










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.19 no.1 Lima Apr. 2012
TRABAJOS ORIGINALES
Anatomy and physicochemical properties of the chambira fiber
Anatomía y propiedades físico-químicas de la fibra de chambira
Manuel Marín1, Betty Millán1 y Francis Kahn2
1 Museo de Historia Natural–Universidad Nacional Mayor de San Marcos, Av. Arenales 1256, Jesús María, Lima, Perú.
2 IRD, UMR DIADE, Casilla 18-1209, Lima, Perú.
Abstract
The anatomy of the unopened spear leaf of the chambira palm (Astrocaryum chambira), from which chambira fiber is extracted, is described and compared with that of three species (Astrocaryum jauari, occasionally used for fiber extraction, A. standleyanum and A. perangustatum, which do not provide any fiber). Chambira fiber consists of non-vascular fiber strands closely adhered to the adaxial hypodermis; this forms a compact unit, which makes it easy to separate from the rest of the lamina. A similar arrangement is found in Astrocaryum jauari; however, the non-vascular fiber cells are shorter and fewer per strand. Differences in leaf anatomy of Astrocaryum standleyanum and A. perangustatum may explain why fiber is not extracted: Astrocaryum standleyanum has a dense row of non-vascular fibrous strands under the adaxial hypodermis similar to Astrocaryum chambira but the large size of non-vascular fibrous strands that are located in the abaxial part of the mesophyll makes it difficult to properly extract fiber as is done with chambira. In Astrocaryum perangustatum, non-vascular fiber strands are small and irregularly dispersed in the mesophyll, which makes it impossible to extract quality fibers. Density, thickness and tensile strength are higher in chambira fiber than in jauari fiber. Chambira fiber presents a very high cellulose content (93.9%), while lignin content is low (4.2%). The ultrastructure of the cell wall of the non-vascular fiber is described for Astrocaryum chambira only. The major trait is a three-layered cell wall, with a notably thick inner layer. The physicochemical properties of chambira fiber are compared with other vegetal fibers used by the industry.
Keywords: Arecaceae, Astrocaryum chambira, vegetal fiber, promising species, Peru.
Resumen
Se describe la anatomía de la lamina de la pinna en hojas no abiertas de la palmera chambira (Astrocaryum chambira), la que se utiliza para extraer la fibra, comparándola con otras tres especies (Astrocaryum jauari, de la cual ocasionalmente se obtiene una fibra, A. standleyanum y A. perangustatum, estas no proveen fibras). La estructura de la fibra consiste en paquetes de fibras no vasculares adheridos a la hipodermis y epidermis adaxial, lo cuales forman una unidad compacta, que se hace fácil separar del resto de la lámina. Una disposición similar se encuentra en la fibra de Astrocaryum jauari; sin embargo, esta especie tiene menos fibras celulares no vasculares por paquete, con menor longitud. Una disposición diferente es hallada en las otras dos especies lo cual no permite una separación de las fibras: en Astrocaryum standleyanum los paquetes de fibras no vasculares son localizados debajo de la hipodermis adaxial como lo son en Astrocaryum chambira, pero los paquetes de fibras no vasculares localizados en la parte abaxial del mesófilo son significativamente más grandes, lo que hace dificil extraer la fibra como se practica en chambira. En Astrocaryum perangustatum, los paquetes de fibras no vasculares son de menor tamaño y están dispersos en el mesófilo, lo que hace imposible la extracción de una fibra de calidad. La densidad, espesor y resistencia a la tracción son mayores en las fibras de Astrocaryum chambira que en las fibras de A. jauari. La fibra de chambira presenta un alto contenido de celulosa (93,9%) y bajo contenido de lignina (4,2%). La ultraestructura de la pared celular de la fibra no vascular es descrita sólo para A. chambira, la cual presenta una pared celular estratificada en tres capas, la interna es notablemente gruesa. Las propiedades físico-químicas de la fibra de chambira son comparadas con las fibras vegetales utilizadas en la industria.
Palabras claves: Arecaceae, Astrocaryum chambira, fibras vegetales, especies promisorias, Perú.
