










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.17 n.1 Lima abr. 2010
TRABAJOS ORIGINALES
Clave de géneros de larvas de Trichoptera (Insecta) de la Vertiente Occidental de los Andes, Lima, Perú
Genera key to Trichoptera (Insecta) larvae from Western slope of the Andes, Lima, Peru
Ana A. Huamantinco y Willington Ortiz
Laboratorio de Ecología, Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Apartado 11-0058, Lima 11, Perú.
Email Ana Huamantinco: ahuamantincoa1@unmsm.edu.pe
Email Willington Ortiz: wil_om07@hotmail.com
Resumen
Se presenta una clave para identificación de larvas de Trichoptera en el último estadio de desarrollo. El material biológico fue colectado en quebradas situadas en la cuenca media y alta de ríos del departamento de Lima, Vertiente Occidental de los Andes. Se presenta una sinopsis de los géneros y especies para el Perú de las familias encontradas en este estudio.
Palabras clave: insectos acuáticos, taxonomía, Andes occidentales.
Abstract
A key for identification of Trichoptera larvae of the last development instar is proposed. The biological material was collected in streams located in the middle and high river basins of Lima, Western Slope of the Andes. A synopsis for genera and species for Peru is presented for each family found in this study.
Keywords: Aquatic insects, taxonomy, western Andes.
Introducción
Las larvas del orden Trichoptera son importantes componentes de la fauna de invertebrados bentónicos de ríos y quebradas. Cumplen un importante rol intermediario en las cadenas tróficas de ríos y arroyos (Wiggins & Mackay 1978). En las últimas décadas han recibido mayor atención debido a que diversos estudios han demostrado su sensibilidad a las alteraciones ambientales, en especial a contaminación orgánica, lo que las hace buenas indicadoras de calidad de agua (Rosenberg & Resh 1993, de Moor 1999). Recientemente han sido estudiadas como indicadoras de la degradación estructural en cursos de agua influenciados por prácticas agrícolas inadecuadas y por deforestación (Chakona et al. 2009).
Existen aproximadamente 13000 especies de Trichoptera a nivel mundial (Holzenthal et al. 2007) y 2196 especies en la Región Neotropical (Flint et al. 1999). La región Oriental y Neotropical muestran la mayor diversidad de especies, evidenciando que este orden diversifico ampliamente en regiones tropicales sobretodo montañosas y de alta pluviosidad. En relación a endemismos de géneros de Trichoptera, la región Neotropical ocupa el segundo lugar a nivel mundial con 69% de géneros endémicos (de Moor & Ivanov 2008).
Pocos estudios de la fauna de Trichoptera han sido realizados en el Perú. El primero fue el realizado por Martynov (1912), donde describe 24 especies, 12 de ellas nuevas para la ciencia. Posteriormente, Flint (1975, 1980) publicó el registro de especies de las expediciones realizadas al sur del Perú. El único trabajo sobre Trichoptera de la Vertiente occidental de los Andes del Perú, La libertad, fue realizado por Flint y Reyes (1991). En 1996, Flint publico el mayor trabajo de registro de especies de Trichoptera para el Perú donde reporta 77 especies de Cuzco y Madre de Dios.
Flint, Holzenthal & Harris (1999) publican el primer catalogo de Trichoptera para la región Neotropical, en este trabajo consignan 185 especies para Perú. Posteriores publicaciones (Calor & Holzenthal 2008; Harris & Flint 2002; Harris et al. 2002a 2002b; Prather 2003, 2004; Robertson & Holzenthal 2008) incrementan el numero de especies a 210, ubicadas en 47 géneros y 11 familias. La mayoría de las especies registradas fueron del sudeste de Perú.
La presente clave pretende servir de ayuda para los interesados en la entomofauna acuática como base en estudios de taxonomía y ecología de éste orden, así como también para quienes hacen evaluaciones de calidad de agua basados en macroinvertebrados acuáticos.
