










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.17 no.2 Lima Aug. 2010
Camera trap survey of medium and large mammals in a montane rainforest of northern Peru
Evaluación de mamíferos medianos y grandes mediante trampas cámara en un bosque montano del norte del Perú
Carlos F. Jiménez1 ; Heidi Quintana1,3 ; Víctor Pacheco1,2 ; Derek Melton3 ; Javier Torrealva3 and Guillermo Tello4
1 Museo de Historia Natural, Departamento de Mastozoología, Universidad Nacional Mayor de San Marcos, Apartado 140434, Lima 14, Perú.
2 Facultad de Ciencias Biológicas Universidad Nacional Mayor de San Marcos.
3 Golder Associates Perú S.A.
4 Medio Ambiente-Río Tinto
Email Carlos Jiménez: cfja-80@yahoo.com
Abstract
Camera traps are a powerful tool for inventorying elusive and rare species and very useful to obtain ecological data for plans that involve wildlife conservation. In Peru, several surveys have been carried out in lowland Amazonia especially in the southeastern part of the country, but none in montane cloud forests or Yungas. We present the first camera trap studies produced in Peruvian Yungas at the locality of Querocoto village (Chota, Cajamarca), based on 2002 (dry season) and 1264 (wet season) camera traps-days (CTD). Two localities were surveyed in wet and dry season: The Pagaibamba Protection Forest and the San Lorenzo Forest. The wet season study was carried out in October and November, and the dry season in July to September of 2008. Eight mammalian species were recorded in both seasons. Some 66 (91.7%) independent records were obtained in the dry season, but only six (8.3%) in the wet one, suggesting a seasonality effect. The Mountain Paca Cuniculus taczanowskii was the most commonly photographed species, with 17.0 and 1.6 capture frequencies (dry and wet season respectively), whereas the Long-tailed weasel Mustela frenata (0.5 capture frequency in the dry season) was the most rare species. Activity patterns suggest that Mountain Paca C. taczanowskii and the Andean Skunk C. chinga are nocturnal, while Spectacled Bear T. ornatus and Tayra E. barbara are diurnal in the study area. Our records of the Ocelot Leopardus pardalis and the Tayra E. barbara are among the highest altitudinal records known for each species. In addition, the Anta Tapirus pinchaque was also identified by its tracks, representing one of the first record known south of the Huancabamba Depression.
Keywords: Camera traps, inventory, montane cloud forest, Cuniculus taczanowskii, Huancabamba Depression, Peru.
Resumen
Las trampas cámara son una herramienta muy poderosa en el registro de mamíferos raros y elusivos; muy útiles en la obtención de datos ecológicos necesarios para formular planes que involucren la conservación de la fauna. Estos estudios principalmente se han realizado en la Amazonía del Perú especialmente en la parte sur oriental; pero muy pocos en los bosques montanos o yungas. Nosotros presentamos uno de los primeros estudios en las yungas peruanas realizado con trampas cámaras; se llevó a cabo en la localidad de Querocoto (Chota, Cajamarca) con un esfuerzo de muestreo de 2002 (estación seca) y 1264 (estación húmeda) trampas cámara-día (TCD). Dos lugares fueron evaluados en las estaciones húmeda y seca: el bosque de protección de Pagaibamba y el bosque de San Lorenzo. El estudio durante la época húmeda fue llevado a cabo entre octubre y setiembre, mientras la evaluación en época seca fue entre julio y setiembre de 2008. En total se registraron ocho especies de mamíferos. En la época seca se obtuvo 66 (91,7%) de los eventos independientes mientras en la época húmeda solamente seis eventos (8,3%) sugiriendo un efecto por estacionalidad. La Paca de Montaña (Cuniculus taczanowskii) fue el mamífero más fotografiado con frecuencias de captura de 17,0 y 1,6 para época seca y húmeda respectivamente mientras que Mustela frenata con 0,5 fue la especie más rara registrada solo en la época seca. Los patrones de actividad sugieren que la Paca de Montaña, C. taczanowskii y el Zorrillo Andino, Conepatus chinga son nocturnos, mientras que el Oso de Anteojos, Tremarctos ornatus y la Tayra Eira barbara son diurnos en el área de evaluación. Nuestros registros del Ocelote Leopardus pardalis y la Tayra E. barbara están entre los registros altitudinales más altos conocidos para ambas especies. En adición el Anta Tapirus pinchaque fue identificado por medio de sus huellas y representa su primer registro al sur de la depresión de Huancabamba.
