










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. vol.19 no.3 Lima dic. 2012
TRABAJOS ORIGINALES
La liebre europea Lepus europaeus (Lagomorpha: Leporidae) una especie invasora en el Perú
European hare Lepus europaeus (Lagomorpha: Leporidae) an invasive species in Peru
Horacio Zeballos1,2*, César Medina2, Kateryn Pino2, Adolfo Mejía-Ríos3, Alexánder Pari2
1 Centro de Investigación para la Promoción de los Pueblos-Bienestar, Coop. Víctor Andrés Belaúnde I-8, Yanahuara, Arequipa, Perú.
2. Colección Científica-Museo de
Historia Natural de la Universidad Nacional San Agustín, Av. Alcides Carrión s/n, Arequipa, Perú.3 Universidad Nacional Agraria
La Molina, Av. La Molina s/n, La Molina, Lima, Perú.* Autor para correspondencia.
Resumen
Este trabajo describe la distribución actual de la liebre europea, Lepus europaeus, en el Perú, la que actualmente abarca la puna, valles andinos, alrededores del lago Titicaca e irrigaciones costeras; en los departamentos de Arequipa, Cusco, Moquegua, Puno y Tacna. En base a su distribución actual desarrollamos modelos de distribución potencial de esta especie, los que pronostican que esta especie llegaría al norte del Perú. Hacemos recomendaciones sobre los principales aspectos que debieran ser estudiados en el Perú sobre L. europaeus y las posibles consecuencias de su proceso invasivo en el territorio peruano.
Palabras clave: Especies invasoras; conservación; biodiversidad; puna; dispersión.
Abstract
We describe the current distribution of the European hare, Lepus europaeus, in Peru which currently covers the highlands, Andean valleys, surrounding areas of the Titicaca Lake and coastal irrigations; in Arequipa, Cusco, Moquegua, Puno and Tacna departments. Based on its current distribution we developed models of potential distribution of this species, which would forecast this species in northern Peru. We make recommendations on the main issues that should be studied in Peru, and the possible consequences of their invasive process of in Peru.
Keysword: Invasive Alien Species; conservation; biodiversity; puna; dispersion.
Introducción
La liebre europea, Lepus europaeus Pallas, 1778, fue introducida intencionalmente en Argentina y el sur de Chile entre 1880 y 1930 (Markham 1971, Carman 1976, Campos 1986, Grigera & Rapoport 1983). Actualmente la encontramos ocupando prácticamente todo el cono sur sudamericano, el altiplano Peruano-Boliviano, las áreas semidesérticas de las vertientes occidentales de los Andes y las áreas irrigadas de la costa sur del Perú; mostrando así que ha sobrepasado algunos límites y barreras biogeográficas para las especies nativas, como el río Tambo y los tablazos costeros (Pearson 1982, Sheffield & Thomas 1997, Yensen & Tarifa 2003, Marín et al. 2007). Las liebres, aunque prefieren primariamente hábitats abiertos, han demostrado una notable plasticidad ecológica en el uso de hábitats y recursos (Kufner et al. 2008), es por ello que las tierras altas de la Puna y los valles costeros parecen jugar un rol de importancia para su dispersión (Jaksic et al. 2002, Bonino et al. 2010). En contraste, los bosques amazónicos y montanos parecen ser una barrera efectiva que evita su expansión.
Esta especie fue reportada formalmente en el Perú desde el año 2002 para Tacna y Arequipa (Cossíos 2004 y Lleellish et al. 2007, citado por Cossíos 2010). Posteriormente su distribución fue ampliada para los alrededores del lago Titicaca en Puno y la puna sur de Cusco, tanto para localidades altoandinas por debajo de 4400 m, como para la costa de Tacna (Cruz 2005, Canales 2008, Bonino et al. 2010).
En el Perú la mayor parte de la información que se tiene sobre esta especie es anecdótica, y se conoce muy poco acerca de los efectos que causa en las comunidades que ha colonizado. Si bien, en algunas localidades de Tacna y en tres comunidades de la península de Capachica en Puno se han reportado daños de consideración sobre cultivos (Lleellish et al. 2007, Canales 2008), hasta la fecha no se conoce la real dimensión del problema. No obstante, es necesario considerar que estamos frente a una especie oportunista, que incluye principalmente a las gramíneas en su dieta y que utiliza una amplia gama de hábitats (Puig et al. 2007, Kufner et al. 2008). Bajo este panorama, cabe resaltar que hace una década atrás, las evidencias indicaban que la liebre solamente había logrado instalarse en las tierras menos productivas del país (Puna seca), sobreviviendo con pasturas de mala calidad; no obstante recientemente está ingresando a tierras de mayor productividad de la puna húmeda en los departamento de Cusco y Puno, donde existen hábitats más vegetados y con mayor riqueza de especies (Weberbauer 1945, Mostacero et al. 1996). De acuerdo a lo que se conoce sobre la biología de esta especie en Argentina y Chile (Carman 1976, Grigera & Rapoport 1983, Jaksic 1998, Mack et al. 2000, Iriarte et al. 2005, Puig et al. 2007, Kufner et al. 2008, Bonino et al. 2010), es posible que ocasionen algunos problemas en el Perú, tales como: competencia con especies nativas y ganado doméstico; modificaciones en la composición de las especies vegetales en las comunidades de pastos naturales, y con ello cambios en las propiedades físicas del ecosistema; alteraciones del ciclo de nutrientes; pérdida de la productividad vegetal; y pérdida de biodiversidad.
