










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.15 no.2 Lima 2008
TRABAJOS ORIGINALES
Caza y estado de conservación de primates en la cuenca del río Itaya, Loreto, Perú
Hunting and conservation status of primates populations in the Itaya river basin, Loreto, Peru
Rolando Aquino1 ; Wagner Terrones2 ; Raúl Navarro2 ; César Terrones2 ; Fanny M. Cornejo3
1. Instituto de Ciencias Biológicas Antonio Raimondi (ICBAR), Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. P.O. Box 575, IVITA-Iquitos, Perú. Email: Rolando Aquino: raquino2005@yahoo.es
2. Centro Amazónico de Educación Ambiental e Investigación (ACEER), Morona 517, Iquitos.
3. Museo de Historia Natural, Universidad Nacional Mayor de San Marcos. Apartado Postal 14-0434, Lima 14, Perú.
RESUMEN
Este reporte contiene información relacionada a la diversidad e impacto de la caza de los primates, así como las amenazas potenciales para sus poblaciones que habitan en la cuenca del río Alto Itaya. Está basado en censos por transectos y registros de caza llevados a cabo en seis comunidades. Como resultado de los censos, fueron registrados 384 grupos pertenecientes a 9 de las 11 especies que habitan en el área de estudio, correspondiendo los mayores registros al pichico pardo (Saguinus fuscicollis Spix ) con 25,3%, huapo negro (Pithecia aequatorialis Hershkovitz) con 20,6% y mono choro (Lagothrix poeppigii Humboldt) con 15,9%. La densidad poblacional fue estimada para nueve especies, siendo Saimiri sciureus Linnaeus la más abundante con 18,2 individuos/km2, seguido por L. poeppigii con 15,5 individuos/km2 y S. fuscicollis con 10,5 individuos/ km2; lo contrario ocurrió en Alouatta seniculus Linnaeus, cuya densidad fue estimada en 0,15 individuos/ km2. Del área de caza estimada en 600 km2 fueron extraídos un promedio anual de 262 ejemplares y cuya biomasa fue calculada en 1677,9. kg. Del total, 119 individuos equivalente al 45% correspondió a L. poeppigii, cuya presión de caza fue calculada en 0,19 individuos/km2. La aplicación del modelo de cosecha sugiere sobrecaza en poblaciones de A. seniculus, L. poeppigii y C. apella. La caza y la deforestación con fines agrícolas son las amenazas potenciales para las poblaciones de primates.
Palabras claves: Cuenca del río Itaya, primates, presión de caza, impacto de la caza, amenazas potenciales.
ABSTRACT
This report contains information related to the diversity and hunting impact of the primates, as well as the potential threats for the populations that inhabit the upper Itaya river basin. It is based on census by transect and hunting records carried out in six communities. During the census 384 groups belonging to 9 were observed of the 11 species that inhabit the study area. Most records corresponded to Saddle-back tamarin (Saguinus fuscicollis Spix) with 25,3%, saki monkey (Pithecia aequatorialis Hershkovitz) with 20,6% and woolly monky (Lagothrix poeppigii Humboldt) with 15,9%. Population density was estimated for nine species, being the squirrel monkey (Saimiri sciureus Linnaeus) the most abundant with 18,2 individuals/km2, followed by woolly monkey (L. Poeppigii) with 15,5 individuals/km2 and saddle-back tamarin (S. fuscicollis) with 10,5 individuos/km2; the opposite occured in red howler monkey (Alouatta seniculus Linnaeus) whose density was estimated in 0,15 individuals/km2. Of the hunting area estimated in 600 km2 were harvested an annual average of 262 individuals, whose biomass was estimated in 1677,9 kg. Of them, 119 individuals equivalent to 45% corresponded to woolly monkey (L. poeppigii) whose hunting pressure was calculated in 0,19 individuos/km2. The application of the harvest model suggests overhunting in populations of red howler monkey (A. Seniculus), woolly monkey (L. Poeppigii) and brown capuchin monkey (Cebus apella). Hunting and deforestation with agricultural purpose are the potential threats for the primates populations.
Keywords: Itaya river basin, primates, hunting pressure, hunting impact, potential threats.
