










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. v.18 n.2 Lima ago. 2011
TRABAJOS ORIGINALES
Hunting pressure on cracids (Cracidae: Aves) in forest concessions in Peru
Presión de caza sobre crácidos (Cracidae: Aves) en concesiones forestales en Perú
Javier Barrio
Centro de Ornitología y Biodiversidad – CORBIDI
Santa Rita 105, Of. 2. Urb. Huertos de San Antonio, Lima 33, Perú.
E-mail: javbar@gmail.com, javbar@corbidi.org
Abstract
The impact of timber exploitation on biodiversity is usually increased by hunting in the exploited area. Proper forest management practices on areas under commercial exploitation minimize hunting and damage to the forest. Large species of Cracidae, the most endangered family of birds in the Neotropics, are among the first to be affected in a Neotropical forest damaged by timber-extraction activities, and where at least moderate hunting occurs. Herein an assessment of cracids is carried out in three areas with selective logging in Peru in 2004 and 2005, is used to evaluate hunting pressure. Tree inventory trails were used as transects, and density was calculated using the line transect methodology. Four species of cracids were evaluated, and density was calculated for three of them. The area with lower hunting pressure, Maderyja, showed higher cracid diversity and was the only with the presence of razor-billed curassows (Mitu tuberosum) and blue-throated pining-guans (Pipile cumanensis), two sought-after prey species. Areas where hunting intensity is higher had lower cracid diversity. The density of the M. tuberosum was high in Maderyja: 11.3 ind/km2 (95% CI: 7.4 – 17.3 ind/km2). In contrast, Spixs guan (Penelope jacquacu) did not show a marked difference among areas, unless compared to heavily hunted sites. The higher diversity of cracids and the density found for razor-billed curassows suggests Maderyja had low hunting pressure in the past and is properly managed towards wildlife. Currently, the Peruvian Amazon is being opened for forestry concessions and hydrocarbons exploitation and proper management towards wildlife is necessary to guarantee the conservation of susceptible taxa such as cracids.
Keywords. Cracidae; Distance sampling; Forest management; Hunting; Peru.
Resumen
El impacto de la explotación maderera sobre la biodiversidad se incrementa por la caza en la zona explotada. Prácticas adecuadas de manejo forestal en áreas bajo explotación comercial reducen la caza y los daños al bosque. Las especies de aves de gran tamaño como los Cracidae, la familia más amenazada de aves en el Neotrópico, se encuentran entre las primeras afectadas por la extracción madedera y con la actividad de caza aunque sea moderada. En el presente trabajo, como una forma de evaluar la presión de caza, entre 2004 y 2005, se estudiaron las poblaciones de crácidos en tres zonas donde se realiza tala selectiva en el Perú. Senderos para inventario de árboles fueron utilizados como transectos, y la densidad se calculo utilizando la metodología de transecto lineal. Cuatro especies de crácidos fueron evaluados, y la densidad fue calculada para tres de ellos. El área con menor presión de la caza, Maderyja, mostro una mayor diversidad de crácidos y fue la única con la presencia del Paujil Común (Mitu tuberosum) y la Pava de Garganta Azul (Pipile cumanensis), dos codiciadas presas. La diversidad de crácidos fue menor en áreas con intensidad de la caza mayor. La densidad de M. tuberosum fue mayor en Maderyja: 11,3 ind/km2 (95% CI: 7,4 – 17,3 ind/km2). Por el contrario, la Pava de Spix (Penelope jacquacu) no mostró diferencia significativa entre las áreas, salvo comparada con sitios con intensa actividad de caza. La mayor diversidad de crácidos y la densidad encontrada para M. tuberosum sugiere que Maderyja tuvo baja presión de la caza en el pasado y tiene un adecuado manejo de la biodiversidad. En la actualidad, la Amazonía peruana se abre para las concesiones forestales y la explotación de hidrocarburos y el manejo adecuado a la vida silvestre es necesario para garantizar la conservación de los taxones sensibles como crácidos.
Palabras clave: Cracidae; transecto lineal; manejo forestal; caza; Perú.
