pisos inferiores andinos y son muy numerosos: Neurolepis, Ageratina, Puya, Loricaria, Disterigma, Chaptalia, Jamesonia, Halenia, Diplostephium, Gonphichis, Gynoxys, Miconia, Lysipomia, etc. Muchos de los géneros de este elemento tienen consistencia leñosa (Rubiaceae, Melastomataceae, Myrtaceae, Bromeliaceae), frente a la naturaleza herbácea dominante de los páramos de pajonal.
La flora de significado septentrional está ligada al cierre del vano centroamericano y la posterior elevación de los Andes y las montañas centroamericanas, lo que permitió el desplazamiento hacia el Sur de flora septentrional templada en sucesivos periodos de expansión y contracción (Raven y Axelrod, 1981; Smith y Cleff, 1988; van der Hammen, 1988; Luteyn, 1999; Takhtajan, 1986; Flores, 2000; Sklenáø & Ramsay, 2001), en distintas oleadas a lo largo de los últimos 2—3 millones de años, principalmente durante los periodos fríos del Pleistoceno. Además, esa proporción se incrementa con la altitud (Luteyn, 1999). Durante los últimos impulsos glaciales todos los pisos estaban desplazados a niveles más bajos, ampliando el pasillo para el desplazamiento de flora antártica hacia el norte y de flora septentrional hacia el Sur; ese desplazamiento supuso también un acercamiento entre macizos montañosos, facilitando la migración. El análisis sobre el origen de las floras tropicales de altura (trópico-alpinas) (Smith & Cleef, 1988) revela un equilibrio entre géneros australes y septentrionales en los Andes de Colombia y Venezuela, aproximadamente 50% en cada caso. Para Monasterio (1980), según recoge Luteyn (1999), la jalca y los páramos ecuatorianos formados particularmente por macollas de gramíneas están formados principalmente por géneros extratropicales. El movimiento hacia el Sur se manifiesta por la presencia de familias como Berberidaceae, Caryophyllaceae, Myricaceae, Rosaceae, Symplocaceae, Gentianaceae, Plantaginaceae, Scrophulariaceae, etc. (Raven y Axelrod, 1981) y géneros como Bartsia, Castilleja, Gentiana, Hypericum, Hypochaeris, Symplocos, Valeriana, Pedicularis, Plantago, Stipa, Berberis, Huperzia, Lycopodium, Vaccinium etc. (Cleff, 1979; Takhtajan, 1986; Smith & Cleef, 1988).
El movimiento de flora austral hacia el norte, hasta alcanzar Ecuador, está representado por los géneros Azorella, Cortaderia, Oreobolus, Nertera, Gaultheria, Oreomyrrhis y Pernettya. Azorella es un género principalmente andino-patagónico, representado en el área por dos especies de la sección Ciliatae (A. biloba y A. multifida), las dos de distribución andina, que en el caso de la primera se extiende hasta Costa Rica (Martínez, 1993).
En los páramos de pajonal estudiados se pone en evidencia la doble adscripción corológica clásica de algunos grupos, por ejemplo las rubiáceas leñosas (Arcytophyllum, Nertera) son tropicales, mientras que las herbáceas pertenecen al género Galium, de óptimo holártico. Algo análogo ocurre con las Ericaceae, familia representada por géneros neotropicales como Bejaria y Disterigma, géneros australes como Pernettya y Themistoclesia, más la presencia de Vaccinium, que tiene su óptimo en el reino Holártico.