Morfología y biología de las larvas
Las larvas de Trichoptera presentan pequeño tamaño, entre pocos milímetros hasta 30 mm. Tienen la cabeza esclerosada y presentan tres pares de patas torácicas, estos dos caracteres sin embargo pueden confundirla con larvas de Coleoptera. El carácter diagnostico más importante para larvas de Trichoptera es la presencia de un par de falsas patas anales (Wiggins 1996), que según las familias, muestran diversos tamaños pero siempre están presentes. El pronoto siempre está esclerosado mientras que el mesonoto y metanoto presentan grado variable de esclerosación, cuando las branquias están presentes se ubican a veces en el tórax pero casi siempre en el abdomen, que es membranoso.
Las larvas de Integripalpia construyen estuches transportables y se desplazan siempre con él, usualmente son tubulares hechos de granos de arena, piedritas o material vegetal. Las larvas de Annulipalpia construyen refugios con una red de captura, el refugio suele tener forma de embudo o de bolsa y está construido de arena y material vegetal. Muy importante es la presencia de la red de captura con abertura de malla de variados tamaños, donde es atrapado el detrito y organismos pequeños transportados por la corriente. La larva pasta, cada cierto tiempo, el material atrapado. Un tercer grupo de larvas Spicipalpia presentan familias que construyen estuches fijos o transportables, mientras que otras son de vida libre y apenas tejen un hilo de seda para anclarse y evitar ser arrastradas por la corriente.
Las larvas de la familia Hydroptilidae (Spicipalpia), los Trichoptera más pequeños (alrededor de 1,5 - 5,0 mm ), presentan un desarrollo hipermetamórfico (Angrisano 1995), por el cual, sólo construyen estuches transportables o adheridos al substrato, en el último estadio larval (quinto). Además durante ésta última etapa larval, agrandan el abdomen y disminuyen el tamaño de las falsas patas anales tomando una apariencia diferente a la de los estadios precedentes.
Las larvas de Trichoptera suelen preferir ambientes lóticos en ríos y arroyos de montaña, pero también se les encuentra en las pozas de los ríos o en lagunas y pantanos.
Sinopsis de las familias y géneros encontrados en la Vertiente Occidental de los Andes, Lima
En colectas realizadas en la sierra de Lima, entre 2172 - 3700 m de altitud, en quebradas de pequeño orden, fueron identificados diez géneros de Trichoptera: Helicopsyche Siebold (Helicopsychidae), Mortoniella Ulmer (Glossosomatidae), Atopsyche Banks (Hydrobiosidae), Cailloma Ross & King (Hydrobiosidae), Smicridea McLachlan (Hydropsychidae), Metrichia Ross (Hydroptilidae), Neotrichia Morton(Hydroptilidae), Nectopsyche Müller (Leptoceridae), Anomalocosmoecus Schmid (Limnephilidae) y Chimarra Stephens (Philopotamidae).
La familia Glossosomatidae es cosmopolita, según Holzenthal et al. (2007) el estudio de la diversidad de esta familia en la región Neotropical es todavía muy incompleto. Prácticamente los 13 géneros neotropicales son endémicos de ésta región. Las larvas construyen estuches transportables con forma de caparazón de tortuga y se alimentan raspando el perifiton de la superficie de las rocas. Estuvieron registrados tres géneros en Perú (Flint et al. 1999): Mexitrichia, Protoptila y Mortoniella. Recientemente Blahnik & Holzenthal (2008) sinonimizaron Mexitrichia a Mortoniella.
Mortoniella está distribuido desde México hasta Argentina y tiene registradas cuatro especies para Perú (Flint et al. 1999; Blahnik & Holzenthal 2008). La larva considerada en esta clave presenta una seta muy engrosada en la base de la uña tarsal anterior. Este es un carácter apomorfico que podría pertenecer solamente al grupo de especies de Mortoniella bilineata Ulmer 1906 o a todo el género (Blahnik & Holzenthal 2008) esta confirmación aguarda mas estudios de asociación larva-adulto. Estas larvas son escasas en las quebradas estudiadas, fueron encontradas en áreas de rápidos en substrato de piedras de diverso tamaño.
La familia Helicopsychidae es una familia primariamente tropical. El género Helicopsyche es de distribución cosmopolita. Todas las larvas conocidas construyen estuches transportables con forma de concha de caracol y usan en su construcción granos de arena. Probablemente sean raspadoras de perifiton en quebradas tanto en rápidos como en pozas. Para el Perú se han descrito 7 especies de éste género. En éste estudio las larvas fueron encontradas en rápidos y pozas, en substrato de piedras pequeñas a medianas.