Introduction
Differences in abundance of cracids (Galliformes: Aves) in disturbed and undisturbed forest areas has been widely used as indicator of the impacts of deforestation and hunting practices (Brooks 2006, Brooks & Fuller 2006, Silva & Strahl 1991, Strahl & Grajal 1991, Strahl & Silva 1997). Large species of cracids -which have intrinsically low rates of reproduction and are dependent on undisturbed forest-, are usually absent or at very low population densities in forests with medium to high hunting pressure (Begazo & Bodmer 1998, Brooks et al. 2001, Brooks 2006, Cancino & Brooks 2006, ONeill 1997, Silva & Strahl 1991). On the other hand, larger Cracid species seems to thrive in areas with low or null hunting pressure (Torres 1997, Yahuarcani et al. 2009).
Evaluating how cracids respond to landscape changes has important ecological and conservation implications (Meijaard et al. 2005, Santamaría & Franco 2000, Strahl & Grajal 1991, Traveset 1998, Yumoto 1999), particularly if they perform important functions in the ecosystem such as seed dispersal or seed predation (Brooks and Fuller 2006, Levey 1994, Muñoz & Kattan 2007, Santamaría & Franco 2000, Yumoto 1999). High fluctuations in population or partial disappearances of cracids in a certain area would notably affect the ecosystem due to their influence on forest regeneration processes (Levey 1994, Meijaard et al. 2005, Santamaría & Franco 2000, Strahl & Grajal 1991, Traveset 1998, Yumoto 1999).
Despite the fact that some Peruvian forestry concessions are located far from human settlements, the impact on wildlife is high when there is uncontrolled hunting, mostly by people working in the concessions (Schulte & Rossiter 2003, Velásquez 2002). The same can be said about native communities which may cause local extinction of wildlife (Bennett & Robinson 2000, Bodmer et al. 1997, Redford & Robinson 1991), even when only subsistence hunting is involved (Aquino et al. 2001, Peres 2001, Zapata 2001).
In this work, I studied the impacts of perturbations caused by logging and increased wildlife hunting on the abundance and density of cracids. Cracids were evaluated in three areas with selective logging in May – June 2004 and April – September 2005, as part of wildlife rapid assessments of forestry concessions in the Amazonian lowlands of southeastern Peru. All of the evaluated areas have been previously active concessions for at least three years.
Material and methods
Study areas
Five different forestry concessions were analyzed (Fig. 1): three on native community land and two on land leased by the Peruvian government (CFA and Maderija). For the purpose of the paper, the three native communities will be treated as a single unit, as logging and hunting practices are similar in the three areas (AIDER 2004). The rainy season spans November to March, and the dry season from May to September. The study was carried on different months of the year, but all during the dry season so they are considered as comparable. Despite forest structure might differ among areas, it is not regarded as a cause for large differences in cracid species abundances, given the cracid species distribution and the use of forest niches by them (Rodríguez-Mahecha et al. 2005). Furthermore, current selective logging practices are carried similarly in all areas, with few selected individual trees being removed, causing perturbations that span at most 1-ha each time. In all areas the process follows a scheme where the area is divided in 20 lots, selecting individual trees from one lot each year, in a 20-year plan, when it starts over.
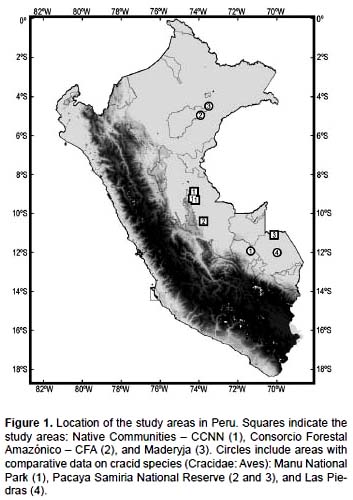
Native Communities along the Ucayali river (CCNN).- These areas are characterized by non-inundated and seasonally-inundated primary forest, with some secondary forest, all under selective logging (AIDER 2004). Three areas are included, centered at 8º5330"S 74º1500"W, 9º1400"S 74º2000"W and 9º2000"S 74º1030"W. The vegetation is heterogeneous in the three areas, with several dominant species including Ficus sp., Virola sp., Manilkara bidentata and Pseudolmedia laevis. The forest canopy is around 35 m high with some emergent trees higher than 40 m. Forest structure was similar throughout the region, mostly covered with lianas, except for some Palm stands. Topography was mostly flat, low terraces with some low hills. Elevation ranges 150-200 m. The areas endure moderate to heavy hunting year round, that is, hunting occurs at least twice a week throughout the three communities, and daily closer to the towns. I additionally assume, based on data from local people, that the forestry concession land at the three native communities has endured continuous hunting throughout the years, leaving an imprint on cracid abundances.