Introducción
Entre los componentes de la fauna silvestre de la Amazonía peruana, los primates constituyen fuente de proteína animal para los pobladores ribereños, por lo que son cazados para el consumo de subsistencia en sustitución a los ungulados y roedores grandes, que por su mayor biomasa son destinados a la venta en los principales mercados del oriente peruano (Aquino et al. 2000), aunque algunas veces también tienen como destino la venta, en particular los denominados de gran tamaño como el choro (Lagothrix spp.) (Bodmer y Pezo 2000). En lo que concierne al nororiente peruano, los estudios realizados por Bodmer el at. (1997; 1999; 2000), Aquino et al. (2000; 2001; 2003) y de otros, dan cuenta de la sobrecaza en poblaciones de algunas especies, en particular de aquellas consideradas de tamaño grande como los maquizapas (Ateles spp.), choros (Lagothrix spp.) y mono aullador (Alouatta seniculus Linnaeus). La cuenca del río Alto Itaya no escapa a esta realidad; en ella se encuentran habitando diversas especies de la fauna silvestre, entre ellos los primates que están sujetos a la caza. De acuerdo con los mapas de distribución de Hershkovitz (1977, 1988), Aquino y Encarnación (1994), Rylands et al. (1995), Van Roosmalen et al. (2002) y la referencia de Alvarez y Soini (2000), en estos bosques estarían habitando alrededor de 14 especies de primates, entre ellas el supay pichico (Callimico goeldii Thomas) y el tocón negro (Callicebus torquatus Thomas). Por otro lado, el único estudio sobre impacto de caza para la mencionada cuenca fue realizado por Aquino et al. (2007), mientras tanto las actividades de caza, extracción de hojas de irapay (Lepidocaryum sp.), extracción de madera de valor comercial (bajo la modalidad de concesiones forestales) y colecta de frutos silvestres es cada vez mayor, los que estarían generando serias alteraciones a los hábitats en desmedro de las poblaciones de primates y de otros componentes de la fauna silvestre, con riesgo incluso de extinción local de algunas especies si es que no se adoptan medidas correctivas que garanticen la conservación de la biodiversidad.
La carencia de información, en particular en lo que se refiere a inventarios, evaluación y presión de caza, nos motivó a la conducción del presente estudio, el mismo que se llevó a cabo de marzo del 2004 a diciembre del 2005 en la cuenca del río Alto Itaya. Los objetivos del estudio estuvieron orientados a: determinar las especies de primates que habitan en el área, evaluar el impacto de la caza en poblaciones de primates considerados de tamaño grande y mediano y analizar las amenazas potenciales a las que están expuestos sus poblaciones.
Área de estudio
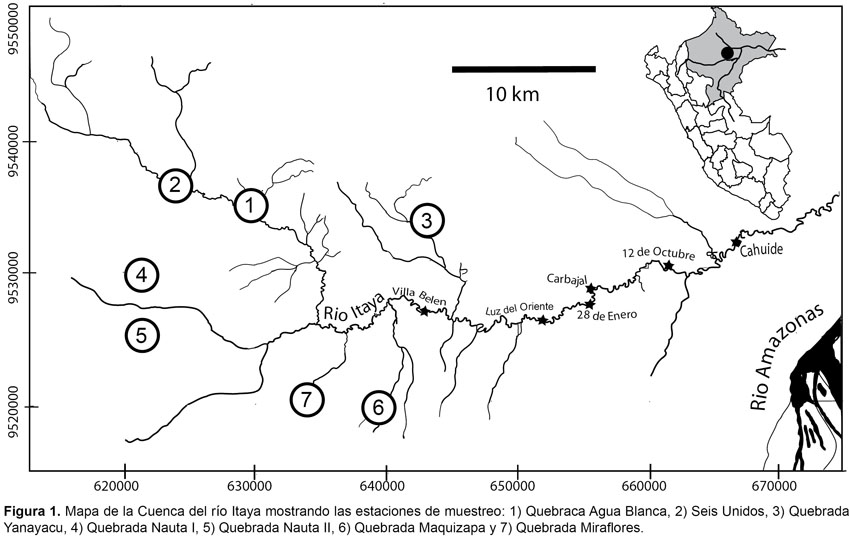
El estudio comprendió principalmente la cuenca del río Alto Itaya (Fig. 1), desde la altura de Luz de Oriente, penúltima comunidad asentada en el río Alto Itaya, aguas arriba hasta muy cerca de las cabeceras del citado río. Este río tiene como principales afluentes a las quebradas Nauta, Maquizapa, Yanayacu, Miraflores y Agua Blanca. En el área, la fisiografía del terreno se caracterizó por la presencia de terrazas altas y colinas bajas con pendientes de hasta 60% de inclinación. En ellas, la cobertura vegetal se desarrolla en suelo areno-arcilloso, cuya composición florística compuesto por árboles de fuste casi rectos alcanzaron alturas entre 10 a 25 m, con algunos emergentes superiores a 35 m como pashaco (Parkia sp.), quinilla (Elaeoluma sp.) sacha caimito (Manilkara sp.) y machimango (Eschweilera sp.), entre otras. Entre las palmeras, sobresalieron por su relativa abundancia la pona (Socratea sp.), cashapona (Iriartea sp.), ungurahui (Oenocarpus sp.) y chambira (Astrocaryum sp.). El sotobosque estuvo poblado algunas veces por palmeras de irapay (Lepidocaryum sp.) y en otras por plantas herbáceas perennes. Piso del bosque generalmente cubierto por un colchón de hojarascas, particularmente en terrazas alta y cimas de colina baja. El bosque desde la orilla del río y quebradas hasta 5,0 km monte adentro presentó serias alteraciones como consecuencia de la extracción de especies maderables y de hojas de irapay y por la presencia de numerosos senderos de cazadores, algunos recientemente abiertos, donde la fauna nos pareció escasa, debido posiblemente a la alta presión de caza; mientras que más allá de los 5,0 km el bosque mostró un perfil menos alterado, pero igualmente fueron encontrados senderos de cazadores en uso, aunque la presencia de animales silvestres nos pareció relativamente más abundante.