Agradecimientos
El trabajo se ha realizado en el marco del proyecto AECI _CPI, Acciones Conjuntas D/1033/03, al que se sumaron mediante convenio la Universidad de Santiago de Compostela (España) y la Universidad Nacional de Loja (Ecuador). Agradecemos particulamente el rector de la UNL, Max González Elizalde, su soporte técnico y de infraestructura para realizar el trabajo. A Modesto Luceño y Simon Laegaard por su asistencia en la determinación de algunos grupos conflictivos. A Pablo Rodrigo Cuenca y Leonardo Ordoñez, de la Fundación Ecológica Arcoiris, por su asistencia durante los trabajos de campo; a la Fundación Kawsay (Saraguro, Loja) por su acogida durante nuestra estancia en Ecuador en 2004, y en concreto a Gustavo Beltrán por su labor técnica durante el trabajo de campo. A Montserrat Villarino, miembro del equipo del proyecto, por su asistencia durante las campañas en Ecuador.
Literatura citada
Acosta M. 1984. Los páramos andinos del Ecuador. Quito: Publicaciones Científicas MAS.
Balslev H. 1988. Distribution patterns of ecuadorean plant species. Taxon 37(3): 567-577.
Cabrera O., Z. Aguirre & B. Maza. 2001. Planificación para la conservación de sitios. Loja: Páramos del Parque Nacional Podocarpus, Fundación ecológica Arcoiris & The Nature Conservancy.
Chaverri A. & A.M. Cleef. 1997. Las comunidades vegetacionales en los páramos de los macizos de Chiripó y Buenavista, Cordillera de Talamanca, Costa Rica. Rev. Forest. Centroamericana, 17: 44-49.
Cleef A.M. 1979. The phytogeographycal position of the Neotropical Vascular Páramo Flora with special reference to the Colombian Cordillera Oriental. In: K. Larsen and L.B. Holm-Nielsen, eds. Botany. Academic Press. London.Pp: 175-184.
Costa M. 2004. Biogeografía. In: J. Izco, ed. Botánica. McGraw Hill Interamericana. Madrid.Pp: 794-852.
Cuatrecasas J. 1934. Observaciones geobotánicas en Colombia. Trab. Museo Nac. Ciencias Naturales, Ser. Botánica 27 : 1-144.
Da Lage A. & G. Métailié. 2000. Dictionnaire de biogéographie végétale. Paris: CNRS Editions.
Flores A. 2000. Geomorfología de los páramos. In: J.O. Rangel-Ch, ed. Colombia. Diversidad biótica, III. La región de vida paramuna. Univ. Nacional de Colombia. Bogotá. Pp: 24-36.
Flores M., J. Alegría & A. Granda. 2005. Diversidad florística asociada a las lagunas andinas Pomacocha y Habascocha, Junin, Perú. Rev. Peruana Biol. 12 (1): 125-134.
Jørgensen, J.P. 1992. Vegetation of the high Andes of Ecuador. PhD. Thesis. Institute of Biological Sciences. Aarhus University.
Jørgensen P.M. & S. León-Yánez. 1999. Catálogo de las plantas vasculares del Ecuador. Saint Louis: Missouri Botanical Garden.
Jørgensen P.M., C. Ulloa, J.E. Madsen & R. Renato. 1995. A floristic analysis of the High Andes of Ecuador. In: S.P. Churchill et al., eds. Biodiversity and Conservation of Neotropial and Montane forests. New York Bot. Gard. New York. Pp: 221-237.
Keating P.L. 1997. An inventory of plant species in the páramo of Cajanuma, Podocarpus National Park (Ecuador). Phytologia 83: 333-344.
Keating P.L. 1999. Changes in páramo vegetation along an elevation gradient in southern Ecuador. Journal Torrey Bot. Soc. 126(2): 159-175.
López F. 2002. Ecuador-Perú: conservación para la paz. Loja: Fundación ecológica Arcoiris & Univ. Téc. Part. de Loja.
Luteyn J.L. 1999. Introduction to the paramo ecosystem. In: J.L. Luteyn, ed. Páramos, a checklist of Plant Diversity, Geographical Distribution, and Botanical Literature. New York Bot. Gard. New York. Pp: 1-39.