De acuerdo con la evidencia presentada sobre su dispersión y los daños que viene causando la liebre en el Perú, se trataría de una especie invasora, la cual pudiera estar amenazando los ecosistemas y hábitats, definitivamente con efectos económicos de consideración. Por estas razones, en este trabajo nos planteamos como objetivo determinar la distribución actual y potencial de la liebre europea en el Perú; y proporcionar elementos de juicio para definir las acciones a tomar frente a esta especie invasora.
Material y métodos
Distribución actual.-
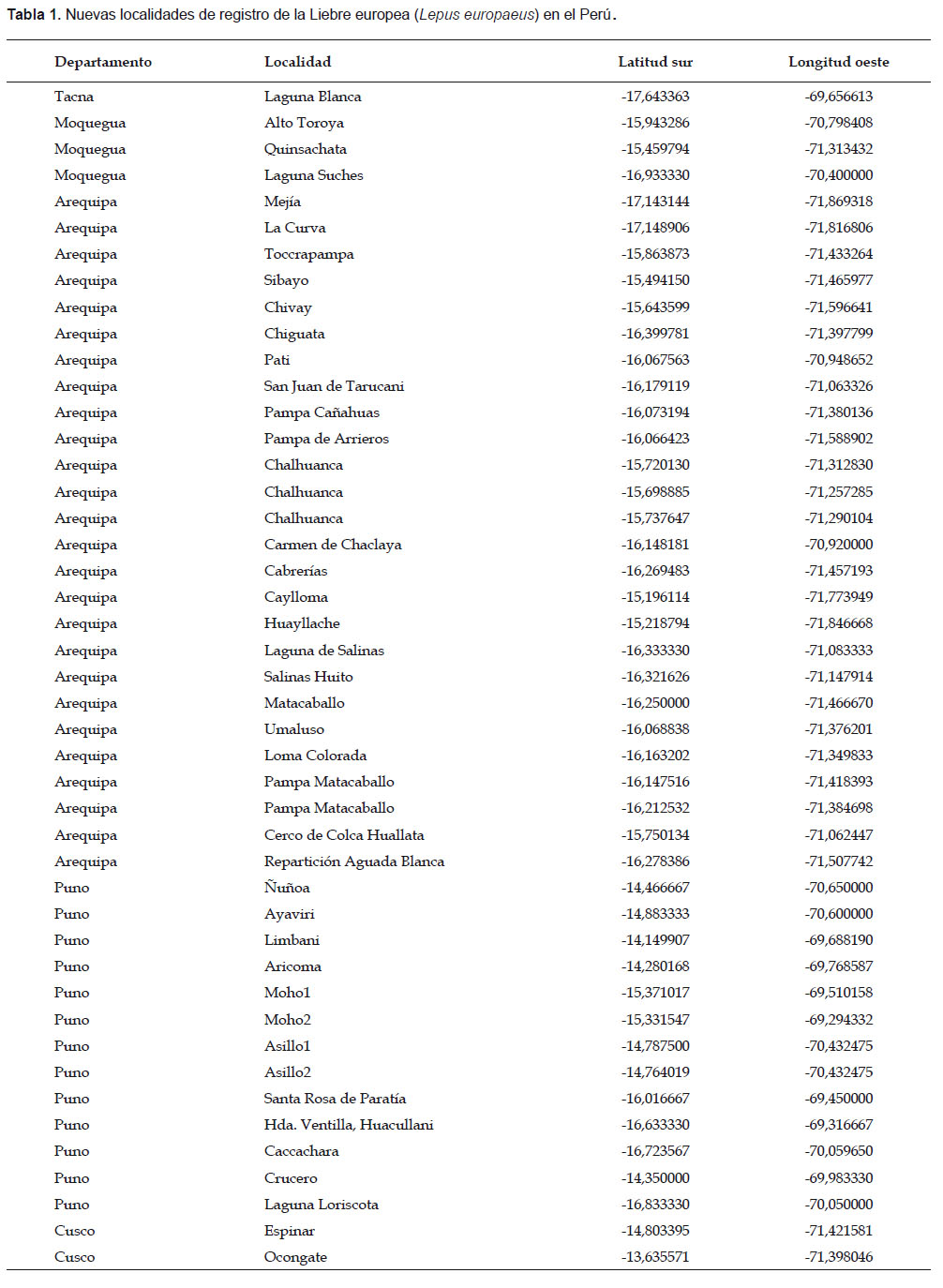
Recopilamos toda la información de campo y publicada sobre la presencia de la liebre europea en el Perú. La mayor parte de esta información en el Perú se basa en observaciones directas y comunicaciones personales, así como también, abundante evidencia física de su presencia en base a restos colectados en carreteras o de los pobladores y fotografías. Otra parte de la información está basada en nuestros registros de campo que documentan su presencia en el departamento de Arequipa desde el año 1999. Para toda esta información se obtuvieron las coordenadas geográficas de 63 registros de esta especie en Perú (46 de este reporte), los restantes provienen de la literatura (Canales 2008, Bonino et al. 2010), con ellos elaboramos el mapa de la distribución actual de la liebre (Fig. 1a). En el año de 1999 registramos la primera evidencia de liebre en Arequipa, así que durante este año y el 2008 realizamos búsqueda de restos o indicios de su presencia reciente en 30 localidades andinas y 5 entre la cabecera del río Tambo y su desembocadura, lo que nos permitió establecer dos rutas de dispersión, una por la puna y otra a lo largo de los valles de las vertientes occidentales y costa; información que también nos sirvió para calcular la velocidad de colonización en cada caso, la que fue estimada promediando la distancia lineal entre cada registro nuevo con el del año anterior, para los parámetros usamos el método de remuestreo de Jacknife (Sokal & Rohlf 2003). 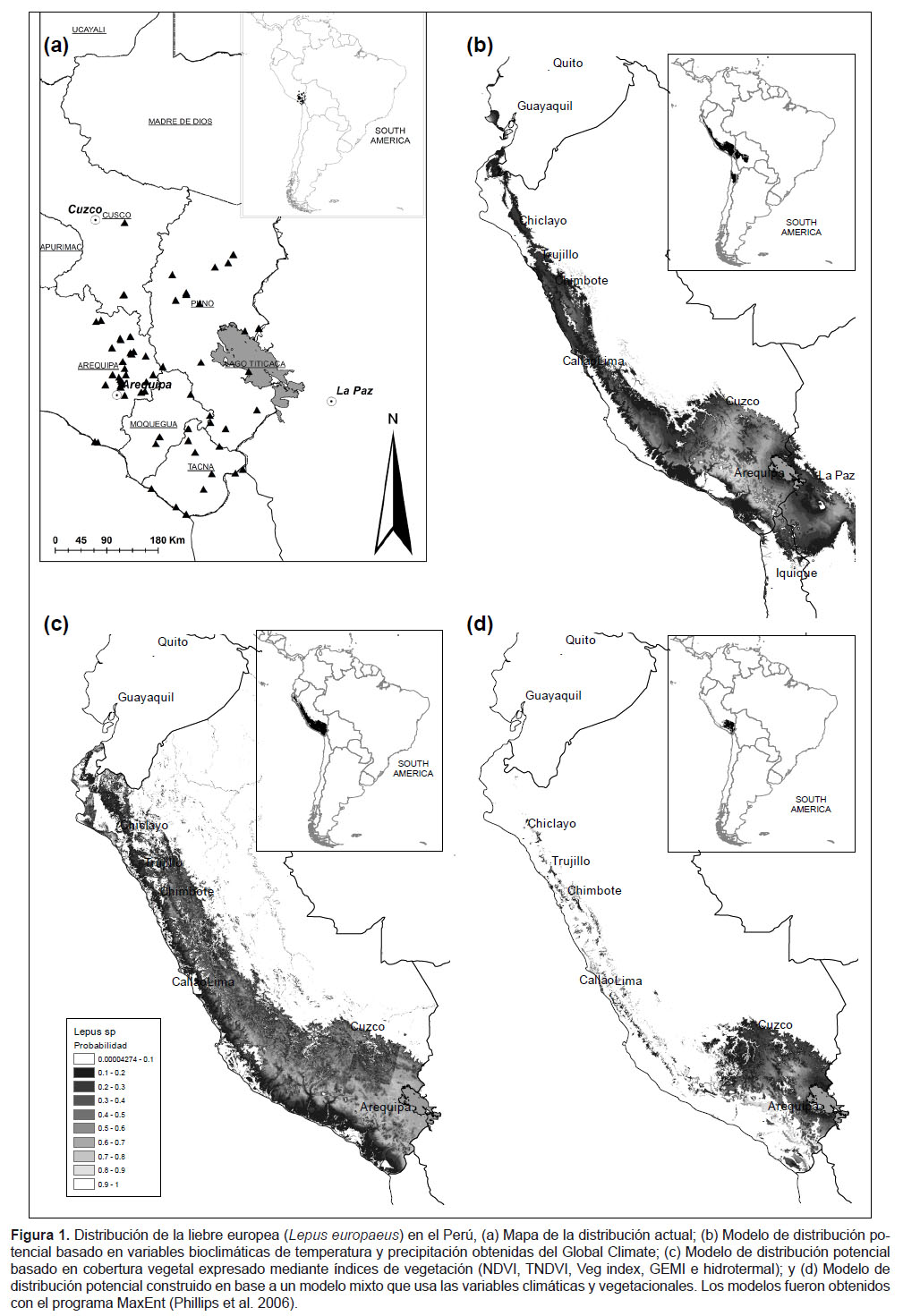