Introduction
Palm fibers are used to manufacture a variety of goods and may represent a locally non-negligible economic resource (Schultes 1977). Several parts of the plant provide fiber: hard fiber for manufacturing brushes from the leaf sheath of some species such as Aphandra natalia (Balslev et al. 2008, Kronborg et al. 2008) or Leopoldinia piassaba (Lescure et al. 1992); rot-resistant fiber, e.g. for stuffing car seats, from the fruit of Cocos nucifera (Balick & Beck 1991); raw material for manufacturing furniture from the stem of the Calamus species (Kalima & Jasni 2004). Cellulose fibers extracted from the leaf of Chamaerops humilis (Benahmed-bouhafsoun et al. 2007) or petiole of Mauritia flexuosa (Bresani 1924, De Los Heros & Zárate 1980) are considered promising material by the cellulose industry.
Astrocaryum chambira, the chambira palm, provides a soft fiber commonly used to make carrying bags and hammocks. Chambira fiber is extracted from the yellowish pinnae of the young spear leaf that emerges from the crown center. According to Holm Jensen & Baslev (1995), who provided a complete description of processing chambira fiber and products for several Ecuadorean ethnic groups, fiber extracted from mature leaves was used to make fishing nets. In the Peruvian Amazon, chambira fiber is locally extracted and manufactured goods are sold in local and regional markets, and in the many souvenir shops found in the main touristic cities (Mejía, 1988; Vormisto, 2002). Other species of the genus Astrocaryum (A. standleyanum, A. aculeatum, A. jauari, and A. vulgare) are occasionally used for fiber extraction from the lamina (Borgtoft Pedersen 1994, Holm Jensen & Baslev 1995, Kahn 2008).
Leaf anatomy of several species of Astrocaryum, including those mentioned in this article, was formerly described from seedlings (Millán & Kahn 2010). Here, anatomical study focuses on chambira fiber and compares it with two species, Astrocaryum jauari and A. standleyanum (the former is occasionally used for fiber extraction, the latter does not provide any individual fiber), which both belong to the subgenus Astrocaryum, and with a third species, Astrocaryum perangustatum, which belongs to the subgenus Monogynanthus and the leaves of which are never used for fiber extraction. Physicochemical and ultrastructural characterization of chambira fiber cell wall completes the anatomical study. The fibers are compared with vegetal fibers used by industry.
Material and methods
Palm species (Fig. 1) Astrocaryum chambira is a large solitary palm; it grows in primary forest as well as in secondary vegetation and open areas in the western Amazonian lowlands and slopes of the eastern Andean foothills in Ecuador, in western Brazil, and extends to the north and central eastern regions of Peru. Astrocaryum jauari is a large riparian caespitose palm that produces several stems from its base; it is commonly found along most rivers of the Amazon basin. Astrocaryum standleyanum is a medium-sized solitary palm, it grows most commonly in lowland rainforests on poorly drained soils along the Pacific slope from Costa Rica to the Pacific lowlands of western Colombia and Ecuador. These three species develop ragged leaves, i.e. the pinnae are irregularly grouped and oriented in several directions from the rachis. Astrocaryum perangustatum is a large-leaved, short-stemmed palm, the pinnae are regularly disposed in one plane; it is endemic to central Sub-Andean valleys (Millán, 2006), where it grows in forest understory as well as in open areas.

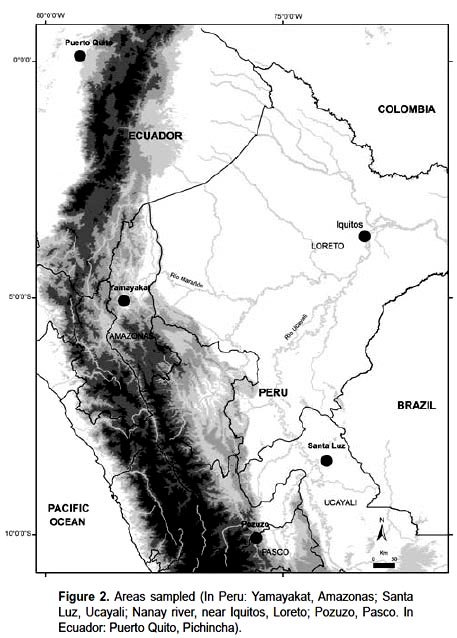
Material collected – Medial pinnae of unopened spear leaf (those commonly used for fiber extraction) were collected in 2-5 individuals from the following localities: Astrocaryum chambira in Yamayakat (78º2011"W, 05º0319"S), Amazonas, northern Peru; Santa Luz (74°0340"W, 08°2605"S), Abujao River, Ucayali, central eastern Peru. Astrocaryum jauari Nanay River valley (73º1528, 3º4146S) Maynas, Loreto, Peru. Astrocaryum standleyanum – in Puerto Quito (00º0700"N, 79º1600"W), Pichincha, Northern Ecuador. Astrocaryum perangustatum in the Pozuzo region (75º3316"W, 10º0405"S), Pasco, Peru (Fig. 2).