Palabras clave: Trampas cámara, inventario, bosques de niebla, Cuniculus taczanowskii, Perú.
Introduction
Camera traps are a powerful and non invasive tool for inventorying elusive and rare species, and have become very popular in the last few years. Camera traps have been used successfully in numerous studies of mammals, and potential applications of this field technique in wildlife studies are increasing. This method provides information on population dynamics (Varma et al. 2006), species richness (Shek et al. 2007, Lyra-Jorge et al. 2008), activity patterns (Maffei et al. 2007, Arispe et al. 2008), habitat use (Bowket et al. 2007, Tobler et al. 2009), population density (Trolle & Kery 2005, Ríos-Uzeda et al. 2007, Rowcliffe et al. 2008), and abundance (Silver et al. 2006, Marnewick et al. 2008); which are essential data for wildlife conservation. Several surveys have been carried out in Peru, principally in lowland Amazonia, but none in montane cloud forests. Montane forests are very difficult landscape with strong slope, dense vegetation and have adverse climate condition. Camera traps, however, are not dependent on the environment and the photographs allow an accurate identification of animals. We present the first biodiversity survey of montane mammals in the Peruvian Yungas in the Querocoto village area (Chota, Cajamarca) using camera traps, to complement a standard evaluation and build baseline information for future conservation planning in the area. Cajamarca, with five protected areas, is poorly represented. Besides, these are small in extension except by the Tabaconas-Namballe Sanctuary National and the Hunting Area of Sunchubamba. Some other areas were created for different purposes (e.g. reserved area of Chancay baños). Data from Cutervo National Park indicate that the park is habitat of endangered species of wild fauna, such as the Jaguar Panthera onça (Linnaeus 1758), the Ocelot Leopardus pardalis (Linnaeus 1758), the Spectacled Bear Tremarctos ornatus (Cuvier 1825), the Otter Lontra longicaudis (Olfers 1818), the Wildcat Leopardus colocolo (Molina 1782), the Mountain Tapir Tapirus pinchaque (Roulin 1829); whereas in the Hunting Area of Sunchubamba, the Peruvian White tailed Deer Odocoileus virginianus Zimmermann 1780, the Brazilian Rabbit Sylvilagus brasiliensis (Linnaeus 1758), the Skunk (Conepatus sp.), the Zariguellas (Didelphis sp.), and the Vizcacha Lagidium peruanum Meyen, 1833 are present.
Material and methods
The study area is located near the Querocoto village (Chota Province, Cajamarca Department) (Fig. 1), on the headwaters of the Río Paltic, which flows into the Río Marañón, and south of the Huancabamba Depression. The study was carried out in the Pagaibamba Protection Forest and surrounding forests from 2500 to 3700 m. The main habitat types are the cloud forest and paramo; however, some areas are currently used for agriculture or to raising livestock, especially along the lower margins. Montane cloud forests of the department of Cajamarca are very important due to the existence of endemic species and their high diversity.
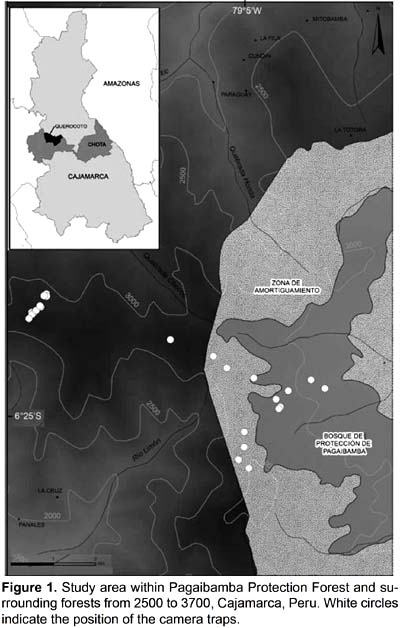
This study was performed during the wet season (October and November 2008) and dry season (July to September 2008) in two localities: The Pagaibamba Protection Forest and San Lorenzo Forest. A set of 25 camera traps (15 Reconyx RC55 and 10 Cuddeback Digital Cam, Expert 3.0 camera traps) were placed on animal trails or where mammals' traces were found. Cameras were set up to an average height of 40 cm above ground, and programmed to take three photos per trigger (Reconyx) with intervals of one second between pictures, or one photo per trigger (Cuddeback). The quiet period or no delay was set to one minute and the sensitivity was also set to high for both. Cameras operated 24h per day recording date and time of each photograph and power was supplied by alkaline batteries. For each Reconyx photograph the temperature and lunar phase were also recorded. Cameras traps were checked twice during the evaluation. The capture frequency was calculated for every given species as the number of photos/1000 camera days (Tobler et al. 2008). To ensure that the events were independent, photographs of the same species and at the same station within a phase of one hour were excluded.