Material y métodos
Censos por transecto: fueron conducidos en ambas márgenes del río Itaya y principales afluentes, desde la orilla hasta 7 km hacia el interior del bosque y en algunos casos hasta más de 10 km. Para los censos fueron definidos siete estaciones de muestreo (Fig. 1), donde fueron abiertos de cuatro a cinco transectos, cada uno de 4 km de longitud. Los censos fueron de ida y vuelta, desde las 07:00 a 11:00 h y desde las 14:00 a 17:00 h (hora local). Cada grupo integrado por dos observadores realizaron recorridos a una velocidad promedio de 1 km/hora. Cada vez que hubo contacto con los primates se procedió a anotar en la libreta de campo la especie, número de individuos, distancia perpendicular del primer individuo observado al transecto, tamaño del grupo, altura con respecto a los estratos del bosque y actividad al momento de la observación. Un total de 1021 km fueron censados. Con excepción del mono aullador (A. seniculus), la densidad para el resto de especies fue analizada con el software DISTANCE versión 4.1. La densidad para el mono aullador fue estimada mediante la fórmula D=N/2dL (Burnham et al., 1980), donde D es la densidad de individuos/km2, N es el número de animales observados, L es la longitud censada en kilómetros, y d es la distancia perpendicular del observador al primer individuo avistado.
Registros de caza: Fueron llevadas a cabo en seis comunidades asentadas a orillas del río Alto Itaya, para el cual contamos con la activa participación de los pobladores dedicados a la caza con fines de subsistencia. Estos registros permitieron estimar el área de caza y la presión de caza (individuos extraídos/km2). Al promedio de caza anual fue adicionado un 35% para compensar los animales extraídos por cazadores que desistieron en participar en el estudio y cuya caza no fue registrada.
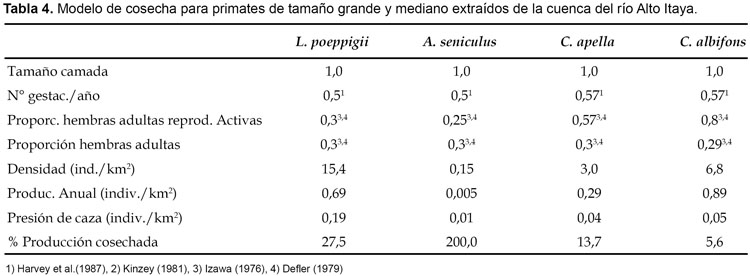
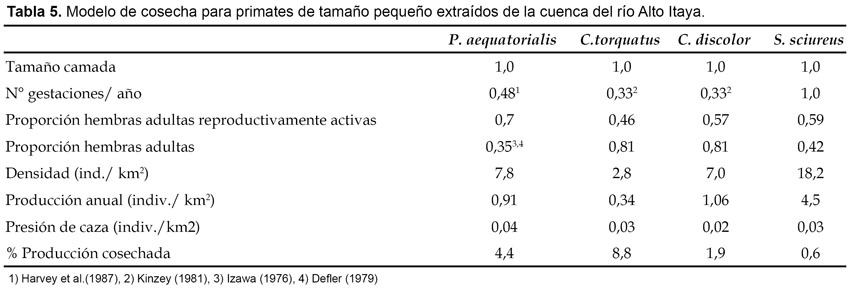
Impacto de la caza: Para este propósito se usó el modelo de cosecha y fue aplicado para especies que usualmente son cazadas para el consumo de subsistencia, por lo tanto, fue excluido del análisis el pichico común (S. fuscicollis Spix). Con excepción del tocón negro (C. torquatus), tocón colorado (Callicebus discolor I. Geoffroy & Deville) y fraile (Saimiri sciureus Linnaeus), el modelo de cosecha para el resto de primates usó información sobre productividad reproductiva de estudios previos realizados fuera del Perú. Para el huapo negro (Pithecia aequatorialis Hershkovitz), la proporción de hembras adultas reproductivamente activas fue determinada de 40 grupos, para el tocón negro de 28 grupos y para el tocón colorado de 35 grupos, todos observados de manera completa durante el estudio. La proporción de hembras adultas para ambas especies de Callicebus fue tomada del análisis de estructura poblacional de siete grupos completos de tocón colorado (Callicebus cupreus Spix) capturados en noviembre de 1989 en la cuenca del río Manití por el Proyecto Peruano de Primatología. Finalmente, la proporción de hembras adultas reproductivamente activas y hembras adultas en general para el fraile (S. sciureus) fue tomada de Ique (1990). Para determinar la sostenibilidad de caza fue asumido la recomendación de Bodmer et al. (1999), quienes consideran que los primates (salvo excepciones) son especies de vida larga, de reproducción lenta y por lo tanto asumen que solamente un máximo del 10% de su producción puede ser cosechado sosteniblemente.