Anatomical study Fresh portions of 5 cm length taken from the midpart of medial pinnae of unopened spear leaves of the four species were fixed in FAA (formalin, alcohol, glacial acetic acid). Cross sections were cut by hand, bleached in commercial sodium hypochlorite and 5% chloral hydrate, stained in 1% safranin; zinc chloride-iodide and phloroglucin microchemical tests were applied for examination of cellulose and lignified fiber walls respectively (D'Ambrogio 1996). Fibers were extracted from medial pinnae from unopened spear leaves of Astrocaryum chambira and A. jauari following the traditional Amazonian process (Fig. 1); fiber portions of 1 cm length were macerated in 5% sodium hydroxide solution, boiled for 30 minutes, bleached with 5% chloral hydrate, and stained in 1% safranin (Msahli et al. 2007 modified).
Observations and photographs were made at 100x and 400x magnification with a Leica ICC50 photomicroscope; digital photographs were processed with Adobe Photoshop CS4.
Biometrics Fibers and fiber cells were measured using an ocular micrometer. Data were obtained for three laminas per species. Mean values were calculated from 30 measurements (10 per lamina) for thickness of lamina, thickness of the fiber extracted from the leaf, length of adaxial and abaxial fibrous strands, number of fiber cells per non-vascular fibrous strand, and 90 measurements (30 per lamina) for length and width of non-vascular fiber cells. Significant differences at p<0.05 and p<0.01 were determined using ANOVA and pairwise comparison of means (Fishers LSD).
Physicochemical analysis of non-vascular fibers Fiber thickness and density (adapted from ASTM D 143) were measured with a Mitutoyo digital caliper, model CD-62BS (adapted from ASTM D 2256 and ASTM D 143, 1993). The tensile test was carried out for six units of Astrocaryum chambira fiber and one unit of Astrocaryum jauari fiber using an Alfred J. Amsler and Co. universal tensile machine, Model 46/224 (ASTM D 2256).
Cellulose and lignin contents were quantified for the chambira fiber; dry fibers were previously macerated in a mixture of 1:2 ethanol-benzene for release of extractives, and processed according to TAPPI methods T 203 I-74 for cellulose and T 222 I-74 for lignin (TAPPI 1978a, b).
Ultrastructure of chambira fiber cell wall Fiber palm samples were fixed in 2.5% glutaraldehyde in 0.1 M phosphate buffer, pH 7.2, washed in 0.1 M phosphate buffer, pH 7.2 for about 8 hours and postfixed in 1% osmium tetroxide in 0.1 M phosphate buffer pH 7.2 for 12 hours. The samples were washed in 0.1 M phosphate buffer, pH 7.2 and dehydration took place in a graded ethanol and acetone series. Infiltration and embedding was performed in successive steps in a mixture of acetone and resin Durcopan ® in a 60°C oven for 48 h. Ultra-thin 90 nm thick cross sections were cut with a Porter Blum MT2 ultramicrotome, the sections were stained with uranyl acetate and lead citrate for observation under transmission electron microscopy. (Ancheta et al. 1996).
Results
Unopened spear leaves of the four species share common anatomical characteristics. These include: non-vascular fibrous strands located under the adaxial hypodermis and in the spongy parenchyma between the major vascular bundles, variable in shape and in number of fibers; round stegmata surrounding fiber strands; cellulose non-vascular fiber very elongated, acute at the ends, with thick walls; lignified vascular fibers short, septate, spindle-shaped and acute, located at the ends of the major and minor vascular bundles.
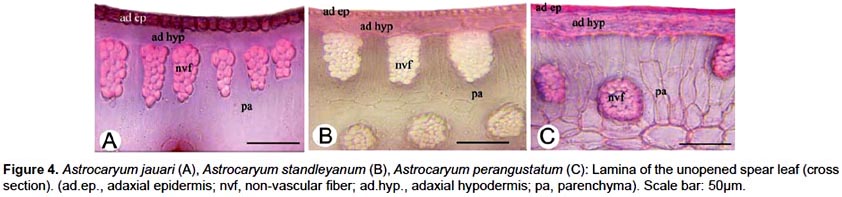
One of the main anatomical differences is that non-vascular fibrous strands form a dense row below the adaxial hypodermis in Astrocaryum chambira, A. jauari and A. standleyanum, while they are irregularly scattered in the mesophyll in Astrocaryum perangustatum (Fig. 3, 4).