Activity periods were classified following Gómez et al. (2005), with the exception that a crepuscular category was included. Sunset and sunrise hours were determined using Moonrise 3.5 (Sidell 2002). Species were classified as diurnal (<10% of observations in the dark), nocturnal (>90% of observations in the dark), mostly diurnal (between 10 and 30% of observations in the dark), mostly nocturnal (between 70 and 90% of observations in the dark) and crepuscular (50% of observations during the crepuscular phase). The rest of the species were classified as cathemeral (e.g., organism that has sporadic and random intervals of activity during the day or night in which food is acquired).
Results
A sampling effort of 2002 (dry season) and 1264 (wet season) camera traps-day (CTD) was performed in both localities: Pagaibamba Protection Forest and San Lorenzo Forest, gathering 313 photographs of wildlife, representing 72 independent events of medium and large mammals. Other photographs for cattle, small mammals (rodents and marsupials) and birds were obtained too, but were not considered for the purpose of this report. A total of eight mammalian species were recorded (Table 1, Fig. 2), some of them are considered cryptic, rare or poorly known mammals. These include the rodent Mountain Paca Cuniculus taczanowskii (Stolzmann 1865), the Ocelot L. pardalis, the Andean Fox Lycalopex culpaeus (Molina 1782), the Spectacled Bear T. ornatus, the Tayra Eira Barbara (Linnaeus 1758), the Long-tailed Weasel Mustela frenata (Lichtenstein 1831), the Andean Skunk Conepatus chinga (Molina 1782), and the White-tailed Deer O. virginianus.

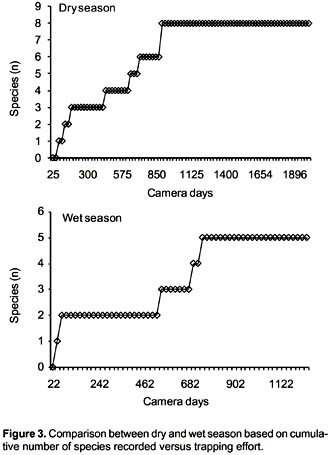
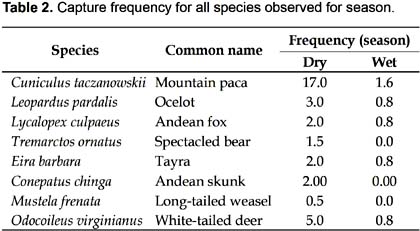
A 75 CTD were employed to record the first mammal during dry season, whereas 44 CTD were needed in wet season. Based on 66 independent events (91.7%) obtained in the dry season (Fig. 3), and only six independent events (8.3%) for the wet season, eight and five species were recorded for the dry and wet season respectively. The species T. ornatus, C. chinga , and M. frenata were not recorded in the wet season. C. taczanowskii was the most frequent recorded species, with 17.0 and 1.6 capture frequency for dry and wet season respectively. On the other hand, the White-tailed deer O. virginianus obtained 5.2 and 0.8 capture frequency for the dry and wet seasons respectively. In case of the Long-tailed weasel M. frenata, with 0.5 capture frequency, this small carnivore obtained the poorest capture frequency, it may be biased for its size (Table 2). Activity patterns for the eight mammals are summarized in Table 3.
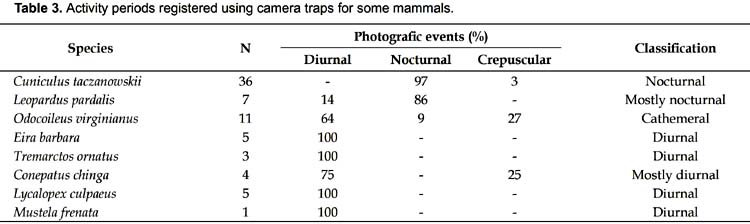
Our results suggest that C. taczanowskii and C. chinga are nocturnal, while M. frenata, L. culpaeus, T. ornatus and E. barbara are diurnal; L. pardalis is mainly nocturnal, whereas O. virginianus is cathemeral in the study area. Two noteworthy records were the Ocelot L. pardalis and the Tayra E. barbara, who were recorded at 3379 m in the Pagaibamba Protection Forest, to our knowledge the highest altitudinal record known for both species.