Resultados
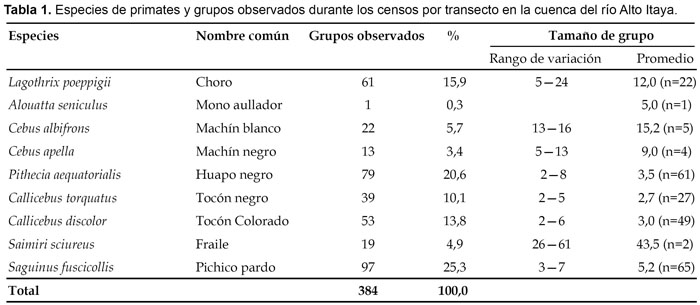
Como resultado de los censos, registro de caza y observaciones adicionales, en la cuenca del río Alto Itaya fueron registradas 11 especies de primates haciendo uso de los diversos tipos de hábitats. De ellas, nueve fueron observadas durante los censos (Tabla 1) y dos fuera de ellos. Las especies excluidas de los censos fueron el mono nocturno o musmuqui (Aotus vociferans Spix) y el leoncito (Cebuella pygmaea Spix); la primera por su hábito nocturno y la segunda por su hábitat casi exclusivo de los bosques ribereños; además, ambas no son cazadas para el consumo de subsistencia, el musmuqui por su escasa o casi nula aceptación como carne de monte debido al fuerte olor desagradable que emanan de la glándula subcaudal y el leoncito por tratarse de un animal diminuto cuyo peso no supera los 150 g.
El nulo contacto con el supay pichico (C. goeldii) en más de 1000 km de longitud censada y el desconocimiento de este primate por parte de los pobladores son argumentos suficientes para sostener que esta especie está ausente en la cuenca del río Alto Itaya. No obstante, a inicios de agosto del 2005 fue comprobada la presencia de esta especie entre el río Tigre y las cabeceras del río Alto Nanay muy cerca de la localidad de Intuto, donde dos de los autores (W. Terrones y F. Cornejo) observaron un grupo de cuatro individuos en asociación con el pichico común (S. fuscicollis), así como también un grupo de tocón negro (C .torquatus). El bosque donde se observó el grupo de supay pichico fue bastante similar a los existentes en el Alto Itaya, con la diferencia que la vegetación de tipo varillal seco, estuvo conformado por árboles de porte mucho más bajo; es decir, entre 15 a 20 m de alto en su mayoría, mientras que el piso estaba cubierto por un denso colchón de hojarascas que incluso dificultó las caminatas. Los registros obtenidos confirman que el tocón negro (C. torquatus) se encuentra habitando la cuenca del río Itaya, donde tiene como hábitat exclusivo la vegetación tipo varillal. En efecto, este tipo de vegetación está presente en ambas márgenes del río Alto Itaya, aún cuando en realidad se trataría de un varillal algo distinto a los existentes en la margen derecha del río Nanay. Este tipo de vegetación conformado por árboles de fustes delgados y casi recto se desarrollan más en suelo areno-arcilloso de color ocre y/o rojizo; cuyo piso en algunos casos estuvo cubierto por abundante hojarasca, particularmente en la cima de colina baja; mientras que el sotobosque se encontraba pobremente poblado por algunas plantas herbáceas perennes y por algunas especies de palmeras, entre ellas el irapay (Lepidocaryum sp.).