La familia Hydrobiosidae presenta larvas de vida libre y depredadoras. Muy característico en ellas es la pata anterior que está modificada para formar una quela, útil para sujetar la presa. Para el Perú se han registrado dos géneros Atopsyche y Cailloma.
Atopsyche es el género con mayor número de especies en la familia, en el Perú se han registrado 11 especies de éste género. En el presente estudio se encontraron gran número de estas larvas en áreas de rápidos tanto en musgos como en piedras.
Según Flint (1974) el género Cailloma es el único Hydrobiosidae distribuido tanto en la subregión Chilena como en la Brasileña donde pertenece Perú. Solo ha sido descrita una especie de Cailloma para el Perú. Las larvas fueron encontradas en área de rápidos en substrato piedras ocupando el mismo hábitat que Atopsyche pero en menor número.
La familia Hydropsychidae es de distribución mundial, presenta una gran cantidad de especies, ocupa el tercer lugar en razón al número de especies descritas (Holzenthal et al. 2007). Son típicas de áreas de rápidos y a menudo muy abundantes. En el Perú se han registrado 6 géneros de esta familia: Centromacronema, Leptonema, Macronema, Macrostemum, Smicridea y Synoestropsis. El género Smicridea es uno de los más ricos en especies, construye refugios de seda con una red de captura para obtener su alimento. Para el Perú se han registrado 16 especies de éste género. Las larvas en éste estudio, fueron encontradas en áreas de rápidos con similar preferencia por substratos de musgos y piedras.
La familia Hydroptilidae se encuentra en todas las regiones faunales del mundo. Se han registrado 1679 especies de ésta familia, siendo la más diversa dentro del orden (de Moor & Ivanov 2008). Construyen refugios de seda o arena en el último estadio larval. Para el Perú se han registrado 13 géneros de ésta familia: Alisotrichia, Bredinia, Ceratotrichia, Costatrichia, Flintiella, Hydroptila, Metrichia, Neotrichia, Ochrotrichia, Orthotrichia, Oxyethira, Rhyacopsyche y Zumatrichia.
El género Metrichia fue considerado hasta hace poco, un subgénero de Ochrotrichia debido a que aparentemente carecía de diagnosis larval congruente. Wiggins (1996) propone una diagnosis para este género, donde se definen caracteres que son usados en éste trabajo. Las larvas se encuentran en áreas de rápidos y asociadas a algas filamentosas, construyen un estuche aplanado oval de seda asociado con arena o material vegetal. Para el Perú este género tiene registradas 7 especies. En el presente estudio, se encontraron abundantes larvas en zonas de rápidos tanto en piedras como musgos, pero con una marcada preferencia por éste último substrato donde suelen anclar sus estuches pupales.
Neotrichia, es uno de los géneros neotropicales con mayor número de especies. Las larvas construyen pequeños estuches tubulares de granos de arena y viven en rápidos de quebradas o ríos. Para el Perú se han descrito 9 especies de éste género. En este estudio se encontraron pocas larvas de éste género, se les colectó tanto en piedras como en musgos en zonas de rápidos.
La familia Leptoceridae es de distribución cosmopolita y ocupa el segundo lugar en número de especies (1549) dentro del orden (de Moor & Ivanov 2008). Las larvas construyen estuches tubulares de variedad de materiales minerales y vegetales o completamente de seda y se alimentan principalmente como detritívoros u omnívoros. Para Perú se han registrado 9 géneros de ésta familia: Achoropsyche, Amphoropsyche, Grumichella, Nectopsyche, Notalina, Oecetis, Osflintia, Triaenodes y Triplectides.
El género Nectopsyche es rico en especies, restringidas al nuevo mundo. Las larvas habitan ambientes tanto loticos como lenticos, se les clasifica según su modo de alimentación, como cortadoras y colectoras de deposito. Este género tiene registradas 12 especies en Perú. En el presente estudio, las larvas fueron encontradas en áreas de rápidos y pozas siendo mayor la abundancia en este último hábitat.