Other mammals were recorded for signs or interviews during the development of this survey, including the Puma Puma concolor (Linnaeus, 1771), the Anta T. pinchaque, the Pacarana Dinomys branickii Peters 1873, the deer Mazama sp., and the Brazilian Rabbit S. brasiliensis. The record of T. pinchaque, by an unequivocal track recorded on the paramo of Pagaibamba Protection Forest, is highlighted because it represents one of the southernmost records of the species, and south of the Huancabamba Depression. The Brazilian Rabbit was registered by a skin and the puma by scats. The Pacarana and the unknown deer were recorded by interviews only.
Discussion
Eight species of medium and large mammals belonging to three orders were confirmed in the Pagaibamba Protection Forest and surrounding Forests. This result shows that camera traps are highly efficient for inventories in Yunga habitats. In addition, other three species were recorded by indirect signs such as tracks, scats or skins. The Pacarana (D. branickii) and the deer (Mazama sp.) were recorded only by interviews. Both species are expected based on current potential distribution maps (Patterson et al. 2007).
Dry season was the most productive, and appears to be the best season to produce inventory surveys with camera traps. Some mammals such as Ocelots in Central America tended to have larger home range in dry season (Dillon & Kelly 2008). In addition higher values of capture frequency during the dry season appear to indicate an increase of movements by mammals to harvest the scant food resources. These results are in line with those obtained by Stirrat (2003) who demonstrated that the Agile Wallaby Macropus agilis (Gould 1841) increased its home range in the dry season when food quality is scarce.
We obtained lower values of capture frequency in comparisons with survey from lowland forests, which might indicate that mammals in mountain cloud forest have lower densities. For example the Mountain Paca had 17.0 capture frequency while the Brown Agouti Dasyprocta punctata Gray 1842, a related species, had 30.7 and 20.5 capture frequencies in two different evaluations with similar capture effort (Tobler et al. 2008); in spite the first species is bigger. Kelly and Holub (2008) showed the relation between body size and photo rates, mentioning that small animals are less detected and have a lower capture probability, explaining the low frequency of the Long-tailed weasel in our survey.
Activity periods for Ocelot, Spectacled bear, White-tailed Deer, Andean Skunk, Mountain Paca, and Tayra were similar to those reported previously in the literature (Peyton 1980, Ludlow & Sunquist 1987, Castro 1993, Sheffield & Thomas 1997, Presley 2000, Donadio et al. 2001, Maffei et al. 2005). However, we disagree with Novaro (1997) and Salvatori et al. (1999) who considered the Andean Fox to be nocturnal. Most of our photographs of this species were predominantly taken during the day. We placed O. virginianus in the category of cathemeral; however this activity pattern might be local. Previous studies in O. virginianus in North America mentioned that the activity patterns for this cervid depended on environment conditions (Beier & McDullough 1990). Our data for Mountain Paca shows more activity from 20:00 h to 22:00 h (Fig. 4), similar to what was obtained for the Lowland Paca Cuniculus paca (Linnaeus 1766) (Gómez et al. 2005).
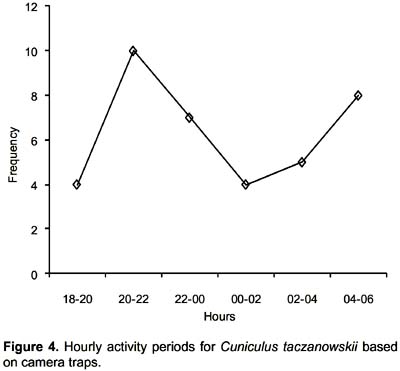
Van Schaik and Griffiths (1996) reported for Indonesian mammals a relation between body size and activity patterns, where small mammals tend to be nocturnal as an anti-predation strategy and medium-size mammals are expected to be cathemeral and diurnal. However, our data suggest that the Mountain Paca (medium mammal) appears to be an exception. Nocturnal activity might be a strategy to avoid predation or to take advantage of underutilized food niches, but not to avoid competition as pointed out by Gómez et al. (2005) for the Lowland Paca Cuniculus paca. Lowland Paca avoids direct competition with the Brown Agouti D. punctata by means of temporal separation of their activity patterns.