De los 384 grupos observados, los mayores registros correspondieron al pichico común (S. fuscicollis) con 25,3%, seguido por el huapo negro (P. aequatorialis) con 21,5% y el choro común (Lagothrix poeppigii Humboldt) con 15,9% (Tabla 1); lo contrario ocurrió con el mono aullador (A. seniculus), con apenas un grupo observado, lo que indicaría que en este cuenca está casi extinta y cuya escasa población se encuentra refugiada en las cabeceras de las quebradas que dan origen al río Itaya.
Sin considerar al fraile o mono ardilla (S. sciureus) y al pichico común (S. fuscicollis), el tamaño de grupo estuvo en función a la distancia con respecto al curso del río Itaya y principales afluentes. Así, en bosques cercanos a los ríos y quebradas, los grupos eran más pequeños, habiendo observado hasta de cinco individuos en el mono choro (L. poeppigii) y machín negro (Cebus apella Linnaeus), respectivamente; mientras que más allá de los 7 km de distancia y muy cerca de la naciente del río, los grupos eran mucho más grandes y menos ariscos, habiendo encontrado grupos de hasta 24 individuos en el mono choro (L. poeppigii), 16 individuos en machín blanco (Cebus albifrons Humboldt) y 13 individuos en machín negro (C. apella) (Tabla 1), por lo que se deduce que la mayor presión de caza está ocurriendo en bosques cercanos a los principales cuerpos de agua que sirven como vías de acceso para pequeñas embarcaciones y canoas.
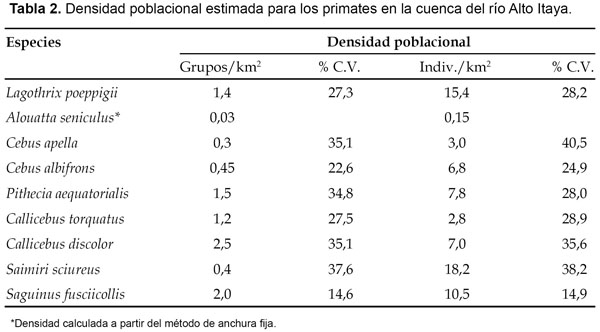
La densidad poblacional fue analizada para nueve especies. De ellas, la densidad más alta correspondió al mono ardilla (S. sciureus) con 0,4 grupos/km2 y/o 18,2 individuos/km2, seguido por el choro (L. poeppigii) con 1,4 grupos/km2 y/o 15,4 individuos/ km2 y el pichico pardo (S. fusciicollis) con 2,0 grupos/ km2 y/o 10,5 individuos/km2; mientras que la densidad más baja correspondió al mono aullador (A. seniculus) con apenas 0,03 grupos/km2 y/o 0,15 individuos/km2 (Tabla 2). Por otro lado, del área de caza estimada en 600 km2 fueron extraídos un promedio anual de 252 individuos correspondientes a ocho especies, que sumaron 1677,9 kg de biomasa extraída (Tabla 3). Sin considerar al mono aullador (A. seniculus), los registros también reflejan que la caza estuvo orientada hacia las especies de mayor biomasa. En efecto, del total de ejemplares cazados, 119 y/o 45,4% correspondió al choro (L. poeppigii), cuya presión de caza estimada en 0,198 individuos/km2 fue la más alta entre los primates (Tabla 3). En el mono aullador (A. seniculus), la presión de caza fue calculada en 0,013 individuos/km2 (Tabla 3), lo que corroboraría la escasa presencia de estos animales en el área de estudio.
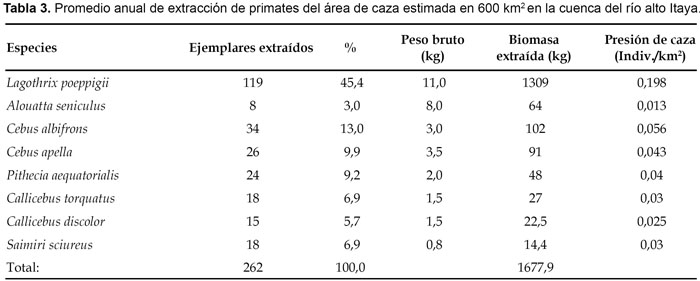
El modelo de cosecha indica que existe sobrecaza en poblaciones del mono aullador (A. seniculus), choro (L. poeppigii) y machín negro (C. apella), cuya cosecha sobrepasó el 10% de la producción sugerida para los primates (Tabla 4). De ellas, la más impactada fue el mono aullador (A. seniculus), con más del 100% de producción extraída, lo que significa que la población base está siendo reducida por haber alcanzado niveles insostenibles. Lo contrario ocurrió en poblaciones de machín blanco (C. albifrons), huapo negro (P. aequatorialis), tocón negro (C. torquatus), tocón colorado (C. discolor) y fraile (S. sciureus), cuya cosecha estuvo por debajo del 10% de la producción anual sugerida (Tabla 5), por lo tanto, está dentro de lo sostenible.