Distribución potencial.- Nos basamos en tres modelos, el primero utiliza 19 variables bioclimáticas basadas en temperatura y precipitación (Fig. 1b), obtenidas del Global Climate Data (World Clim 2011), esta es una base de datos de variables climáticas que se presenta en la forma de archivos raster, con una resolución de un kilómetro. El segundo modelo utiliza únicamente variables vegetacionales (Fig. 1c), específicamente asociados a la cobertura vegetal expresado mediante índices de vegetación (NDVI, TNDVI, Veg index, GEMI e hidrotermal), obtenidos a partir de un mosaico de imágenes landsat 5 TM, desarrolladas por uno de los autores: AM, estas fueron obtenidas del "Catálogo de Imágenes Satelitales" (INPE 2011-2012). Finalmente, el tercer modelo fue construido en base a un modelo mixto que usa las variables climáticas y vegetacionales descritas antes (Fig. 1d); en los tres casos usamos el método de máxima entropía (Benito & Peñas 2007) implementado en el programa Maxent (http://www.cs.princeton.edu/~schapire/maxent/) (Phillips et al. 2006, Phillips & Dudík 2008). Para configurar MaxEnt se introdujo 1000 como número máximo de iteraciones, estableciendo el límite de convergencia en 0,00001 y el valor de regularización en 0,0001. Estos valores, según el autor (Phillips et al. 2006), son adecuados para garantizar la convergencia del algoritmo. Para comparar la capacidad de discriminación de los distintos modelos seleccionmos el método del área bajo la curva, "Receiver Operating Characteristic" (ROC) (Hanley & McNeil 1982, Fielding & Bell 1997, Segurado & Araújo 2004, Muñoz & Felicísimo 2004) y en algoritmos de solo presencia (Phillips et al. 2006). El ROC indica que para un punto de presencia y uno aleatorio seleccionados al azar, la probabilidad de que el valor de idoneidad previsto por el modelo para el punto de presencia sea mayor que el previsto para el punto aleatorio. Es además una medida directa de la capacidad de discriminación del modelo, que toma valores próximos a 1 cuando existe un buen ajuste con los datos de evaluación y cercanos a 0.5 cuando el ajuste no es mejor que el obtenido por azar (Phillips et al. 2006).
Resultados