Consorcio Forestal Amazónico (CFA).- This region is characterized by non-inundated primary forest with selective logging. The center of the area was located at 10º24S, 73º46W. The vegetation is heterogeneous and dominated by Ficus sp., Cariniana domesticata, Ocotea sp., Cedrelinga catenaeformis, Terminalia sp. and Virola sp. The undergrowth is semi-open and dominated by the palm Lepidocaryum tessmani. The forest canopy is higher than 40 m and mostly covered with lianas. Topography was mostly nearly flat high terraces with few low terraces to the east. Elevation ranges 280 – 330 m. The area endures moderate hunting throughout the concession, around once a week by the forest inventory and wood extraction crews, which are in the field, but heavy hunting pressure occurs along the northern side, almost daily by local people living close to CFA.
Maderyja.- This region is characterized by non-inundated primary forest with selective logging. The center of the area was located at 11º06S, 70º08W. The vegetation is heterogeneous with several dominant species, including Ficus sp., Swietenia macrophylla and Coumarouna odorata. The undergrowth is semi-open and mostly dominated by Giant bamboo (Guadua sp.). The forest canopy is higher than 40 m and mostly covered with lianas. Topography was dominated by undulated high terraces with some small vertical cliffs, and nearly flat low terraces along the main rivers. Elevation ranges 280 – 350 m. Hunting pressure is low in the region, around twice a month by the forest inventory and wood extraction crews, and centered on medium-size prey such as peccaries.
Species
Four species of cracids were expected to occur in all the study areas. The species are: Speckled chachalaca (Ortalis guttata), Spix's guan (Penelope jacquacu), Blue-throated piping-guan (Pipile cumanensis), and Razor-billed curassow (Mitu tuberosum).
Some characteristics of the species are explained in Table 1.
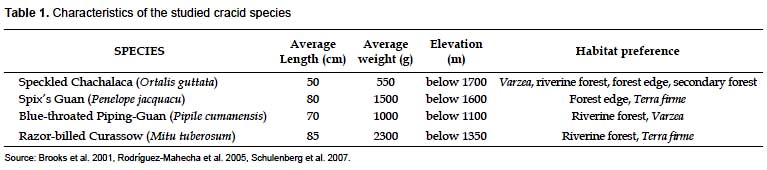
All four species are widespread and, excluding the Razor-billed curassow, are relatively common in western Amazonian rainforest (Rodríguez-Mahecha et al. 2005, Schulenberg et al. 2007). Despite its preferred habitats, the piping-guan can be seen on terra firme occasionally (Schulenberg et al. 2007). As the curassow avoids secondary growth (Rodríguez-Mahecha et al. 2005) and is one of the most commonly hunted species of the Amazonia (Cancino & Brooks 2006), its presence usually denotes areas in good condition and with low or null hunting.
Trails
Selected trails were previously established for forestry inventories and the ones used were separated > 500 m to avoid correlation between them (Buckland et al. 1993, Jiménez et al. 2003). Trails were evaluated approximately for four hours per transect, from 06:00- 10:00 h, and in the afternoon from 15:00 – 19:00 h. Data collected on the trails included species, number of individuals/group, perpendicular distance from the trail to the bird or center of the flock, and coordinates (where possible). Most trails were parallel and routed in a systematic pattern. The native community areas were evaluated during April 2005. Of 14 trails averaging 5.9 km (83 km total), one-half of the trails were evaluated three times each, and the remaining trails were evaluated twice. Total length evaluated was 202 km. at CFA, the evaluation was carried out during May and June 2004. Of 10 five km trails (50 km total), eight trails were evaluated three times, and two were evaluated once. Total length evaluated was 130 km. At Maderyja, the evaluation was carried out during September 2005. Of 13 trails averaging 5.8 km (75 km total) eight trails were evaluated four times each, and five were evaluated three times. Thus, total length evaluated was 271 km.