Discusión
De los resultados obtenidos, se puede afirmar que en la cuenca del río Alto Itaya, las especies de primates de tamaño grande como el choro (L. poeppigii), se encuentran generalmente habitando más allá de los 7 km de distancia con respecto a la orilla y en ambas márgenes. Naturalmente esto ocurre cuando los animales se ven presionados por las actividades de caza, por lo que tienden a refugiarse en áreas más lejanas y poco accesibles. También es evidente que la caza está orientada principalmente hacia las especies de tamaño grande y mediano, aún cuando entre los registros de caza se encuentran especies de tamaño relativamente pequeño como el tocón negro (C. torquatus), tocón colorado (C. discolor) y el fraile (S. sciureus), este último con 0,8 kg de peso como máximo. Aún así, en los bosques de la cuenca del río Itaya con excepción del mono aullador (A. seniculus), todavía existen poblaciones relativamente abundantes de primates, en particular del choro (L. poeppigii), machín negro (C. apella), machín blanco (C. albifrons), huapo negro (P. aequatorialis) y tocón colorado (C. discolor), cuyas densidades fueron superiores a los reportados por Aquino et al. (2000) para los bosques de altura de la cuenca del río Pucacuro y por Bodmer et al. (1999) para los bosques inundables de caza persistente dentro de la Reserva Nacional Pacaya Samiria. En cuanto al mono aullador (A. seniculus) y machín negro (C. apella), su escasa presencia podría estar relacionada aparte de la caza con factores ecológicos. En efecto, estas especies han demostrado mayor predisposición por los bosques inundables, particularmente aguajales densos y mixtos y los renacales (asociación de Picus spp.), donde se han reportado altas densidades incluso en áreas de persistente caza (Bodmer et al. 1997, 1999; Aquino et al. 2001, 2003), por coincidencia ausentes en el área de estudio.
Con excepción del machín negro (C. apella), el modelo de cosecha no indica sobrecaza para las especies denominadas de tamaño mediano y pequeño, lo cual coincide con los resultados obtenidos para los bosques inundables aledaños a las comunidades de Parinari y San Miguel (Aquino et al. 2003) y zonas de moderada y ligera caza de la cuenca del río Samiria (Aquino et al. 2001), pero difiere con los resultados obtenidos para el área de estudio de Maipuco, Nueva Esperanza y San Antonio (Bodmer et al., 1999), donde a excepción de C. apella, las demás especies se encontraban sobrecazadas. Este resultado conjuntamente con el de Aquino et al. (2001, 2003), indican que por el momento las especies de tamaño pequeño son las únicas que estarían soportando la presión de caza, al menos en lo que corresponde al nororiente peruano.
La diversidad de primates y de otros componentes de la fauna silvestre que habitan en la cuenca del río Alto Itaya está sujeto a las constantes amenazas que derivan de las actividades del hombre, muchas veces poniendo en grave riesgo su existencia. De acuerdo con las observaciones realizadas in situ, las amenazas potenciales son la caza y la deforestación y en el corto plazo también lo serán las exploraciones sísmicas con miras a la búsqueda y explotación de hidrocarburos por cuanto la cuenca del Alto Itaya se encuentra dentro del Lote 124 adjudicado a la Compañía petrolera Burlington Resources.
La caza es la principal amenaza potencial, actividad que mayormente tiene sus efectos negativos en poblaciones de primates considerados de tamaño grande; así lo demuestra este estudio y muchos otros realizados dentro y fuera de la Amazonía peruana (Bodmer et al. 1988, 1999; Peres 1990; Aquino et al. 2003). La caza no solamente es practicada por las comunidades asentadas, sino también por los denominados madereros, quienes indirectamente se han convertido en amenaza potencial para la sobrevivencia de los primates y de otros componentes de la fauna silvestre.
