










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. vol.19 no.3 Lima dic. 2012
TRABAJOS ORIGINALES
Weddellian marine/coastal vertebrates diversity from a basal horizon (Ypresian, Eocene) of the Cucullaea I Allomember, La Meseta formation, Seymour (Marambio) Island, Antarctica
Diversidad de vertebrados marino costeros de la Provincia Weddelliana en un horizonte basal (Ypresiano, Eoceno) del Alomiembro Cucullaea I, Formación La Meseta, isla Seymour (Marambio), Antártida
Marcelo A. Reguero1,2,3,*, Sergio A. Marenss1,3and Sergio N. Santillana1
1 Instituto Antártico Argentino, Cerrito 1248, C1010AAZ Ciudad Autónoma de Buenos Aires, Argentina.
2 División Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n, B1900FWA, La Plata, Argentina.
3 Consejo Nacional de Investi
gaciones Científicas y Técnicas, Argentina (CONICET).* Corresponding author.
Abstract
The La Meseta Formation crops out in Seymour/Marambio Island, Weddell Sea, northeast of the Antarctic Peninsula and contains one of the world's most diverse assemblages of Weddellian marine/coastal vertebrates of Early Eocene (Ypresian) age. The La Meseta Formation is composed of poorly consolidated, marine sandstones and siltstones which were deposited in a coastal, deltaic and/or estuarine environment. It includes marine invertebrates and vertebrates as well as terrestrial vertebrates and plants. The highly fossiliferous basal horizon (Cucullaea shell bed, Telm 4 of Sadler 1988) of the Cucullaea I Allomember is a laterally extensive shell bed with sandy matrix. The fish remains, including 35 species from 26 families, of the Ypresian Cucullaea bed represent one of the most abundant and diverse fossil vertebrate faunas yet recorded in southern latitudes. Stratigraphic distribution and phylogenetic relationships of the Weddellian sphenisciforms are consistent with a first radiation of this group in the Early Eocene. The first inquestionable archaeocete from Antarctica is recorded in this unit and is referred to a new taxon.
Keywords: Antarctica; Seymour Island; Early Eocene (Ypresian); La Meseta Formation (Cucullaea I Allomember); Vertebrates; Paleobiogeography
Resumen
La Formación La Meseta aflora en la Isla Seymour/Marambio, Mar de Weddell, noreste de la Península Antártica y contiene una de las asociaciones de vertebrados costeros/marinos de edad Eoceno temprano (Ypresiano) más diversa que se conoce a nivel mundial. Esta unidad está compuesta por areniscas marinas pobremente consolidadas las cuales fueron depositadas en ambientes costeros, deltaicos y/o estuarinos. Esta incluye invertebrados y vertebrados marinos así como plantas y vertebrados terrestres. El horizonte basal (el banco de Cucullaea, Telm 4) del Alomiembro Cucullaea I es lateralmente extensor y altamente fosilífero. Los restos de peces del banco de Cucullaea (Ypresiano) incluyen 35 especies con 26 familias y representa una de las más abundantes y diversas fauna de vertebrados fósiles registradas en latitudes altas. La distribución estratigráfica y las relaciones filogenéticas de los pingüinos fósiles (Sphenisciformes) son consistentes con la primera radiación de este grupo en el Eoceno temprano. El primer incuestionable Archaeoceti de Antártida es registrado en esta unidad y es referido un nuevo taxón.
Palabras claves: Antártida; Isla Seymour; Eoceno temprano (Ypresiano); Formación La Meseta (Alomiembro Cucullaea I); Vertebrados; Paleobiogeografía.
Introduction
Paleogene Antarctic marine/coastal vertebrates come almost exclusively from the James Ross Basin, Wedddell Sea, Antarctic Peninsula (Reguero & Gasparini, 2007), mostly from Early Eocene-earliest Oligocene? fossils of the La Meseta Formation in Seymour (Marambio) Island (Fig. 1), secondly from the Eocene of the Fildes Peninsula, 25 de Mayo (King George) Island (Covacevich & Rich 1982; Li Jianjun & Zhen Shuonan 1994), and from Eocene erratics of the McMurdo Sound, East Antarctica (Stilwell & Zinsmeister 2000).
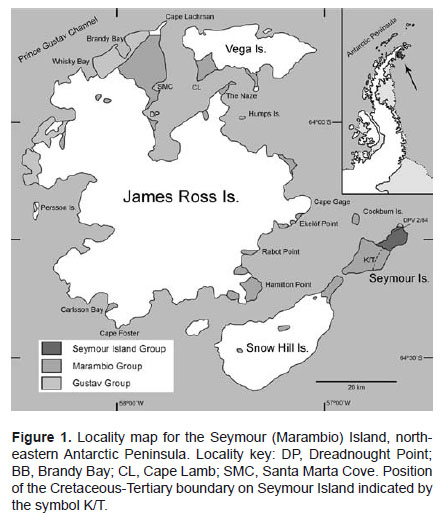
The record of fossil vertebrates in the La Meseta Formation is extremely diverse. In this paper we will refer only to the basal horizon of the Cucullaea I Allomember of that formation (Marenssi et al. 1998a, b). This horizon is characterized by a laterally extensive shell bed (Cucullaea shell bed). During the austral summers of 1990–2000 Argentinean teams’ recovered more than 10,000 teeth of fishes by dry-sieving and surface-prospecting of different localities along this horizon.