La liebre europea en el Perú, hacia diciembre del 2011 describe una amplia distribución en el altiplano y la costa sur, siempre en ecosistemas abiertos. Actualmente ocupa cinco departamentos del sur peruano (Arequipa, Cusco, Moquegua, Puno y Tacna), invadiendo prácticamente todo el altiplano del sur peruano, ha ingresado a las únicas tierras productivas que se ubican en el desierto, como las áreas ribereñas, irrigaciones y los humedales costeros (Fig. 1a, Tabla 1). En cuanto a su hábitat, nuestros datos muestran que ha ocupado principalmente las planicies y tierras onduladas con pajonales de puna, bofedales y tolares (Parastrephia spp.); así como también, en las tierras de cultivo a diferente altitud y áreas vegetadas que rodean las lagunas costeras como juncales y salicorniales (Salicornia sp.). No encontramos ningún registro se ha efectuado en áreas rocosas o escarpadas.
Los primeros avistamientos en la puna de Arequipa ocurrieron en 1999, en las localidades de Pati (16,0676 °S y 70,9487 °W, 4456 m), Chalhuanca (15,720 °S y 71,3128 °W, 4334 m) y Carmen de Chaclaya (16,1482 °S y 70,9226 °W, 4437 m), ambas cerca del límite con el departamento sureño de Moquegua. Desde ese año a la fecha, observamos dos rutas de dispersión: la primera se desarrolló de forma progresiva sobre la puna de Arequipa a una velocidad de dispersión de 14,5 km/año (n= 30; s= 0,04), a la cuál llamamos ruta puneña; mientras que la segunda ocurrió a lo largo de los ríos hacia la costa (especialmente en el río Tambo), en esta "ruta ribereña" la velocidad de dispersión fue de 18,2 km/año aproximadamente.
Los modelos de distribución presentan un valor de ROC mayor de 0,989 derivado de la base de datos proveniente de las 19 variables bioclimáticas, Lo que nos permite establecer que la distribución de los puntos de presencia de la liebre europea es consistente y/o homogénea para cada una de las variables climáticas tomadas en el modelo. Para los modelos surgidos con la base de datos de variables provenientes de la base de datos Mixta y de la derivada de los Índices de Vegetación, se tienen valores de ROC de 0,971 y 0,935 respectivamente. Los tres modelos teóricos son consistentes y sugieren que existiría disponibilidad de hábitat para que la liebre europea colonice el norte del país (Fig. 1 a, b, c).