La familia Limnephilidae, es el grupo dominante en la mayor parte del Hemisferio Norte, muestra su mayor diversidad en las áreas temperadas de la región Holartica y es también la familia de Trichoptera con mayor diversidad ecológica (Holzenthal et al. 2007). En el Perú solo se ha registrado el género Anomalocosmoecus que es endémico de Sudamérica y conocido de grandes elevaciones en los Andes, desde Argentina hasta Colombia (Flint 1982). En Perú, se conocen tres especies de éste género. En este estudio, se evidenció que su distribución parece estar fuertemente influenciada por la altitud, se les encontró en rápidos con substrato de piedras.
La familia Philopotamidae se encuentra en todas las regiones del mundo, la larva construye redes tubulares largas usualmente ancladas a objetos por debajo de la superficie, estas redes filtran materia orgánica fina del agua la cual le sirve de alimento. Para Perú se han descrito 3 géneros de esta familia: Chimarra, Chimarrhodella y Wormaldia.
El género Chimarra se encuentra en todas las regiones del mundo, es uno de los géneros más grandes en el orden Trichoptera (Holzenthal et al. 2007). Para el Perú se han descrito 23 especies de Chimarra. En éste estudio, se encontraron pocos ejemplares de estas larvas, colectadas en rápidos tanto en musgo como en piedras.
En esta sinopsis se han considerado las ocho familias encontradas en la zona de estudio. Existen además otras tres familias registradas para el Perú Anomalopsychidae, Calamoceratidae y Polycentropodidae.
Las larvas de Anomalopsychidae se caracterizan porque la cabeza posee una carena que rodea el vertex hasta el margen anterior del frontoclipeo. En el pronoto también existe una carena cerca al margen posterior, que se prolonga anterolateralmente como un lóbulo redondeado o en punta. La uña anal posee numerosos dientes accesorios ubicados en un lóbulo que emerge de la uña. Construyen estuches transportables tubulares (Holzenthal & Flint, 1995). Para el Perú solo ha sido registrado el género Contulma (Flint et al., 1999).
Las larvas de Calamoceratidae se caracterizan porque el labro presenta una hilera transversal de aproximadamente 16 setas largas y gruesas. El pronoto presenta a veces lóbulos anterolaterales prominentes, mientras que el mesonoto esta mayormente cubierto por placas esclerosadas usualmente pigmentadas. El abdomen presenta branquias individuales o ramificadas y algunos géneros se distinguen por una banda de finos filamentos laterales. Construyen estuches transportables de material vegetal (Wiggins, 1996). En el Perú han sido registrados los géneros Banyallarga y Phylloicus (Flint et al., 1999).
En Polycentropodidae, las larvas se distinguen por que el trocantin anterior es puntiagudo y fusionado al episterno. Sólo el pronoto esta esclerosado. Construyen redes de captura con forma de embudo (Wiggins, 1996). Cuatro géneros de esta familia han sido registrados para el Perú Cernotina, Cyrnellus, Polycentropus y Polyplectropus (Flint et al., 1999).
Material y métodos
El material biológico para el estudio fue colectado en diversas quebradas de la sierra de Lima (2172-3700 m altitud), sin aparente impacto antropogénico. Las quebradas de menor altitud, se encuentran en la zona de vida Estepa espinosa - Montano Bajo Tropical (ee-MBT) caracterizada por la presencia de valles y laderas de la vertiente occidental entre 2000 - 3000 m de altitud. El clima es semiárido - templado cálido con temperatura media anual 17 - 12 °C y promedio anual de precipitación pluvial de 250 - 450 mm. La vegetación típica es de herbáceas, arboles como Schinus molle y cactáceas. Durante la época de lluvias veraniegas emergen hierbas efímeras, que se asocian con la vegetación arbustiva y algunas cactáceas que existen permanentemente. Las quebradas de mayor altitud se encuentran en la zona de vida Estepa - Montano Tropical (e-MT) ubicada entre 3000 -4000 m. El clima es subhúmedo - templado frio con temperatura media anual de 12 - 6°C y promedio anual de precipitación pluvial de 350 - 500 mm. La cobertura vegetal consiste de vegetación graminal de pradera altoandina asociado con cactáceas del género Opuntia (Inrena, 2000). Las colectas fueron realizadas entre el año 2005 al 2009, en los siguientes sitios de estudio: tributario de segundo orden 18L 0281443 UTM 8845669, 3700 m y de tercer orden 18L 0278280 UTM 8850251, 2902 m en la cuenca del río Pativilca (Cajatambo). Tributario de tercer orden 18L 0351835 UTM 8577827, 2912 m; tributario de segundo orden 18L 0343270 UTM 8684690, 2172 m y tributario de primer orden 18L 0352434 UTM 8688316, 3144 m en la cuenca del río Rímac (Huarochirí). Tributario de tercer orden 18L 0347841 UTM 8674068, 3010 m en la cuenca del río Lurín (Huarochirí). Los tributarios de segundo orden de la cuenca del Rímac y de tercer orden de la cuenca del Lurín fueron muestreados mensualmente durante un año, mientras que los otros tributarios lo fueron estacionalmente también por un periodo anual.