Las concesiones forestales otorgadas en la cuenca del río Alto Itaya por el Instituto Nacional de Recursos Naturales (INRENA) en abierta contradicción a los dispositivos legales vigentes de prohibición de la tala del bosque en cabeceras de cuencas han iniciado sus operaciones de extracción de madera de valor comercial a inicios del 2005. Los denominados madereros, afín de abaratar costos de operación, están recurriendo a la fauna silvestre para obtener la fuente de proteína animal, por lo que para garantizar la provisión diaria de carne de monte, cada brigada cuenta con cazadores profesionales, quienes cazan a los primates debido a que son más vulnerables por su relativa facilidad para descubrirlos. Es indudable que la escasa presencia de primates en los bosques cercanos al curso del río Itaya y de sus afluentes tiene mucha relación con la presencia de madereros, y de continuar la actividad madedera no cabe duda que en el mediano plazo habrán exterminado las poblaciones, incluso de los denominados de tamaño mediano y pequeño entre los que se encuentran dos especies con distribución geográfica restringida: el huapo negro (P. aequatorialis), con distribución al norte del río Marañón, entre la margen izquierda del río Tigre y margen derecha de los ríos Curaray y Napo (Hershkovitz 1987) y el tocón negro (C. torquatus) (Fig. 2), con distribución al norte del río Marañón, entre la margen derecha del río Nanay y margen izquierda del río Tigre, siguiendo la orilla izquierda hacia el norte hasta la quebrada Alemán, afluente izquierdo del río Pucacuro donde termina la vegetación de tipo varillal (Aquino, obs. pers.). Es oportuno recalcar que los ejemplares del tocón negro que habitan en esta parte de la Amazonía peruana presentan marcadas diferencias fenotípicas con respecto a los que habitan entre las cuencas de los ríos Napo y Putumayo, por lo tanto, no correspondería a C. torquatus lucifer, sino más bien podría tratarse de un nuevo taxa, por lo que recomendamos los análisis de cariotipo y ADN para ambas poblaciones.

La deforestación constituye la segunda amenaza para la supervivencia de los primates. La tala y destrucción de los bosques está en estrecha relación con la actividad agrícola y de extracción de madera comercial; de ellas, la agrícola se puede considerar como la más importante, por cuanto se van asentado cada vez más grupos organizados hasta muy cerca de la cabecera del Itaya donde ya se han talado algunas hectáreas de bosques para convertirlos en campos de cultivo de pan llevar. La deforestación también incluye la tala de árboles cuyos frutos constituyen recursos alimenticios para los primates, pero que el hombre en abierta competencia los cosechan de manera inapropiada para su expendio, los mismos que tienen gran demanda y aceptación en los mercados de Iquitos, particularmente leche huayo (Couma macrocarpa Barb. Rodr.), yahuar huayo (Rhigospira sp.) y masarandúa (Micropholis sp.).
Si bien la exploración sísmica aún no se ha dado, ésta se convertirá en la principal amenaza potencial cuando inicien sus actividades el 2009, por cuanto el tráfico aéreo, generación de ruidos, presencia de fuerza laboral, apertura de helipuertos, zonas de descarga e instalación de campamentos, apertura de líneas sísmicas, detonación de explosivos y muchas otras actividades relacionadas con la exploración sísmica tendrán sus efectos generando ahuyentamiento y alteraciones en los desplazamientos de los primates y de la fauna terrestre y arborícola en general, cuya consecuencia podría ser una drástica disminución de las poblaciones de primates y de otros componentes de la fauna silvestre en la mencionada cuenca.
Frente a estas amenazas urge la elaboración y aplicación de un plan de manejo integral que mitigue las amenazas, caso contrario las especies con mayor riesgo de extinción correspondería a los primates, entre ellos el tocón negro (C. torquatus), quien tiene como hábitat específico los varillales, los cuales se encuentran de manera dispersa en el área de estudio. Estos varillales ya están siendo seriamente afectados por la extracción del aceite caspi (Caraipa sp.), especie muy cotizada para construcciones de viviendas por su resistencia a las polillas.
Agradecimiento
Nuestro agradecimiento al Centro Amazónico para la Educación Ambiental e Investigación (ACEER) y a su representante Aura Murrieta por el apoyo económico oportuno. Al Consejo Superior de Investigaciones de la Universidad Nacional Mayor de San Marcos por el cofinanciamiento. Nuestra gratitud a las autoridades políticas y moradores de las comunidades 12 de Enero, Carbajal, Luz de Oriente y Villa Belén por su activa participación y facilidades otorgadas durante las actividades de campo. Nuestra gratitud a los asistentes de campo con quienes compartimos gratas experiencias durante las exploraciones al interior del bosque.
Literatura citada
Alvarez, J.A. & P. Soini. 2000. Evaluación de la fauna silvestre del área de influencia carretera Iquitos – Nauta. Informe Final del Subproyecto. Instituto de Investigaciones de la Amazonía Peruana. Iquitos, Perú..
Aquino, R. & F. Encarnación. 1994. Primates of Peru/Los Primates del Perú. Primate Report 40: 1 – 127.
Aquino, R.; R. E. Bodmer & E. Pezo. 2000. Evaluación de primates en la cuenca del río Pucacuro, Amazonía peruana. En: La Primatología en el Perú, Vol. II. F. San Martín y M. García (eds), pp. 92 – 100. Lima, Perú.