The elevational record of the Tayra was increased in 179 meters from the previous one by Alberico et al. (2000). This species is unusual < 1200 m (Eisenberg 1989). The Ocelot's highest observed altitude was at 2900 m in Colombia based on tracks (Sánchez et al. 2004). In this study, the Ocelot was observed at 3379 m, 479 m higher than the previous record. Tewes and Schmidly (1987) indicated the Ocelot is able to occupy different sort of habitats such as humid tropical, subtropical forests, swampy savannas, estuarine mangroves and thorny bushes, but it is most common at elevations lower than 1200 m (Eisenberg 1989). We recorded it in both season for which we think that felid is a permanent resident in the area. Furthermore this carnivore had been recorded previously in other mountain forest of Mexico (Chávez-León 2005) and Colombia (Sánchez et al. 2004)
The Mountain Tapir T. pinchaque inhabits montane forests and Paramos between 2,000 and 4,000 m a.s.l. (Downer 1996, 1997). Lizcano and Sissa (2003) recorded it in the Tabaconas-Namballe National Sanctuary and mentioned that the mountain tapir is absent south of Huancabamba valley. Our record south of the Huancabamba depression by an unequivocal track extends significantly its range and challenge the role of the Huancabamba Depression as a potential barrier for large mammals.
Noteworthy records of mammals found in the study site demonstrates its importance and the need for a better protection status of the Pagaibamba Forest. In spite of the advance in knowledge, more species are expected to be known in the region.
Acknowledgments
We wish to express our gratitude to Rio Tinto Minera Perú and Golder Associates Perú S.A for allowing us to use and publish this data, and for the logistic support to accomplish this work. We also thank our colleagues Richard Cadenillas and Dina Farfán for their support during the field work.
Literature cited
Alberico M., A. Cadena, J. Hernández & Y. Muñoz-Saba. 2000. Mamíferos (Synapsida: Theria) de Colombia. Biota Colombiana 1: 43-75.
Arispe R., C. Venegas & D. Rumiz. 2008. Abundancia y patrones de actividad del mapache (Procyon cancrivorus) en un bosque Chiquitano de Bolivia. Mastozoología Neotropical 15(2): 323-333.
Beier P. & D.R. McDullough. 1990. Factors influencing white-tailed deer activity patterns and habitat use. Wildlife Monographs, 109. 51pp.
Castro J.J. 1993. Agouti taczanowskii. En: Aspectos etológicos y reproductivos del tinajo o guagua negra Agouti taczanowskii (Rodentia: Agoutidae) en cautiverio. Revista Asoc. Col. Ciencias Biológicas 6(1):7-11.
Chávez-León, G. 2005. A recent record of Leopardus pardalis from Michoacan, México. Revista Mexicana de Mastozoología. 9:123-127.
Dillon A. & M.J. Kelly. 2008. Ocelot home range, overlap and density: comparing radio telemetry with camera trapping. Journal of Zoology (London) 275: 391-398.
Donadio E., S. Di Martino, M. Aubone & A.J. Novaro. 2001. Activity patterns, home range, and habitat selection of the common hog-nosed skunk, Conepatus chinga (Mammalia, Mustelidae), in northwestern Patagonia. Mammalia 65(1): 49-54.
Downer C.C. 1996. The mountain tapir, endangered "flagship" species of the high Andes. Oryx 30:45-58.
Downer C.C. 1997. Status and action plan of the mountain tapir (Tapirus pinchaque). In: D. M. Brooks, R. E. Bodmer & S. Matola, (Eds.) Tapirs, Status, Survey and Conservation Action Plan. IUCN, Gland, Switzerland.
Eisenberg J. F. 1989. Mammals of the Neotropics: the northern Neotropics, Panama, Colombia, Venezuela, Guyana, Suriname, French Guiana. The University of Chicago Press, Illinois, 1:1-449.
Gómez H., R.B. Wallace, G. Ayala & R. Tejada. 2005. Dry season activity periods of some Amazonian mammals. Studies Neotropical Fauna Environmental 40: 91-95.
Kelly J.M. & E.L. Holub. (2008). Camera trapping of carnivores:trap success among camera types and across species, and habitat selection by species on Salt Pond Mountain, Giles County, Virginia. Northeastern Naturalist, 15(2), 249-262.
Lizcano D.J. & A. Sissa. 2003. Notes on the Distribution, and Conservation Status of Mountain Tapir (Tapirus pinchaque) in North Peru. Tapir Cons. 12:21-24.
Ludlow M.E. & M.E. Sunquist. 1987. Ecology and behavior of ocelots in Venezuela. National Geographic Research 3: 447-461.