Fueron analizadas 8,520 larvas de Trichoptera de substrato preferentemente piedras en áreas de rápidos, eventualmente también se colectó en correderas y pozas y en substratos de naturaleza orgánica como hojas retenidas en la corriente, musgos, raíces de berro y algas. Se uso una red Surber (180 µm, 30x30 cm) y el material fue fijado en alcohol al 80%. Fueron observados, siempre que fue posible, un mínimo de diez especímenes de cada género.
Los caracteres usados en esta clave se seleccionaron atendiendo a su fácil visualización y definición y están basados en los trabajos de Wiggins (1996), Flint (1963, 1974) y Fernández & Domínguez (2001). Para las ilustraciones, los especímenes completos y estructuras de valor taxonómico, fueron fotografiados con una cámara digital acondicionada a un microscopio estereoscópico o compuesto. El trazo final en tinta china, fue realizado sobre una mesa de luz en papel Canson de dibujo colocado sobre la foto impresa.
Clave para familias de larvas de Trichoptera
1 Estuche transportable de forma helicoidal. Uña anal con varios dientes que le da un aspecto de peine. Fuera del estuche, la larva presenta el abdomen enrollado dorsoventralmente. Helicopsychidae
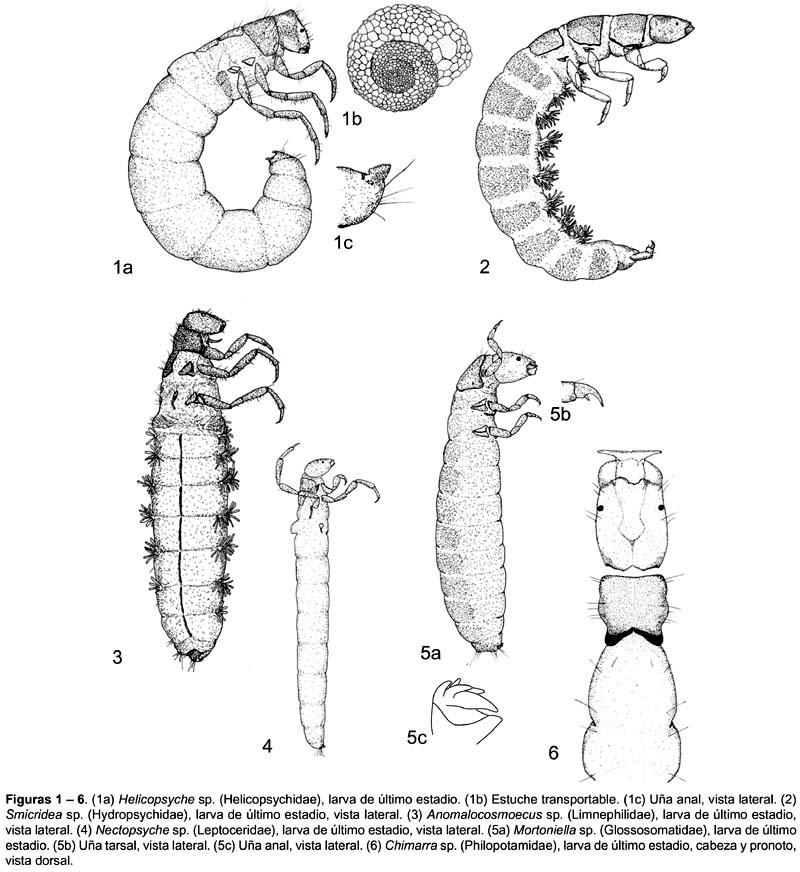
Helicopsyche. Mandíbulas fuertes de forma triangular. Pro y meso noto con placas esclerosadas, metanoto con pequeñas escleritas débilmente pigmentadas. Placa de pequeñas espinas a ambos lados del primer segmento abdominal (Fig.1).