Aquino, R.; R. E. Bodmer & J. G. Gil. 2001. Mamíferos de la cuenca del río Samiria: Ecología Poblacional y Sustentabilidad de la Caza. Imprenta Rosegraff, Lima, Perú.
Aquino, R. & A. Calle. 2003. Evaluación del estado de conservación de los mamíferos de caza: Un modelo comparativo en comunidades de la Reserva Nacional Pacaya Samiria (Loreto, Perú). Rev. Per. biol. 10 (2): 163 – 174.
Aquino, R.; Terrones, C.; Navarro, R. y W. Terrones. 2007. Evaluación del impacto de la caza en mamíferos de la cuenca del río Alto Itaya, Amazonía peruana. Rev. Per. biol. 14(2): 181 – 186.
Bodmer., R. E.; T.G. Fang & L. Moya. 1988. Primates and ungulates: A comparison in susceptibility to hunting. Primate Conservation 9: 79 – 83.
Bodmer, R. E.; R. Aquino & P. Puertas. 1997. Alternativas de manejo para la Reserva Nacional Pacaya Samiria: Un análisis sobre el uso sostenible de la caza. En: Manejo de Fauna Silvestre en la Amazonia. T. Fang, R. Bodmer, R. Aquino y M. Valqui (eds), pp. 65 - 74, Quito, Ecuador.
Bodmer, R.; C. Allen ; J. Penn; R. Aquino & C. Reyes. 1999. Evaluación del Uso Sostenible de la Fauna Silvestre en la Reserva Nacional Pacaya Samiria. Documento de Trabajo América Verde 4b. The Nature Conservancy.
Bodmer, R. E. & E. Pezo. 2000. Análisis económico el uso de fauna silvestre en la Amazonía Peruana. En: Manejo y Conservación de Fauna Silvestre en América Latina. T. G. Fang, O. L. Montenegro y R. E. Bodmer (eds), pp. 171 – 182. La Paz, Bolivia.
Burnham, K. P.; D. R. Anderson & J. L. Laake. 1980. Estimation of density from line transects sampling of biological populations. Wildlife Monographs 72.
Defler, T. R. 1979. On the ecology and behavior of Cebus albifrons in eastern Colombia. Ecology 20: 475 – 490.
Harvey, P. H.; H. D. Martin & T. H. Cluton-Broock. 1987. Life histories in comparative perspective. In: Primates Societies. B. B.Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham and T. T. Struhsaker (eds), 191 – 196. University of Chicago Press, Chicago.
Hershkovitz, P. 1977. Living New World Monkeys (Platyrrhini). With an Introduction to Primates, Vol. 1. University of Chicago Press, Chicago.
Hershkovitz, P. 1987. The taxonomy of south american sakis, genus Pithecia (Cebidae: Platyrrhini): A preliminary report and critical review with the description of a new species and a new subspecies. Am. J. Primatol. 12: 387 – 468.
Hershkovitz, P. 1988. Origen, speciation and distributios of South American titi monkeys, genus Callicebus (Family Cebidae, Platyrrhini). Proc. Acad. Nat. Sci. Philadelphia. 140: 240 – 272.
Ique, C. 1990. estudio de la bioecología de Saimiri sciureus en la Isla de Iquitos, Loreto, Perú. En: La Primatología en el Perú. N. Castro (ed), pp. 489 – 505. Lima, Perú.
Izawa, K. 1976. Groups size and composition of monkeys in the upper Amazon basin. Primates 17: 376 – 399.
Kinzey, W.G. 1981. The Titi monkey, genus Callicebus. In: Ecology and behavior of neotropical primates. A. Coimbra-Filho and R. Mittermeier (eds), pp. 241 - 276. Academia Brasileira de Ciencias, Río de Janeiro.
Peres, C. 1990. Effects of hunting on western Amazonian primate communities. Biological Conservation 54: 47 – 59.
Rylands, A.B.; R. A. Mittermeier & E. Rodriguez-Luna, 1995. A species list for the New World primates (Platyrrhini): Distribution by country , endemism, and conservation status according to the Mace-Lande sistem. Neotropical Primates 3 (Suppl.): 113 – 160.
Van Roosmalen, , M.G.M.; T. Van Roosmalen & R. A. Mittermeier. 2002. A taxonomic review of the titi monkey, genus Callicebus Thomas. 1903, with the description of two new species, Callicebus bernhardi and Callicebus sthephennashi from Brazilian Amazonia. Neotropical Primates 10: 1 – 46.
Presentado: 26/05/2008
Aceptado: 03/08/2008
Publicado online: 26/02/2009