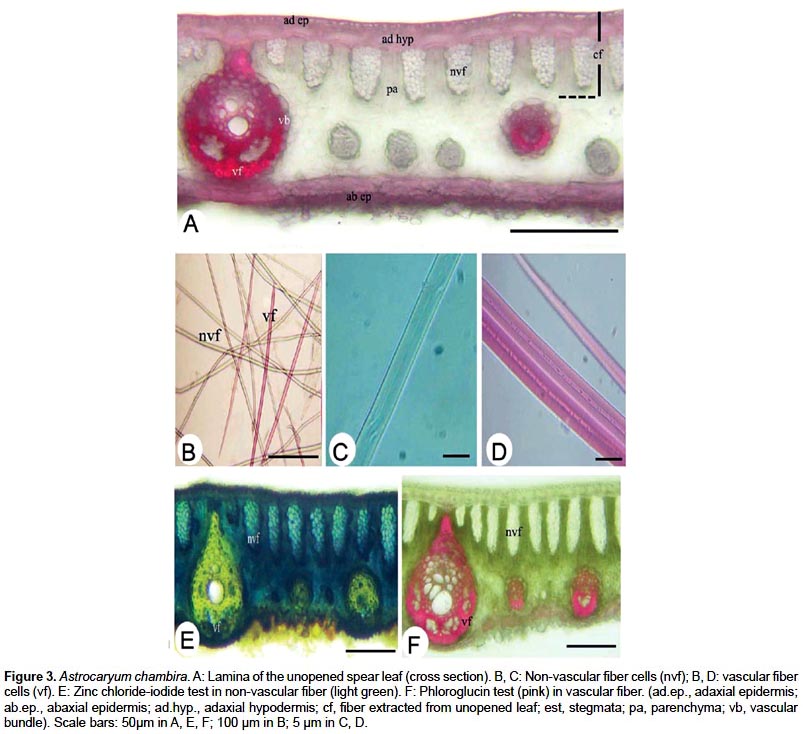
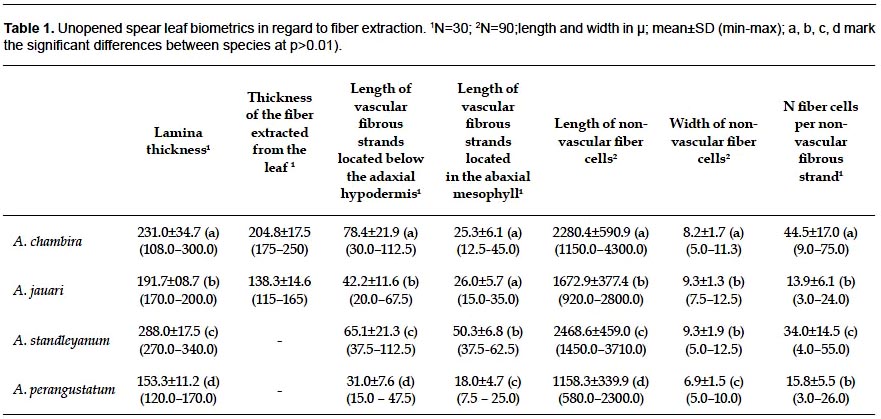
ANOVA applied to biometric parameters obtained from the four species supports significant differences at p<0.01; pairwise comparisons of the means specify those differences as follows (Table 1): (i) Lamina of unopened spear leaves is thicker in Astrocaryum standleyanum, A. chambira and A. chambira than in A. jauari and A. perangustatum. (ii) The fiber extracted from the leaf is thicker in Astrocaryum chambira than in A. jauari (fiber are not extracted from the other two species). (iii) Non-vascular fibrous strands located below the adaxial hypodermis are significantly longer in Astrocaryum chambira and A. standleyanum than in A. jauari and A. perangustatum. Those located in the abaxial mesophyll are shorter than the former, the longest are found in Astrocaryum standleyanum. (iv) Non-vascular fiber cells are longer in Astrocaryum standleyanum and A. chambira than in A. jauari, short in A. perangustatum; they are wider in Astrocaryum jauari and A. standleyanum than in A. chambira and A. perangustatum. (v) The number of fiber cells per non-vascular fibrous strand is highest in Astrocaryum chambira.