Estuche transportable no helicoidal o no construye estuche 2
2 (1) Los tres notos torácicos esclerosados 3
Pronoto esclerosado, meso y meta noto con variados grados de esclerosación. 4
3 (2) Con branquias en la superficie ventral del abdomen, mechón de setas largas cerca al ápice de la pata anal. No construyen estuches transportables pero si refugios con red de captura. Hydropsychidae
Smicridea. Dorso del abdomen con setas aplanadas. Los filamentos branquiales emergen de un tallo central poco evidente (Fig.2).
Branquias ausentes. Larvas pequeñas, 2 - 3 mm longitud. En el último estadio construyen estuches transportables de variadas formas, en la más típica está constituida de dos valvas de seda que pueden incorporar arena u otros materiales del medio. Hydroptilidae
4 (2)Mesonoto con placas esclerosadas, estuche transportable cilíndrico adelgazándose hacia el extremo posterior. 5
Mesonoto sin placas o con pequeñas escleritas poco esclerosadas. Sin estuche transportable, o cuando presente en forma de caparazón de tortuga. 6
5 (4) Cuerno prosternal presente. Abundantes setas gruesas y negras en el primer segmento abdominal. Con branquias abdominales individuales o múltiples. Limnephilidae
Anomalocosmoecus. Cabeza con carena dorsolateral y ápice de la mandíbula sin dientes (Fig.3).
Cuerno prosternal ausente. Patas posteriores más largas que las anteriores. Primer segmento abdominal con escleritas espinosas en las jorobas laterales. Leptoceridae
Nectopsyche. Esclerita espinosa en el primer segmento abdominal con un área circular y barra esclerosada (Fig.4).
6 (4) Pronoto en vista dorsal mas ancho en su parte media. Mesonoto con pequeñas escleritas poco esclerosadas. Mitad basal de pata anal ampliamente unida con el segmento IX. Uña anal con al menos una uña accesoria. Estuche transportable en forma de caparazón de tortuga. Glossosomatidae
Mortoniella. Uña de la pata anal con 4 uñas accesorias. Seta corta lobular en la base de las uñas tarsales (Fig.5).
Pronoto en vista dorsal no es mas ancho en su parte media. Meso y meta noto no esclerosados. Pata anal libre del segmento IX. Uña anal sin uña accesoria. No construye estuche transportable. 7
7 (6)Primer par de patas torácicas forman un apéndice quelado. Son de vida libre. Hydrobiosidae
Primer par de patas torácicas no forman un apéndice quelado. Labro membranoso, que cuando está proyectado tiene forma de T. Philopotamidae
Chimarra. Con una muesca notoria en el margen anterior del apotoma frontoclipeal. Proceso largo, como espina, en la coxa de la pata anterior (Fig.6).