Density and abundance
The line transect sampling methodology was used to estimate density (Buckland et al. 1993). The program Distance, version 6.0 (Buckland et al. 1993), was used to estimate densities of species with at least 40 sightings. Distances to the observed individuals were measured with a 50 m measuring tape and/or with a rangefinder to use the datum in the Distance software. However, if distribution patterns of observations (number of observations in a distance range) are arranged very similar to pre-selected data distribution models of the Distance software, then fewer sightings may be enough to give a reliable estimate of density, usually corresponding to a coefficient of variability around or less than 30% (Buckland et al. 1993). All models were run by the program to analyze which one fit the data best.
Density is presented as number of individuals/km2. Additionally, an abundance index was used for the four species, and was calculated as the number of individuals observed/10 km walked. Results were compared to densities and encounter rates calculated in other studies, some of them carried out with other methods, and on different seasons.
Comparisons with other studies
The densities and encounter rates found in this study are compared with density data from two Peruvian protected areas: Manu National Park (Torres 1997) and Pacaya Samiria National Reserve (Begazo 1997, Yahuarcani et al. 2009), and with encounter rates at sites near Manu with hunting pressure along the Las Piedras River (Schulte & Rossiter 2003) (Fig. 1). Relative abundance data from Brazils Jaú National Park (Borges et al. 2001) are also compared. Density data for a congener of the razor-billed curassow (Salvin´s curassow Mitu salvini; Johnson 1993) and for the wattled curassow (Crax globulosa, Yahuarcani et al. 2009) are also used here for comparisons.
Results
Four species of cracids, described in methods, were found in the three study areas: speckled chachalaca, Spix's guan, blue-throated piping-guan and razor-billed curassow. The Spixs guan was found in all three areas, the chachalaca was absent from the CFA, and the curassow and piping-guan were only found in Maderyja. The curassow in Maderyja was the only species observed more than 40 times. Based on the distribution pattern of distances and the coefficient of variability of the results for both guans in Maderyja (n= 22 and 23), and the Spixs guan in CFA (n= 21), densities were also calculated for the guans. The three species were commonly found in pairs or small groups, and density of groups (clusters) was also calculated (Table 2).
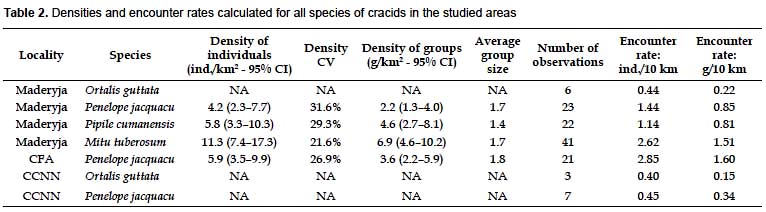
The curassow in Maderyja had by far the highest density of the four species in any area with 11.3 ind/km2 (Table 2). Densities for the piping-guan in Maderyja and the Spixs guan in Maderyja and CFA were more or less similar when considering individuals, ranging 4.2 – 5.9 ind/km2 (see Table 2). The chachalaca was found on few occasions in Maderyja and CCNN, and therefore only the encounter rate was calculated (Table 2).
The encounter rate for the Spixs guan in CFA and the razor-billed curassow in Maderyja were highest with 2.85 and 2.62 ind/10 km, respectively (Table 2). However, encounter rate results are not comparable, given that species do not have the same probability of being detected, and the distribution of observation distances differ among them.