Discusión
Según la evidencia publicada L. europaeus debe haber llegado al territorio peruano desde el sur, posiblemente de Chile, ya que los primeros registros se conocen de Tacna; pero no se descarta que haya ingresado por Bolivia, país con el que mantenemos una línea fronteriza mayor.
Los reportes más antiguos sobre la presencia de liebre europea en el territorio peruano fueron los de Lleellish et al. (2007) en Candarave y Yarada Media, Tacna, fechados en 1975 y 1989 respectivamente; Cruz (2005) estimó su llegada entre 1994 – 1996 y Cossíos (2004) en 1995-1998. Por su parte Cossíos (2010) postula que la estimación más confiable de su ingreso al Perú habría ocurrido en el lapso de 1994 - 1998 y que 1975 es una propuesta extrema, argumentando que a la velocidad de dispersión de 10 − 44 km/año (Grigera & Rapoport 1983, Cossíos et al. 2004, Bonino et al. 2010) habría alcanzado la costa antes de lo propuesto por Lleelish y colaboradores y que el 97% de los entrevistados por este último indicaron su presencia a partir de 1996. No obstante, usando el primer indicio certificado de la presencia de la liebre en Arequipa en 1999 y Asillo en 2002, (ubicados a aproximadamente 215 y 265 km en línea recta de la frontera de Tacna y Puno respectivamente), y usando la velocidad de dispersión propuesta en este estudio tenemos un estimado de ingreso que oscila entre 1980 - 1986 y 1979 - 1986 respectivamente. Considerando que estos estimados se basan en distancias lineales que no consideran la topografía y que los primeros colonos deben haber arribado en número reducido por lo que su presencia pudiera ser no percibida; proponemos reconsiderar el año 1975 como fecha probable de ingreso al país (Lleelish et al. 2007). Sin embargo, independientemente de la fecha y punto de ingreso al país, este debió haber ocurrido al sur del Lago Titicaca donde predominan dos tipos de ambientes, el primero circunlacustre más benigno y de menor altitud (3700 - 3800 m); y el segundo, la puna seca, con alturas promedio de 4300 m, con presencia de bofedales aislados, extensos pajonales y tolares que están ampliamente alterados por sobrepastoreo. No descartamos que el empobrecimiento de la vegetación haya facilitado su dispersión, porque habrían menos competidores y su plasticidad para sobrevivir en ambientes degradados lo favorecería.
La velocidad de dispersión estimada por nosotros de 14,5 ± 2,4 km/ año para la puna y 18,2 km/año a lo largo de los ríos es congruente con lo presentado los 18,6 km/año estimado por Griguera y Rapoport (1983) para datos experimentales, valores similares son reportados por Bonino et al. (2010) para poblaciones de Paraguay. Este último estima 30 km/año para poblaciones de Bolivia. Por su parte Cossíos (2004) y Bonino et al. (2010) han estimado velocidades de dispersión de 44,3 y 34 km/año, respectivamente, para la distancia entre Tarija y el borde sur del Perú.
En el caso de la dispersión por la puna observamos que permite una distribución latitudinal, mientras que la ribereña es altitudinal, por la que accederían a la costa desde las tierras altas. En las áreas abiertas de la Puna la velocidad de dispersión ha sido menor que a lo largo de los ríos. La diferencia observada en el avance en la Puna y la ribereña en nuestros datos, podría deberse a que a lo largo de los ríos el avance es lineal y unidireccional con un incremento proporcional en el área de ocupación, mientras que en la puna los desplazamientos pueden ocurrir en distintas direcciones por lo que la superficie cubierta sería cuadrática, y por lo tanto se daría a menor velocidad.
En esta misma línea de pensamiento, es importante destacar que los tres modelos, si bien tienen diferencias en el área proyectada, lo relevante es que predicen su dispersión hasta el norte del país, ya que posiblemente encuentren hábitat adecuado (tierras abiertas y pastizales), y donde prácticamente no existirían barreras biogeográficas ni fisiológicas para su dispersión. En el caso del modelo mixto (Fig. 1d) nos describe una estrecha franja en el lado occidental de la cordillera que puede deberse a que está sesgado por la información de la puna seca del altiplano sur; no obstante nos inclinamos a pensar que podría ser una franja de hábitat óptimo para esta especie, característicamente árido, con una baja población de depredadores, clima benigno, altamente alterado y con pocos competidores herbívoros.
De acuerdo a lo observado en estudios previos (Elton 1958, Jacksic 1998, Levine & D’Antonio 1999, Mack et al. 2000, Arim et al. 2006), las áreas más propensas para la presencia de las especies invasoras como la liebre europea serían: a) áreas perturbadas por el hombre, como las pasturas de la puna y áreas ribereñas a lo largo de los valles de las vertientes occidentales; b) sistemas pobres sin interacciones arraigadas, como en muchas áreas de la puna; c) sistemas donde hay más hábitats o nichos que eventualmente pueden ser ocupados, tal es el caso de la alta variabilidad orográfica en la puna y el gradiente altitudinal de los valles costeros.