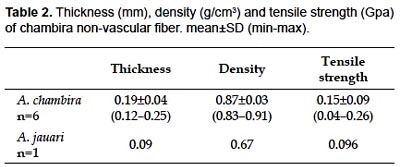
Thickness and density of non-vascular fibers and tensile strength are higher in Astrocaryum chambira than in A. jauari (Table 2). Only one unit of jauari fiber was analyzed, however. Zinc chloride-iodide microchemical testing shows that non-vascular fiber cell walls are mainly composed of cellulose (Fig. 3E). Chemical analysis for commercial fiber extracted from unopened spear leaves gives a high cellulose content of 93.9% in dry matter, while the lignin content is low at 4.2%. Phloroglucin microchemical testing shows that lignin is concentrated in lignified fibers located in fibrous vascular strands (Fig. 3F).
In Astrocaryum chambira, the ultrastructure of non-vascular fiber wall shows a thick, three-layered cell wall (1.75 µ ) and a narrow cytoplasmic lumen (0.91 µ diameter). The primary wall, made of the outer layer (S1), is 0.02 µ thick; the secondary wall is made of both medial (S2) and inner (S3) layers, 0.26 µ and 1.47 µ thick, respectively. Darkness decreases centripetally from S1 to S3. Vascular fiber wall is thin (0.8 µ ) and unstratified with wider lumen than in the non-vascular fiber.
Discussion
Chambira fiber extracted from unopened spear leaf consists of non-vascular fibers closely adhered to adaxial epidermis and hypodermis. A mechanical extraction process removes the unusable part of the mesophyll (spongy parenchyma, non-vascular and vascular fibers).
Anatomical features make clear why leaves of Astrocaryum jauari, A. standleyanum and A. perangustatum are not used regularly or at all for fiber extraction. Non-vascular fiber cells and strands are shorter, and their number per strand is lower in Astrocaryum jauari than in A. chambira. That may explain why the spear leaf of this species is not commonly used for fiber extraction. Moreover, Astrocaryum jauari is a riparian species flooded for several weeks each year; it is less accessible than Astrocaryum chambira that is commonly found in non-flooded areas and cultivated places. Astrocaryum standleyanum has a dense row of non-vascular fibrous strands under the adaxial hypodermis similar to A. chambira but the large size of non-vascular fibrous strands that are located in the abaxial part of the mesophyll makes it difficult to properly extract fibers as is done for chambira fiber. However, this species is used in making handicrafts such as hats and baskets, using the entire pinnae from unopened spear leaves with an appropriate treatment described in Borgtoft Pedersen (1994). Non-vascular fiber strands are small and irregularly dispersed in the mesophyll in Astrocaryum perangustatum, which makes it impossible to extract quality fibers.
Chambira fiber is the largest among the known fibers from palm leaves; moreover, its length is comparable to some fibers of palm stems (e.g. Calamus, Table 3). Fiber cell wall is thicker than in Cocos nucifera fiber (0.089 µ ) and thinner than Linum usitassisimum fiber (5–15 µ ) according to Morvan et al. (2003) and Khalil et al. (2006). Length and width of chambira fiber can be compared to those of several commercial fibers (Table 4); however, only extreme values for these latter are available in the literature cited, which limits the meaning of such a comparison.
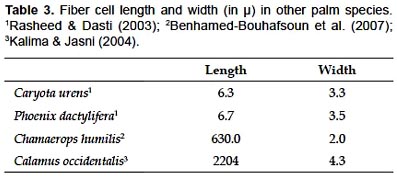
Except for New Zealand linen, Phormium tenax, tensile strength of chambira fiber is weaker than the values reported for other commercial fibers (Table 4), which would be at least partially due to the lack of lignin in the cell wall. Tensile strength is a function of several factors such as the rigidity of the lignified wall (McDougall et al. 1993), the strength of covalent bonds in the pyranose rings and between glucose units of the cellulose polymer chain (Kalima & Jasni 2004), or the orientation of cellulose microfibrils along the axis of the fiber (Sanchis Gritsch & Murphy 2005).