Discusion
Razor-billed curassow
The encounter rate of the curassow in Maderyja (2.6 ind/10 km) is much higher than encounter rates found at Las Piedras (0.0 – 1.2 ind/10 km; Schulte & Rossiter 2003). This species was not found in CCNN along the Ucayali River or in CFA. The contrast in densities between Maderyja and the hunted sites may suggest that curassows could be affected by hunting and population recovery takes several years (Begazo & Bodmer 1998, Estudillo-Lopez 1988, Silva & Strahl 1991, Strahl & Grajal 1991). In Las Piedras, hunting in prior years seems to have influenced the curassow population more than current hunting, as there is no statistical difference between areas with and without timber activity (Schulte & Rossiter 2003).
The density found in Maderyja is comparable to densities found in a year-round study at Manu National Park (15.7 ind/ km2; Torres, 1997), and it is much higher than the reported inside Pacaya-Samiria National Reserve (1.7 ind/km2; Begazo 1997), or at hunted sites surrounding Pacaya-Samiria (0.02 –2.1 ind/km2; Begazo 1997). In Jaú National Park the species is recorded as rare, despite only light hunting in the area (Borges et al. 2001). Density of Salvin´s curassow (Mitu salvini), an ecologically similar congener to M. tuberosum, was calculated as 3.8 ind/km2 in Ecuadorian terra-firme forest with low hunting pressure, and 1.6 ind/km2 in areas with moderate hunting pressure (Johnson 1993). Density of the rare and similar sized wattled curassow (Crax globulosa) in an area of Pacaya Samiria National Reserve was calculated as 3 – 4 ind/km2 and 0.8 – 1.5 groups/km2 (Yahuarcani et al. 2009).
Spixs guan & Blue-throated piping-guan
The encounter rate found for Spixs guan in CFA (2.9 ind/10 km) is twice as high as the rate in Maderyja (1.4 ind/10 km), which in turn is twice as high as the highest density found in any of the CCNN evaluated along the Ucayali river (0.14 – 0.7 ind/10 km). Yet, the encounter rates found in Maderyja and CFA are lower than that of several of the six areas evaluated along Las Piedras river (1.6 – 6.0 ind/10 km; Schulte & Rossiter 2003). The blue-throated piping-guan was not found around CCNN or in the CFA. The encounter rate for blue-throated piping-guans in Maderija (1.1 ind/10 km) is higher than Las Piedras (0.1 – 0.7 ind/10 km; Schulte & Rossiter 2003).
The density of Spixs guan is lower in Maderyja (4.2 ind/ km2) and CFA (5.9 ind/km2) than in Manu (19.8 ind/km2; Torres, 1997), but can be compared to fully protected zones of Pacaya-Samiria (5.5 ind/km2, 95% CI = 3.2 – 9.2; Begazo 1997). The density of the blue-throated piping-guan in Maderyja (5.8 ind/km2) is comparable to Pacaya-Samiria (6.8 ind/km2, 95% CI= 3.3 – 13.8; Begazo 1997), and it is much higher than Manu (1.8 ind/km2; Torres, 1997). Heavily hunted areas outside Pacaya Samiria have lower densities of both species (0.22 and 0.44 for Spixs and piping-guans, respectively; Begazo 1997). In Jaú National Park, the Spix´s guan was only infrequently recorded, while the blue-throated piping-guan was considered rare (Borges et al. 2001).
Densities and encounter rates for both guans are more or less stable in areas with low hunting, where the pressure is distributed over several species, so that guans are only hunted occasionally. However, in areas with continuous hunting such as throughout CCNN, hunting pressure of guans increases because of the lack of other prey which was already over-hunted to local extinction. Differences in proportions of preferred habitats and competition may explain asymmetric encounter rate values between the two species among areas studied or used for comparisons. For example, Spixs guans prefer terra firme forest whereas piping-guans prefer varzea (seasonally inundated forest) and riverine forests (Brooks et al. 2001). Interestingly, the area of varzea and riverine forests in Maderija is small compared to the concessions on CCNN or CFA; nevertheless, Maderija was the only study area where piping-guans were found. The density of piping-guans found there indicates a healthy population along its riverine forests.