En cuanto a la resistencia a las invasiones, Elton (1958) postuló que crece en proporción directa al número de especies en la comunidad, por que a mayor riqueza hay mayor estabilidad. Concordantemente con esto cabe mencionar que las áreas más ricas en especies en la puna están ubicadas en las áreas rocosas y los bofedales, en cambio, las pampas por lo general alteradas por sobrepastoreo, presentan un menor número de especies, y es precisamente donde es más fácil de encontrar a la liebre.
La puna Sudamericana es un área extensivamente utilizada por los pobladores desde muy antiguo (García & Beck 2006), presentándose las mayores perturbaciones por sobrepastoreo en la puna seca, lo cual ha desencadenado serios procesos de desertificación (Zeballos 2008).
Si bien la puna es un área altamente diversa, con complejos ensambles de vertebrados y grupos faunísticos muy diversos como los roedores (Pearson 1982, Pacheco et al. 2009), se esperaría que la riqueza fuera una efectiva barrera, tal como lo predice la hipótesis de Elton (1958); no obstante, en la puna la diversidad se encuentra asociada a ciertas localidades existiendo extensas áreas muy pobres donde las interacciones son reducidas; esto no contradice la hipótesis de Elton, sino que la refuerza, ya que la alta riqueza para ser una barrera efectiva tendría que ser continua, no como el caso de la puna donde hay un mosaico de hábitats con diferente grado de riqueza.
Si bien, la liebre encuentra en la puna una continuidad de hábitats adecuados para su dispersión, su presencia en la serranía esteparia facilitaría su dispersión entre los ríos y es lo que demuestran los modelos presentados. Por su parte, el desierto parece ser una barrera infranqueable para esta especie, y su llegada a las irrigaciones y humedales costeros habría sido facilitada por los valles de la cuenca pacífica, los que han sido utilizados como vías de dispersión.
Es notable que a lo largo de estos ambientes la liebre tiene condiciones adecuadas para su expansión, ya que encontramos: a) disponibilidad de áreas perturbadas con buen alimento para su supervivencia (v.g. cultivos); b) ausencia de depredadores naturales efectivos y parásitos, tanto zorros, gatos y pumas aún no han aprendido a cazarlas, tal como ocurrió en el sur del Continente (Jaksic 1998) donde actualmente las liebres se constituyen en importante dieta de varios depredadores (Zanón-Martínez et al. 2012); y c) una reducida presencia de competidores nativos, muchos de los que se encuentran en proceso de declinación (venados, guanacos, viscachas) (Novaro et al. 2000, Farías & Kittlein 2008).
La pregunta crucial en estos momentos es: ¿Qué pasará cuando lleguen a tierras más productivas del centro y norte del país? De acuerdo con su alta capacidad de dispersión, no debería tener dificultades para seguir avanzando hacia el norte, inclusive hasta la depresión de Huancabamba (Bonino et al. 2010), recorriendo vastas áreas de puna perturbada, la que hacia el norte se hace más productiva y está ampliamente usada en agricultura. Inclusive llegaría a las irrigaciones costeras dispersándose a lo largo de los valles y las lomas costeras. Esto es motivo de una amplia preocupación, pues al llegar a tierras de mayor productividad, donde podrían alcanzar altas densidades en torno a la capacidad de carga del sistema; podría ocurrir una explosión poblacional diferente de las reportadas en el Perú con pequeños roedores (Gilmore 1947, Pearson 1975, Zeballos et al. 2000), y que podría ser muy dañina, causando daños como los que ya se han visto en Tacna y Puno. Lamentablemente, hasta la fecha en el Perú muy poco se está investigando sobre esta especie invasora, siendo necesario conocer como se está comportando, y poder así plantear propuestas objetivas y debidamente sustentadas para resolver los conflictos e impactos que genere (Ojasti 2001).
Por estas razones es urgente que se tomen medidas para controlar la propagación de L. europaeus y establecer las decisiones a tomar ante su eminente llegada al centro y norte del Perú. Asimismo, es necesario que los organismos gubernamentales financien y promuevan investigaciones con el fin de conocer la real magnitud del efecto que tiene su presencia en el Perú, para que se conozca: a) su efecto sobre la diversidad biológica (herbivoría, competencia y depredación) y sus implicancias para la conservación de esta biodiversidad; b) el efecto que tiene sobre la composición y la productividad vegetal; c) el efecto sobre el funcionamiento de los ecosistemas; d) el impacto sobre los productores en las áreas afectadas; y e) la posibilidad de aprovechamiento como recurso alimenticio y peletero como ha sido hecho en otros países (Iriarte et al. 2005). Esperamos que el presente trabajo sirva de base para iniciar la toma de decisiones que definan los principios y metodologías para monitorear los impactos de L. europaeus, plantear una estrategia para su control; y prevenir sus efectos sobre nuestros ecosistemas.