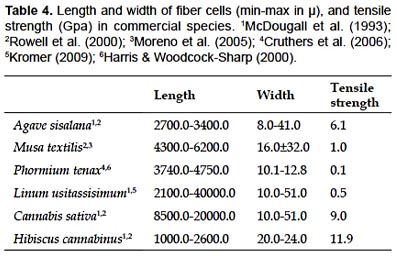
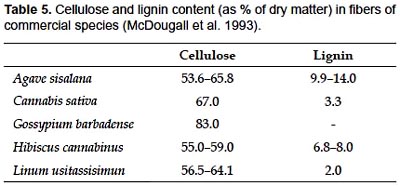
Chambira fiber is essentially composed of cellulose non-vascular fibers, which makes them flexible enough to be valuable for manufacturing bags and hammocks. Cellulose content is higher than those reported for other commercial fibers (Table 5). In this way, chambira fiber differs from most Monocotyledon fibers, which are composed of vascular tissue with lignified cell walls (Tomlinson 1990, McDougall et al. 1993, Evert 2006).
According to Holm Jensen & Baslev (1995), the exploitation of the chambira palm for fiber extraction is a classic example of extractivism in Ecuador; it is used as a shade tree for crops and could be an important component in agro-forestry systems or for revaluing deforested areas. This is also true of Peru. Introducing chambira fiber in industrial fiber process would be a good option to intensify the utilization of this palm and increase local income. However, further research on the ecology and genetic diversity of Astrocaryum chambira are really needed in order to optimize this resource within a sustainable development framework.
Acknowledgements
This work was carried out under the agreement between UNMSM/San Marcos National University, Peru and IRD/Research Institute for Development (UMR DIADE/DYNADIV), France, and supported by the PALMS project funded by the European Community, 7th Framework Programme, Grant Agreement N°212631, and the PROCYT-CONCYTEC-OAJ Project (230-2008). We thank Delya Yabar for her helpul assistance on the English manuscript.
Literature cited
Ancheta, O., M. Ramos, M.C De la Rosa & S. Rodríguez. 1996. Metodología para el procesamiento de tejido vegetal en microscopía electrónica de trasmisión. Biotecnología 13: 195–196.
ASTM D 143. 1993. Standard test method for tensile properties of single fibres. Annual Book of ASTM Standards, Engllwood VS & Brackell.
ASTM D 2256. 1993. Standard test method for tensile properties of single fibres. Annual Book of ASTM Standards, Engllwood VS & Brackell.
Balick, M.J. & H.T. Beck. 1991. Useful palms of the world. A synoptic bibliography. Columbia University Press, New York.
Balslev H., C. Grandez, N. Paniagua, A.L. Møller & S. Lykke. 2008. Useful palms (Arecaceae) near Iquitos, Peruvian Amazon. Rev. peru. biol. 15 (supl. 1): 121–132.
Benahmed-bouhafsoun A., F. Cherifi, M. Haili, Z. Bekhlifi, N. Maamar &M. Kaid-Harche. 2007. Anatomy, histochimy and biometrics of fibres of chamaerops humilis l. Grown in two different locality in algeria. Asian Journal of Plant Sciences 6 (2): 252–260.
Borgtoft Pedersen, H. 1994. Mocora Palm-Fiber: use and management of Astrocaryum standleyanum (Arecaceae) in Ecuador. Economic Botany 48 (3): 310-325.
Bresani, E. 1924. La palmera "aguaje" como productora de celulosa. Boletín de la Sociedad Geográfica de Lima. Tomo XLI: 209–210.
Cruthers N., D. Carr, R. Laing & B. Niven. 2006. Structural differences among fibers from six cultivars of Harakeke (Phormium tenax, New Zealand flax). Textile Research Journal 76 (8): 601–606.
DAmbrogio de Argüeso A. 1996. Manual de técnicas en histología vegetal. Editorial Hemisferio Sur. Buenos Aires.
De Los Heros, G.M.J. & J.B. Zárate. 1980/1981. Posibilidades papeleras de pulpa al sulfato de pecíolos de aguaje. Revista Forestal del Perú 10: 83–90.
Evert, R. 2006. Esau, Anatomía Vegetal. Ediciones Omega. Barcelona.
Harris W. & M. Woodcock-Sharp. 2000. Extraction, content, strength, and extension of Phormium variety fibres prepared for traditional Maori weaving. New Zealand Journal of Botany 38: 469–487.
Holm Jensen, O. & H. Balslev. 1995. Ehtnobotany of the fiber palm Astrocaryum chambira (Arecaceae) in the Amazonian Ecuador. Economic Botany 49 (3): 309–319.