Hunting and logging throughout the area
Uncontrolled or excessive hunting may cause a larger impact on the function of the forest than selective logging (Bennett & Robinson 2000, Meijaard et al. 2005). Trails and road networks inside forestry concessions facilitate hunting and can cause an increase in hunting pressure. The main cause of differences in large cracid densities is human perturbations (Begazo & Bodmer 1998, Brooks et al. 2001, Cancino & Brooks 2006, ONeill 1997).
In other areas of the Amazon curassows are the most preferred game birds as they are large and easy to find in areas where the population is stable (e.g. Brooks et al. 2001). Hunting was still low and under supervision based on controls by the timber company at the main road at Maderyja.
The high abundance of razor-billed curassows and their unwary behavior might corroborate that the hunting level in Maderyja was very low and forest structure was adequate. Other studied areas in CCNN were heavily hunted in the past, and thus, lacked razor-billed curassows. Unperturbed primary Amazonian forests with little or no hunting are correlated with the abundance of large cracids (Aranibar-Rojas & Hennessey 2006, Begazo 1997, Brooks 1999, ONeill 1997, Silva & Strahl 1991). However, in the case of guans, extension of preferred habitats and competition between species of more or less similar size and habits might be taken into account when comparing sites (Remsen & Cardiff 1990). If there is a decrease in the number of Cracid density or if their behavior becomes more wary we can assume an increase in hunting pressure has occurred. As current numbers indicate it might be possible to maintain an area under selective logging while Cracid populations remain stable. If the damage to the forest is low under selective logging, and hunting is directed towards less susceptible wildlife species, then the impact on cracids should be low in turn.
Conservation and Development
The populations of Cracid species are primarily affected by hunting and habitat destruction (Brooks & Fuller 2006, Meijaard et al. 2005, Rodríguez-Mahecha et al. 2005). Some cracid species use secondary forests, but large areas of primary forest free of hunting are necessary for their conservation (Begazo & Bodmer 1998, Borges 1999). At present, when most of the Peruvian Amazon is being deforested due to forestry concessions and/ or oil or gas exploration and exploitation; proper management towards wildlife is necessary to guarantee the conservation of the taxa most susceptible to be affected, such as cracids.
Despite the four species of cracids treated here are considered not to be threatened by the IUCN red list, in all cases it is stated that the population trend appears to be decreasing (IUCN 2010). Moreover, some of the species are not only decreasing but have disappeared from large tracts of the Amazonian rainforest or occur at very low densities. A comprehensive assessment on the razor-billed curassow and the blue-throated piping-guan, species affected by unsustainable hunting (Begazo & Bodmer 1998), might show a declining trend sufficient as to be listed as endangered species.
Acknowledgements
The data used in this study were obtained while conducting wildlife evaluations for the World Wildlife Fund-Peru (WWF), under the project CEDEFOR. I am grateful to WWF for the opportunity to publish some of the results. D. Brooks was invaluable reviewing the manuscript. I thank F. Angulo and R. McNeil for reviewing an early manuscript. I also thank all the people that helped during the evaluations.
Literature cited
Aquino R., R. Bodmer & J. Gil. 2001.
Mamíferos de la cuenca del río Samiria: ecología poblacional y sustentabilidad de la caza. Junglevagt for Amazonas, AIF-WWF/DK & Wildlife Conservation Society, Washington, D.C.Aranibar-Rojas H. & A. B. Hennessey.
2006. Historia natural y estimación preliminar de la abundancia relativa mediante tasas de encuentro de Crax globulosa en un bosque de várzea en el departamento de Beni, Bolivia. Cotinga 26: 32-35.Asociación para la Investigación y el Desarrollo Integral – AIDER. 2004. Planes generales de manejo forestal elaborados por AIDER para bosques comunales de la Amazonía peruana. Volúmenes 1 y 2. AIDER y Embajada Real de los Países Bajos.
Pucallpa, Perú.Begazo A. 1997. Use and conservation of the Cracidae in the Peruvian Amazon. In: S. Strahl, S. Beujon, D. Brooks, A. Begazo, G. Sedaghatkish and F. Olmos, eds. The Cracidae: Their biology and conservation. Washington, D.C.: Hancock House. Pp 449-459.