Agradecimientos
Agradecemos al Dr. Ariel Farías por la revisión y notables aportes al manuscrito; al Museo de Historia Natural de la Universidad Nacional de San Agustín de Arequipa por el acceso a los especímenes allí depositados; a los pobladores por compartir su información; a los profesionales del Centro de Estudios y Promoción del Desarrollo–desco, por la información facilitada y por el uso del Software ArcGIS (Licencia: KEY 555983330); al INPE de Brasil por el uso de las imágenes Landsat; y al World Clim por el uso de la base de datos bioclimáticos.
Literatura citada
Arim M., S.R. Abades, P.E. Neill, M. Lima, & P.A.Marquet. 2006. Spread dynamics of invasive species. Proceeding of the National Academy of Science 103: 374-378. [ Links ]
Benito de Pando, B. & J. Peñas de Giles. 2007. Aplicación de modelos de distribución de especies a la conservación de la biodiversidad en el sureste de la Península Ibérica, GeoFocus (7): 100-119. [ Links ]
Bonino N., D. Cossíos & J. Menegheti. 2010. Dispersal of the European hare, Lepus europaeus in South America. Folia Zool. 59(1): 9-15. [ Links ]
Campos H. 1986. Mamíferos terrestres de Chile. Valdivia: Corporación Nacional Forestal y Marisa Cuneo Ediciones. [ Links ]
Canales A. 2008. Evaluación poblacional de Lepus europaeus Pallas, 1778 sus efectos en la agricultura de la península de Capachica. Revista de Investigaciones 4(2): 187-198. [ Links ]
Carman R.L. 1976. En torno a la liebre y su introducción en la Argentina. Diario La Prensa, Buenos Aires. [ Links ]
Cossíos D. 2004. La liebre europea Lepus europaeus (Mammalia, Leporidae), especie invasora en el sur del Perú. Rev. Per. Biol. 11(2): 209-211. [ Links ]
Cossíos, E.D. 2010. Vertebrados naturalizados en el Perú: historia y estado del conocimiento. Revista Peruana de Biología, 17(2):179-189. [ Links ]
Cruz A. 2005. Efectos del establecimiento y dispersión de la liebre europea, Lepus europaeus (Mammalia: Leporidae) en Tacna, Perú. Revista Scientiae (Universidad Jorge Basadre Grohmann, Tacna) 1:41-42. [ Links ]
Elton C.S. 1958. The Ecology of Invasions by Animals and Plants. Edit. Chapman and Hall. London. [ Links ]
Farias A.A. & M.J. Kittlein. 2008. Small-scale spatial variability in the diet of pampas foxes (Pseudalopex gymnocercus) and human-induced changes in prey base. Ecological Research 23 (3): 543–550. [ Links ]
Fielding, A.H. & J.F. Bell. 1997. A review of methods for the assessment of prediction errors in conservation presence/absence models, Environmental Conservation, 24: 38-49. [ Links ]
García E. & S.G. Beck. 2006. Puna, 51-76 pp, en M. Moraes, R.B. Øllgaard, L.P. kvist, F. Borchsenius H. Balslev (eds.) Botánica Económica de los Andes Centrales, Universidad Mayor de San Andrés, La Paz. [ Links ]
Gilmore R. M. 1947. Cyclic bahavior and economic importance of the Rata-muca (Oryzomys) in Peru. Journal of Mammalogy 28(3): 231–241. [ Links ]
Grigera D.E. & E.H. Rapoport 1983. Status and distribution of the European hare in South America. Journal of Mammalogy 64:163-166. [ Links ]
Hanley, J.A. & B. J. McNeil. 1982. The meaning and use of the area under a receiver-operating characteristic (ROC) curve, Radiology 143: 29-36. [ Links ]
INPE. 2011-2012. Catálogo de Imagens. Instituto Nacional de Pesquisas Espaciais. Ministèrio da Ciència, Tecnologia e Inovaçáo, Brasil. www.inpe.br/CDSR/ [ Links ]
Iriarte J.A., G.A. Lobos & F.M. Jaksic. 2005. Invasive vertebrate species in Chile and their control and monitoring by governmental agencies. Revista Chilena de Historia Natural, 78: 143-154. [ Links ]
Jaksic F. 1998. Vertebrate invaders and their ecological impacts in Chile. Biodiversity and Conservation 7: 1427-1445. [ Links ]
Jaksic M.F., A. Iriarte, J. Jiménez, & D. Martínez D. 2002. Invaders without frontiers: cross border invasions of exotic mammals. Biological Invasions 4: 157–173, 2002. [ Links ]
Kufner L., M.B. Sepúlveda, G. Gavier, L. Madoery, & L. Giraudo. 2008. Is the native deer Mazama gouazoubira threatened by competition for food with the exotic hare Lepus europaeus in the degraded Chaco in Córdoba, Argentina? Journal of Arid Environments 72 (2008) 2159–2167 [ Links ]