Clave para géneros de larvas de Hydroptilidae
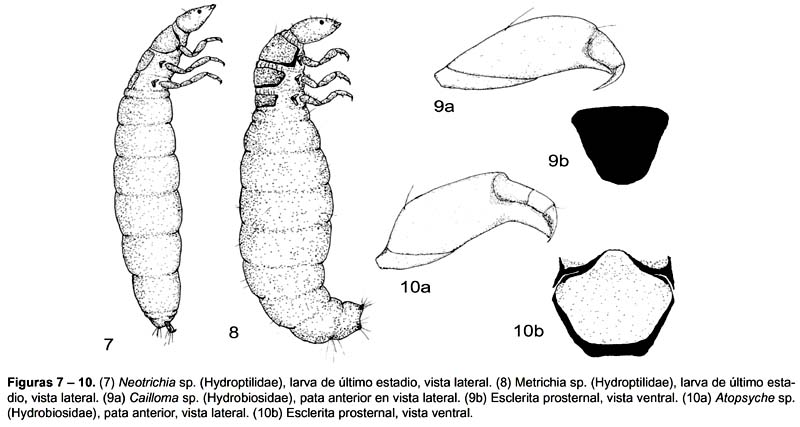
1 Cabeza piriforme alargada anteriormente. Patas torácicas medias y posteriores mas largas que las anteriores, seta basal en todas las uñas tarsales. Patas anales relativamente largas, se proyectan libremente por fuera del cuerpo. Estuche transportable tubular adherido de granos de arena (Fig.7). Neotrichia
Cabeza no alargada. Patas torácicas aproximadamente de la misma longitud, seta basal solo en la uña tarsal media y posterior. Patas anales cortas no proyectadas por fuera del cuerpo. Estuche constituido de dos valvas de seda a veces adherido de arena, algas filamentosas o brácteas de musgos (Fig.8). Metrichia
Clave para géneros de larvas de Hydrobiosidae
1 Tibia y tarso de la pata anterior fusionados, extensión distal del fémur delgada. Esclerita prosternal angosta (Fig.9). Cailloma
Tibia y tarso de la pata anterior no fusionados, extensión distal del fémur ancha. Esclerita prosternal ancha (Fig.10). Atopsyche
Agradecimientos
Agradecemos a Jorge L. Nessimian y Allan P. Santos (Departamento de Zoología, Instituto de Biología, Universidad Federal de Río de Janeiro, Brasil) y a tres revisores anónimos por la lectura crítica del manuscrito. También agradecemos a Diana Alcántara, Juan Carlos Cusi, Rosmery Huaman, Gissela Pascual, Jorge Peralta Argomeda, Ernesto Razuri, Edwin Sifuentes y Darwin Valcárcel, por la ayuda en las colectas.
Literatura citada
Angrisano, E.H. 1995. Insecta Trichoptera. In: E.C. Lopretto y G Tell, eds. Ecosistemas de Aguas Continentales: Metodologias para su estúdio, Tomo III, Identificación de organismos. Ediciones Sur, La Plata. Pp. 1199-1237.
Blahnik R.J. & R.W. Holzenthal. 2008. Revision of the Mexican and Central America species of Mortoniella (Trichoptera: Glossosomatidae: Protoptilinae). Zootaxa 1711: 1-72.
Calor, A.R. & R.W. Holzenthal. 2008. Phylogeny of Grumichellini Morse, 1981 (Trichoptera: Leptoceridae) with the description of a new genus from southeastern Peru. Aquatic Insects 30(4): 245-259.
Chakona A., C. Phiri & J.A. Day. 2009. Potential for Trichoptera communities as biological indicators of morphological degradation in riverine systems. Hydrobiologia 621: 155-167.
de Moor, F.C. 1999. The use of Trichoptera to assess biodiversity and conservation status of South African river systems. In: Proc. 9th Int. Symp. Trichop. Chiang Mai. Pp. 237-244.
de Moor, F.C. & V.D. Ivanov. 2008. Global diversity of Caddisflies (Trichoptera: Insecta) in freshwater. Hydrobiologia 595: 393-407.
Fernandez H.R. & E. Dominguez. 2001. Guía papa la determinación de los artrópodos bentónicos sudamericanos. Serie: Investigaciones de la Universidad Nacional de Tucumán Editorial Universitaria de Tucumán. 282pp.
Flint, O. S., Jr. 1963. Studies of Neotropical caddisflies, I: Rhyacophilidae and Glossosomatidae (Trichoptera). Proc. U.S. Nat. Mus. 114(3473): 453-477.
Flint, O.S., Jr. 1974. Studies of Neotropical caddisflies, XIX: The genus Cailloma (Trichoptera: Rhyacophilidae). Proc. Biol. Soc. Wash. 87(41): 473-484.
Flint, O.S., Jr. 1975. Studies of Neotropical caddisflies, XX: Trichoptera collected by the Hamburg South-Peruvian Expedition. Entomologische Mitteilungen aus dem Zoologischen Museum Hamburg 4 Band Nr.90: 565-573.
Flint, O.S. Jr. 1980. Trichoptera VI. In: S. W. Roback, eds. The results of the Catherwood Foundation Bolivian-Peruvian Altiplano Expedition. Part I Aquatic Insects except Diptera. Proc. Acad. Nat. Sci. Philadelphia. 132: 213-217.