Begazo A. & R. Bodmer. 1998. Use and conservation of Cracidae (Aves: Galliformes) in the Peruvian Amazon. Oryx 32: 301–309.
Bennett E. & J. Robinson. 2000. Hunting of wildlife in tropical forests. Implications for biodiversity and forest peoples. Paper 76, Biodiversity Series: Impact Studies, The World Bank Environment Department, Washington, D.C.
Bodmer R., J. Eisenberg & K. Redford. 1997. Hunting and the likelihood of extinction of Amazonian mammals. Conservation Biology 11: 460–466.
Borges S. H. 1999. Relative use of secondary forests by cracids in central Amazonia. Ornitología Neotropical 10: 77-80.
Borges S. H., M. Cohn-Haft, A. M. Pereira Carvalhaes, L. M. Henriques, J. F. Pacheco & A. Whittaker. 2001. Birds of Jaú National Park, Brazilian Amazon: species check-list, biogeography and conservation. Ornitología Neotropical 12: 109–140.
Brooks D.M. 2006. The utility of hotspot identification for forest management: Cracids as bioindicators. Acta Zoologica Sinica 52(Supplement): 199-201.
Brooks D. M. 1999. Pipile as a protein source to rural hunters and Amerindians. In D. A. Brooks, A. Begazo and F. Olmos, eds. Biology and conservation of the Piping Guans (Pipile). Special Publication Cracid Specialist Group, 1. Houston, Texas. Pp 42-50.
Brooks D. M., and R. A. Fuller. 2006. Biology and Conservation of Cracids. In L. Cancino and D. M. Brooks, eds. Conserving Cracids: The most threatened family of birds in the Americas. Misc. Publ., Nº 6, Houston Museum of Natural Science, Houston, Texas.
Pp 11-26.Brooks D. M., L. Pando, A. Ocmin & J. Tejada.
2001. Resource separation in a Napo-Amazonian gamebird community. In D. M. Brooks and F. Gonzalez, eds. Cracid Ecology and Conservation in the New Millenium. Misc. Publ. Nº 2, Houston Museum of Natural Science, Houston, Texas. Pp 213-225.Buckland S. T., D. R. Anderson, K. P. Burnham & J. L. Laake. 1993. Distance sampling: Estimating abundance of biological populations. London, UK: Chapman and Hall.
Cancino L. & D. M. Brooks (eds.). Conserving Cracids: The most threatened family of birds in the Americas. Misc. Publ., Nº 6, Houston Museum of Natural Science, Houston, Texas.
Estudillo-Lopez, J. 1988. Breeding cracids in captivity. Proceedings of World Conference of Breeding of Endangered Species in Captivity, 5: 159-163.
International Union for Conservation of Nature and Natural Resources (IUCN) (on line). 2010. IUCN Red List of Threatened Species. Version 2010.4 <www.iucnredlist.org> Downloaded on 25 October 2010.
Jiménez I., G.A. Londoño & C. D. Cadena.
2003. Efficiency, bias, and consistency of visual and aural censuses for estimating the abundance of Curassows (Cracidae) in tropical forests. Journal of Field Ornithology 74: 210-216.Johnson A. 1993. A case study of the effects of land use zoning by the Cofaun Indians on the conservation of threatened cracids (Cracidae) in the Cuyabeno Reserve, Ecuador. M.Sc. thesis, University of Wisconsin.
Levey D. 1994. Cracids as seed dispersers: What we need to know and why it is important. The Cracid Newsletter 3: 3-4.
Meijaard E., D. Sheil, R. Nasi, D. Augeri, B. Rosenbaum, D. Iskandar, T. Setyawati, M. Lammertink, I. Rachmatika, A. Wong, T. Soehartono, S. Stanley & T. OBrian. 2005. Life after logging: Reconciling wildlife conservation and production forestry in Indonesian Borneo. Bogor, Indonesia: CIFOR.
Muñoz M. & G. H. Kattan. 2007. Diets of cracids: How much do we know?