Luteyn J.L. & M.R. Gavilanes. 1999. Ecuador. In: J.L. Luteyn, ed. Páramos, a checklist of Plant Diversity, Geographical Distribution, and Botanical Literature. New York Bot. Gard. New York. Pp: 193-218.
Martínez S. 1993. Sinopsis del género Azorella (Apiaceae, Hydrocotyloideae). Darwiniana 32(1-4): 171-184.
Missouri Botanical Garden. 2002. Checklist of the Vascular Plants of Ecuador. W3TROPICOS. (en línea) <http://mobot.mobot.org/W3T/Search/index/Ecuadort.html> Acceso 09/03/2007.
Monasterio M. 1980. Los páramos andinos como región natural. In: M. Monasterio, ed. Estudios ecológicos de los páramos andinos. Univ. de los Andes. Mérida. Pp: 15-27.
Quizhpe W., Z. Aguirre, O. Cabrera & T.E. Delgado. 2002. Los páramos del Parque Nacional Podocarpus. In: Z. Aguirre, J.E. Madsen, E. Cotton and H. Balslev, eds. Botanica austroecuatoriana. Abya Yala. Quito.Pp: 79-89.
Rangel-Ch. J.O. 2000a. La región paramuna y la franja aledaña de Colombia. In: J.O. Rangel-Ch, ed. Colombia. Diversidad biótica, III. La región de vida paramuna. Univ. Nacional de Colombia. Bogotá. Pp: 1-23.
Rangel-Ch. J.O. 2000b. Catálogo florístico de los macizos de Chingaza y Sumapaz. In: J.O. Rangel-Ch, ed. Colombia. Diversidad biótica, III. La región de vida paramuna. Univ. Nacional de Colombia. Bogotá. Pp: 563-598.
Raven P.H. & D.I. Axelrod. 1981. Angiosperm biogeography and Past Continental Movements. Ann. Missouri Bot. Gard. 61: 539-673.
Rivas-Martínez S. & O. Tovar. 1983. Síntesis biogeográfica de los Andes. Collectanea Botanica 14: 515-521.
Sklenáø P. & P.M. Jørgensen. 1999. Distribution patterns of páramo plants in Ecuador. Journal of Biogeography 26: 681-691.
Sklenáø P. & P.M. Ramsay. 2001. Diversity of zonal páramo plant communities in Ecuador. Diversity and Distribution 7: 113-124.
Salamanca S., A.M. Cleef & O.J. Rangel-Ch. 2003. The paramo vegetation of the volcanic Ruiz-Tolima massif. In: T. van der Hammen and A.G. Dos Santos, eds. La Cordillera Central Colombiana. Transecto Parque Los Nevados. J. Cramer. Berlin. Pp: 1-77.
Sierra, R. 1999. Propuesta preliminar de un sistema de clasificación de vegetación para el Ecuador continental. Proyecto INEFAN/GEF-BIRF y Eco-Ciencia. Quito.
Smith J.M.B. & A.M. Cleef. 1988. Composition and origins of the world's tropicalpine floras. Journal of Biogeography 15: 631-645.
Takhtajan A. 1986. Floristics regions of the world. Berkeley: Univ. California Press.
Ulloa C., S. Álvarez, P.M. Jørgensen & D. Minga. 2004. Cien plantas silvestres del páramo. Cuenca: P.N. Cajas, ETAPA & Missouri Bot. Garden.
Van der Hammen T. 1988. The tropical flora in historical perspective. Taxon 37 (3): 515-518.
Yarupaitán G. & J. Albán. 2003. Flora silvestre de los Andes centrales del Perú: un estudio en la zona de Quilcas, Junín. Rev. Peruana Biol. 10 (2): 155-162.
Correspondencia
1 Departamento de Botánica, Universidad de Santiago de Compostela. E- 15782 Santiago de Compostela. España.
Email: Jesús Izco bvizco@usc.es
Presentado: 28/03/2007
Aceptado: 06/12/2007