Levine J.M. & C.M. D’Antonio. 1999. Elton revisited: A review of evidence linking diversity and invasibility. Oikos 87: 15-26. [ Links ]
Lleellish M., R. Cadenillas & G. Chipana. 2007. Presencia de liebre europea Lepus europaeus en el Departamento de Tacna. Serie de Publicaciones de Flora y Fauna. INRENA, Lima, Perú. (Citado por Cossíos 2010) [ Links ]
Mack R.N., D. Simberloff, W.M. Lonsdale, H. Evans, M. Clout, & F. Bazzaz. 2000. Invasiones Biológicas: Causas, Epidemiología, Consecuencias globales y Control. Tópicos en Ecología 5: 1-19. [ Links ]
Marin J.C, C.S. Casey, M. Kadwell, K. Yaya, D. Hoces, J. Olazabal, R. Rosadio, J. Rodríguez, A. Spotorno, M.W. Bruford & J.C. Wheeler. 2007. Mitochondrial phylogeography and demographic history of the Vicuna: implications for conservation. Heredity 99(1): 70-80. [ Links ]
Markham B.J. 1971. Catálogo de los anfibios, reptiles, aves y mamíferos de la provincia de Magallanes (Chile). Publ. Inst. Pat. (Chile), Ser. Monogr. 1, 1±64. [ Links ]
Mostacero J., F. Mejía & F. Peláez. 1996. Fitogeografía del norte del Perú. Serie Ciencias CONCYTEC, N° 14: 406 pp. [ Links ]
Muñoz, J. & A.M. Felicísimo. 2004. Comparison of statistical methods commonly used in predictive modelling, Journal of Vegetation Science, 15: 285-292. [ Links ]
Novaro A.J., M.C. Funes & R.S. Walker. 2000. Ecological extinction of native prey of a carnivore assemblage in Argentine Patagonia. Biological Conservation 92: 25-33. [ Links ]
Ojasti J. 2001. Especies Exóticas Invasoras. Estrategia Regional de Biodiversidad para los Países del Trópico Andino. Convenio de Cooperación Técnica no Reembolsable ATN/JF-5887-RG CAN – BID. 1-63 pp. [ Links ]
Pacheco V.R. Cadenillas, E. Salas, C. Tello & H. Zeballos. 2009. Diversidad y Endemismo de los Mamíferos del Perú. Revista Peruana de Biología 16: 5-32 [ Links ]
Pearson O.P. 1975. An outbreak of mice in the coastal desert of Peru. Mammalia, t 39(3): 375-386 [ Links ]
Pearson O.P. 1982. Distribución de pequeños mamíferos en el altiplano y los desiertos del Perú. Actas 8vo. Congreso Latinoamericano de Zoología. Mérida, Venezuela. 263-284 pp. [ Links ]
Phillips S.J., R.P. Anderson & R.P. Schapire. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190(3/4): 231-259. [ Links ]
Phillips S.J. & M. Dudík. 2008. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31: 161-175. [ Links ]
Puig S., F. Videla, M.I. Cona & S.A. Monge. 2007. Diet of the brown hare (Lepus europaeus) and food availability in northern Patagonia (Mendoza, Argentina). Mammalian Biology 72(4): 240–250 [ Links ]
Sheffield S. & H. Thomas. 1997. Mustela frenata. Mammalian Species, American Society of Mammalogists N° 570: 1–9. [ Links ]
Segurado, P. & M.B. Araújo. 2004. An evaluation of methods for modelling species distributions, Journal of Biogegraphy 31: 1555-1568. [ Links ]
Sokal, R. R. & F. J. Rohlf. 2008. Biometry, The principles and practice of statistics in biological research. 8va. Edición. Edit. W.H. Freeman and Company. Nueva York. [ Links ]
Weberbauer, A. 1945. El mundo vegetal de los Andes Peruanos, Editorial Lumen. Lima [ Links ]
World Clim. 2011. Global Climate Data. Versión 1.4. www.worldclim.org/ [ Links ]
Yensen E. & T. Tarifa. 2003. Galictis cuja. Mammalian Species 728: 1-8. [ Links ]
Zanón-Martínez J.I, A.Travaini, S. Zapata D. Procopio & M.A. Santillán. 2012. The ecological role of native and introduced species in the diet of the puma Puma concolor in southern Patagonia Oryx, 46(1): 106–111. [ Links ]
Zeballos H. 2008. Conservación y desarrollo sostenible en la puna seca de América del Sur: manejo de recursos naturales en la Reserva Nacional de Salinas y Aguada Blanca. Perú Hoy año 2008(2): 272-282. [ Links ]
Zeballos H., A. Cornejo, L. Villegas, A. Ortega, K. Molina & R. Gutiérrez. 2000. Densidad poblacional de pequeños mamíferos en una "ratada" en las lomas de Atiquipa - Arequipa, Perú 1998. Actas IV Congreso Latinoamericano de Ecología, octubre 1998, Arequipa, Perú. [ Links ]
Correspondencia: Centro de Investigación para la Promoción de los Pueblos-Bienestar, Coop. Víctor Andrés Belaúnde I-8, Yanahuara, Arequipa, Perú .
Email: horaciozeballos@gmail.com
Presentado: 02/06/2012
Aceptado: 14/10/2012
Publicado online: 15/01/2013