Ornitología Neotropical 18: 21-36ONeill J. 1997. Los Crácidos: una sobrevista. In S. Strahl, S. Beujon, D. Brooks, A. Begazo, G. Sedaghatkish and F. Olmos, eds.
The Cracidae: Their biology and conservation. Washington, D.C.: Hancock House. Pp 398-400.Peres C. 2001. Synergistic effects of subsistence hunting and habitat fragmentation on Amazonian forest vertebrates.
Conservation Biology 15: 1490–1505.Redford K. & J. Robinson. 1991. Subsistence and commercial uses of wildlife in Latin America. In J. Robinson and K. Redford, eds. Neotropical wildlife use and conservation.
Chicago, Illinois: Univ. of Chicago Press. Pp 6-23.Remsen J. V. Jr. & S. W. Cardiff. 1990. Patterns of elevational and latitudinal distribution, including a "niche switch," in some guans (Cracidae) of the Andes.
Condor 92: 970-981.Rodríguez-Mahecha J.V., N. Hughes, O. Nieto & A.M. Franco. 2005. Paujiles, Pavones, Pavas & Guacharacas Neotropicales. Conservación Internacional, serie libretas de campo. Bogotá, Colombia.
Santamaría M. & A. M. Franco.
2000. Frugivory of Salvin's Curassow in a rainforest of the Colombian Amazon. Wilson Bulletin 112: 473-481.Schulenberg T.S., D.F. Stotz, D.F. Lane, J.P. ONeill & T.A. Parker III. 2007. Birds of Peru. Princeton Field Guides. New Jersey: Princeton University Press.
Schulte-Herbrüggen B. & H. Rossiter. 2003. Project Las Piedras. A socio-ecological investigation into the impact of illegal logging activity in Las Piedras, Madre de Dios, Peru. University of Edinburgh, Edinburgh, U.K.
Silva J. L. & S. Strahl. 1991. Human impact on population of chachalacas, guans and Curassows (Galliformes: Cracidae) in Venezuela. In J. Robinson and K. Redford, eds. Neotropical wildlife use and conservation. Chicago, Illinois: Univ. of Chicago Press. Pp 37–52.
Strahl S. & A. Grajal. 1991. Conservation of large avian frugivores and the management of Neotropical protected areas. Oryx 25: 50-55.
Strahl S. & J. L. Silva. 1997. The Status of the Family Cracidae in Venezuela. In S. Strahl, S. Beujon, D. Brooks, A. Begazo, G. Sedaghatkish and F. Olmos, eds. The Cracidae: Their biology and conservation. Washington, D.C.: Hancock House.
Pp 383-395.Torres B. 1997. Densidades poblacionales de la comunidad de Crácidos en el Parque Nacional del Manú (Perú).
In S. Strahl, S. Beujon, D. Brooks, A. Begazo, G. Sedaghatkish and F. Olmos, eds. The Cracidae: Their biology and conservation. Washington, D.C.: Hancock House. Pp 376-379.Traveset A. 1998. Effect of seed passage through vertebrate frugivores guts on germination: A review.
Perspectives in Plant Ecology 1: 151–190.Velásquez V. 2002. La actividad forestal y su impacto negativo a la fauna silvestre en Madre de Dios. Boletín de Lima 25: 86–98.
Yahuarcani A., K. Morote, A. Calle & M. Chujandama. 2009. Estado de conservación de Crax globulosa en la Reserva Nacional Pacaya Samiria, Loreto. Revista Peruana de Biología 15: 41–49.
Yumoto T. 1999. Seed dispersal by Salvins Curassow, Mitu salvini (Cracidae), in a tropical forest of Colombia: Direct measurement of dispersal distance. Biotropica 31: 654–660.
Zapata G. 2001. Sustentabilidad de la caza de subsistencia: el caso de cuatro comunidades Quichuas en la Amazonía nororiental ecuatoriana. Mastozoología Neotropical 8: 59–66.
Presentado: 25/01/2011
Aceptado: 27/07/2011
Publicado online: 25/08/2011