Flint, O.S., Jr. 1982. Studies of Neotropical caddisflies, XXX: Larvae of the genera of South American Limnephilidae (Trichoptera). Smithson. Contr. Zool. 355: 1-30.
Flint, O.S., Jr. 1996. The Trichoptera Collected on the Expeditions to Parque Manu, Madre de Dios, Peru. In: Don Wilson & A. Sandoval, eds. Manu The Biodiversity of Southeastern Peru. Pp. 369-430.
Flint, O.S., Jr. & L.A. Reyes. 1991. Studies of Neotropical caddisflies, XLVI: the Trichoptera of the rio Moche basin, department of La Libertad, Peru. Proc. Biol. Soc. Wash. 104(3): 474-492.
Flint, O.S. Jr., R.W. Holzenthal & S.C. Harris. 1999. Catalog of the Neotropical Caddisflies (Insecta: Trichoptera). Ohio Biological Survey, Columbus, iv+239p.
Harris, S.C. & O.S. Flint, Jr. 2002. New Alisotrichia (Trichoptera: Hydroptilidae) from Central and South America and the Greater Antilles. Proc. Ent. Soc. Wash. 104(1): 195-210.
Harris, S.C., O.S. Flint, Jr. & R.W. Holzenthal. 2002a. Review of the Neotropical genus Flintella (Trichoptera: Hydroptilidae: Stactobiini). J. New York. Entomol. Soc. 110(1): 65-90.
Harris, S.C., R.W. Holzenthal. & O.S. Flint, Jr. 2002b. Review of the Neotropical genus Bredinia(Trichoptera: Hydroptilidae: Stactobiini). Annals of Carnegie Museum. 71(1): 13-45.
Holzenthal, R.W. & O.S. Flint, Jr. 1995. Studies of Neotropical Caddisflies, LI: Systematics of the Neotropical Caddisflies Genus Contulma (Trichoptera: Anomalopsychidae). Smithson. Contrib. Zool. 575: 1-59.
Holzenthal, R.W., R.J. Blahnik, A.L. Prather & K.M. Kjer. 2007. Order Trichoptera Kirby, 1813 (Insecta), caddisflies. Zootaxa 1668: 639-698.
Inrena (Instituto Nacional de Recursos Naturales). 2000. Base de datos de recursos naturales e infraestructura para el desarrollo socioeconómico del Perú (Departamento de Lima). Dirección general de estudios y proyectos de recursos naturales. Ministerio de Agricultura. 119pp.
Martynov, A.B. 1912. On two collections of Trichoptera from Peru. Annu. Mus. Zool. Acad. Imp. Sci. St Petersbourg 17: 1-40.
Prather, A.L. 2003. Revision of the Neotropical caddisfly genus Phylloicus (Trichoptera: Calamoceratidae). Zootaxa 275: 1-214.
Prather, A.L. 2004. Revision of the Neotropical caddisfly genus Banyalarga (Trichoptera: Calamoceratidae). Zootaxa: 435: 1-76.
Robertson, D.R. & R.W. Holzenthal. 2008. Two new species and a new record of Protoptila from Bolivia (Trichoptera: Glossosomatidae: Protoptilinae). Ann. Entomol. Soc. Am. 101(3): 465-473.
Rosenberg, D.M. & V.H. Resh. 1993. Freshwater biomonitoring and benthic macroinvertebrates. Chapman & Hall. New York. 488pp.
Wiggins, G.B. & R.J. Mackay. 1978. Some relations between systematics and trophic ecology in Neartic aquatic insects with special reference to Trichoptera. Ecology 59(6): 1211-1220.
Wiggins, G.B. 1996. Larvae of the North Americam caddisfly genera (Trichoptera). 2nd ed. University of Toronto Press. xiii+457pp.
Trabajo presentado a la XVIII Reunión Científica del Instituto de Investigaciones en Ciencias Biológicas Antonio Raimondi, 200 años del nacimiento de Charles Darwin y el 150 aniversario de la publicación de On the Origin of Species by Means of Natural Selection. Del 19 al 21 de agosto de 2009.
Publicado impreso: 20/10/2010
Publicado online: 29/09/2010