Lyra-Jorge M.C., G. Ciocheti, V.R. Pivello & S.T. Meirelles. 2008. Comparing methods for sampling large- and medium-sized mammals: camera traps and track plots. European Journal of Wildlife Research 54(4): 739-744.
Maffei L., A.J. Noss & C. Fiorello. 2007. The jaguarundi (puma yagouaroundi) In the kaa-iya del gran chaco national park, Santa Cruz, Bolivia. Mastozoología Neotropical 14(2): 263-266.
Maffei L., A.J. Noss, E. Cuellar & D.I. Rumiz. 2005. Ocelot (Felis pardalis) population densities, activity and ranging behaviour in the dry forests of eastern Bolivia: data from camera trapping. Journal Tropical Ecology. 21: 349-353.
Marnewick K., P. Funston & U. Karanth. 2008. Camera trapping as method for estimating cheetah abundance. South African Journal of Wildlife Research 38(1): 59-65.
Novaro A.J. 1997. Pseudalopex culpaeus. Mammalian species 558: 1-8.
Patterson, B.D., G. Ceballos, W. Sechrest, et al. 2007. Digital Distribution Maps of the Mammals of the Western Hemisphere, version 3.0. NatureServe, Arlington, Virginia, USA.
Peyton B. 1980. Ecology, distribution, and food habits of spectacled bears, Tremarctos ornatus, in Peru. Journal of Mammalogy 61: 639-652.
Presley S.J. 2000. Eira barbara. Mammalian species 636: 1-6.
Ríos-Uzeda B., H. Gómez & R.B. Wallace. 2007. A preliminary density estimate for Andean bear using camera-trapping methods. Ursus 18(1):124-128.
Rowcliffe J.M., J. Field, S.T. Turvey & C. Carbone. 2008 Estimating animal density using camera traps without the need for individual recognition. Journal of Applied Ecology 45(4): 1228-1236.
Salvatori V., G. Vaglio-Laurin, P.L. Meserve, et al. 1999. Spatial organization, activity, and social interactions of culpeo foxes (Pseudalopex culpaeus) in north-central Chile. Journal of Mammalogy 80: 980-985.
Sánchez F., P. Sánchez-Palomino & A. Cadena. 2004. Inventario de mamíferos en un bosque de los andes centrales de Colombia. Caldasia 26(1): 291-309.
Sheffield S.R. & H.H. Thomas. 1997. Mustela frenata. Mammalian Species 570:1-9.
Shek C.T., C.S.M. Chan & Y.F. Wan. 2007. Camera trap survey of Hong Kong terrestrial mammals in 2002-06. Hong Kong. Biodiversity 15: 1-11.
Sidell B.P. 2002. Moonrise 3.5 (32Bit) Software.
Silver S.C., L. Ostro, L.K. Marsh, et al. 2004. The use of camera traps for estimating jaguar Panthera onca abundance and density using capture/recapture analysis. Oryx 38(2):148-154.
Stirrat S.C. 2003. Seasonal changes in home-range area and habitat use by the agile wallaby (Macropus agilis). Wildlife Research 30, 593-600.
Tewes M.E., & D.J. Schmidly. 1987. The neotropical felids: jaguar, ocelot, margay, and jaguarundi. Pp. 695-712, en: M. Novak, J.A. Baker, M.E. Obbard, and B. Malloch Eds. Wild furbearer management and conservation in North America. Ministry of Natural Resources, Ontario, Canada.
Tobler M.W., S.E. Carrillo-Percastegui & G. Powell. 2009. Habitat use, activity patterns and use of mineral licks by five species of ungulate in south-eastern Peru. Journal of Tropical Ecology, 25(3): 261-270.
Tobler M.W., S.E. Carrillo-Percastegui, R. Leite Pitman, et al. 2008. An evaluation of camera traps for inventorying large and medium sized terrestrial rainforest mammals. Animal Conservation 11: 169-178.
Trolle M. & M. Kery. 2005. Camera-trap study of ocelot and other secretive mammals in the northern Pantanal. Mammalia 69(3-4): 405-412.
Van Schaik C.P. & M. Griffiths. 1996. Activity periods of Indonesian rain forest mammals. Biotropica 28: 105-112.
Varma S., A. Pittet & H.S. Jamadagni. 2006. Experimenting usage of camera-traps for population dynamics study of the Asian elephant Elephas maximus in southern India. Current Science 91(3): 324-331.
Presentado: 24/11/2009
Aceptado: 11/03/2010
Publicado online: 14/12/2010