Kahn F. 2008. The genus Astrocaryum. Rev. peru. biol. 15 (supl. 1): 31–48.
Kalima T. & Jasni. 2004. Study of Calamus occidentalis J.R. Witono & J. Dransf. Species Commercial Values and Possible Utilization. Biodiversitas 5 (2): 61–66.
Kahlil, A., M.S. Alwani, & A.K. Omar. 2006. Chemical composition and cell wall structure of Malaysian plant waste fiber. Bioresource 1 (2): 220–232.
Kromer K. 2009. Physical properties of flax fibre for non-textile-use. Res. Agr. Eng. 55 (2): 52–61.
Kronborg, M., C.A. Grandez, E. Ferreira & H. Baslev. 2008. Aphandra Natalia (Arecaceae) - a little known source of piassaba fibers from the western Amazon. Rev. peru. biol. 15 (supl.1): 103–113.
Lescure, J.-P., Emperaire, L. & C. Franciscon. 1992. Leopoldinia piassaba Wallace (Arecaceae): a few ecological and economic data from the rio Negro region (Brazil). For. Ecol. Manage. 55: 83–86.
McDougall G.J., I.M. Morrison, D. Stewart, J.D.B Weyers & J.R. Hillamn. 1993. Plant Fibres: Botany, Chemistry and processing for Industrial Use. J. Sci. Food Agric. 62: 1–20.
Mejia K. 1988. Utilization of palms in eleven mestizo villages of the Peruvian Amazon (Ucayali River, Department of Loreto). Adv. Econ. Bot. 6: 130–135.
Millán B. 2006. Arecaceae endémicas del Perú. en: León B. et al. (ed.), El Libro Rojo de las Plantas Endémicas del Perú. Rev. Peru. Biol., Número especial 13(2): 706–707.
Millán, B. & F. Kahn. 2010. Characterization of leaf anatomy in species of Astrocaryum and Hexopetion (Arecaceae). Rev. peru. biol. 17 (1): 81–94.
Moreno L., A. Parac & F. Ocon. 2005. Fiber characteristics of promising abaca (Musa textilis Nee) accessions in NARC [National Abaca Research Center] germplasm suited for specific industry and uses. Philippine Journal of Crop Science, Vol. 30 (1): 60.
Morvan C., C. Andème-Onzighi, R. Girault, D. Himmelsbach, A. Driouich & D. Akin. 2003. Building flax fibres: more than one brick in the walls. Plant Physiology and Biochemistry 41: 935–944.
Msahli S., Y. Chaabouni, F. Sakli & J.Y. Drean. 2007. Mechanical behavior of Agave americana L. Fibres: Correlation Between Fine structure and mechanical properties. J. Applied Sci. 7 (24): 3951-3957.
Rasheed S. & A. Dasti. 2003. Quality and mechanical properties of Plant Commercial Fibers. Pakistan J. Biol. Sci. 6 (9): 840-843.
Rowell R.M., J.S. Han, & J.S. Rowell. 2000. Characterization and factors effecting fiber properties. In Natural Polymers and Agrofibers Composites. E. Frollini (ed.). Embrapa Instrumentação Agropecuária.
Sanchis Gritsch C. & R.J. Murphy. 2005. Ultrastructure of fibre and parenchyma cell walls during early stages of culm development in Dendrocalamus asper. Annals of Botany 95: 619–629.
Schultes, R.E. 1977. Promising structural fiber palms of the Colombian Amazon. Príncipes, 21: 72–82.
Tomlinson P. 1990. The structural biology of palms. Oxford University Press. New York.
TAPPI, Technical Association for the Pulp and Paper Industries. 1978a. Alpha-, Beta and Gamma Cellulose in pulp. TAPPI Test Method T 203 os-74. TAPPI Press. Atlanta.
TAPPI, Technical Association for the Pulp and Paper Industries. 1978b. Acid-insoluble lignin in wood and pulp. TAPPI Test Method T 222 os-74. TAPPI Press. Atlanta.
Vormisto J. 2002. Making and marketing chambira hammocks and bags in the Village of Brillo Nuevo, Northeastern Peru. Econ. Bot. 56(1): 27–40.
Email: Betty Millán bmillans@gmail.com
Presentado: 30/06/2011
Aceptado: 08/07/2012
Publicado online: 01/10/2012