This first description of the Ypresian basal horizon marine/coastal vertebrate fauna documents the previously unappreciated diversity and unique character of Antarctica’s Early Eocene marine vertebrates, and indicates a cool-water paleoenvironment for the marine vertebrate assemblage.
Biogeographically all these vertebrates lived in the Weddellian Province. The Weddellian Province was proposed by Zinsmeister (1979, 1982) as a biogeographic unit of shallow marine waters that encompassed the coasts of Australia, New Zealand, Tasmania, Antarctica and southern South America (Magallanic Region in Chile, and Patagonia in Argentina) during the late Cretaceous through Eocene.
Locality and geological setting
The Early Eocene to earliest Oligocene? La Meseta Formation (Elliot & Trautman 1982) crops out in Seymour and Cockburn islands, close to the northern tip of the Antarctic Peninsula, Antarctica and is an unconformity-bounded unit (La Meseta Allo-Alloformation of Marenssi et al. (1998a). This unit is the topmost
exposed part of the sedimentary fill of the Late Jurassic-Tertiary James Ross Basin (Del Valle et al. 1992) and was interpreted as the filling of an incised-valley system (Marenssi et al. 1998a). The La Meseta Formation rests unconformably on either the Late Cretaceous López de Bertodano Formation or on the Paleocene Sobral and Cross Valley formations (Sadler 1988; Marenssi et al. 1998a). The unit is composed of sandstones and mudstones with interbedded shell-rich conglomerates. It was subdivided by Sadler (1988) into seven lithofacies units (Telms 1-7), and later organized into six erosionally-based internal units, named from base to top Valle de Las Focas, Acantilados, Campamento, Cucullaea I, Cucullaea II and Submeseta Allomembers (Marenssi et al. 1998a) (Fig. 2). These units were deposited mainly during the Eocene in deltaic, estuarine and shallow marine settings, mostly within a northwest-southeast trending incised valley (Marenssi et al. 1998a, 1998b). 
The basal part of the Cucullaea I Allomember is a horizon dominated by the pelecypod Cucullaea and darwinellid gastropods. Sadler (1988) characterized this shell bed (its Telm 4) by its thickness (as thick as 3 m), coarseness, and relatively high content of phosphatic teeth and bones, Sadler (1988) interpreted it as a transgressive lag characterized by abundant phosphate pebbles and glauconite. The age of this horizon of ca. 52.5 Ma (Ypresian) is indicated by the strontium stratigraphy based on the 87Sr/86Sr ratios of carbonate shells of the overlying beds of the Cucullaea I Allomember and it is consistent with paleomagnetic and biostratigraphic data (Montes et al. 2010)(Fig. 2).
The Cucullaea I shell bed represents a laterally continuous (several kilometers) horizon with thicknesses up to 3 meters. It bears complex internal reactivation (erosion-sedimentation) surfaces indicating multiple events. Individual beds are lens-shaped (up to 0.5 m thick) with erosional bases. They are mostly matrix supported, composed by bioclasts and some gravels immersed in a coarse-sand matrix with some granules. Internally each bed is massive or present through cross bedding or rarely parallel bedding.
Shells are disarticulated and oriented with the maximum projection area parallel to bedding. Concave-up, concave-down and nesting are common. Reworking of the underlying deposits (time-averaging) is suggested by fossil shells and fishes. There are a mixture of whole (although disarticulated) shells mostly of the species Cucullaea raea reworked directly from the underlying sands (Telm 3 or Campamento Allomember) and higly abraded and broken shells of many other invertebrate groups (Fig. 3).

Sedimentological evidences indicate that the Cucullaea I shell bed represents channel lags of laterally migrating subtidal channels (inlets?) developed at the mouth of a tide and wave influenced estuary (Marenssi et al. 1998b) during a slow transgressive event.
Many authors (Sadler 1988; Marenssi et al. 1998b; Ivany et al. 2006) indicated that this horizon bears all the hallmarks of a time-condensed transgressive deposit and placed a major hiatus at its base. Although the time averaging and erosion of the underlying beds are evident new stratigraphic data (Montes et al. 2010) suggest no hiatus but slow accumulation rate.
Material and methods
Surface collecting was carried out over several kilometers of La Meseta Formation exposures. Numerous fossils were recovered by surface collecting, although the majority of the specimens were collected through screen sieving and limited excavation. Approximately 1300 kg of matrix was collected and dry sieved in the field. The residues were sorted to 850 microns, with the high quantity of glauconite and ferruginous grains preventing sorting of finer fractions.
Dry-sieving sediments from a single site, DPV (División Paleontología de Vertebrados) 2/84 (64°14’21.782"S; 56°36’11.685"W, Cucullaea shell bank, basal part of the Cucullaea I Allomember, Fig. 1), has produced hundreds of bone fragments and teeth of sharks, rays, chimaeroids, bony fishes, turtles, penguins, whales, terrestrial mammals (marsupials and meridiungulates) and trunks and leaves (Reguero et al. 2002). The diversity of this vertebrate-bearing horizon can be taken to represent essentially a single marine fauna.
All the material described here is deposited in the fossil vertebrate collection of the Museo de La Plata (División Paleontología de Vertebrados) and have registration numbers prefixed by "MLP".
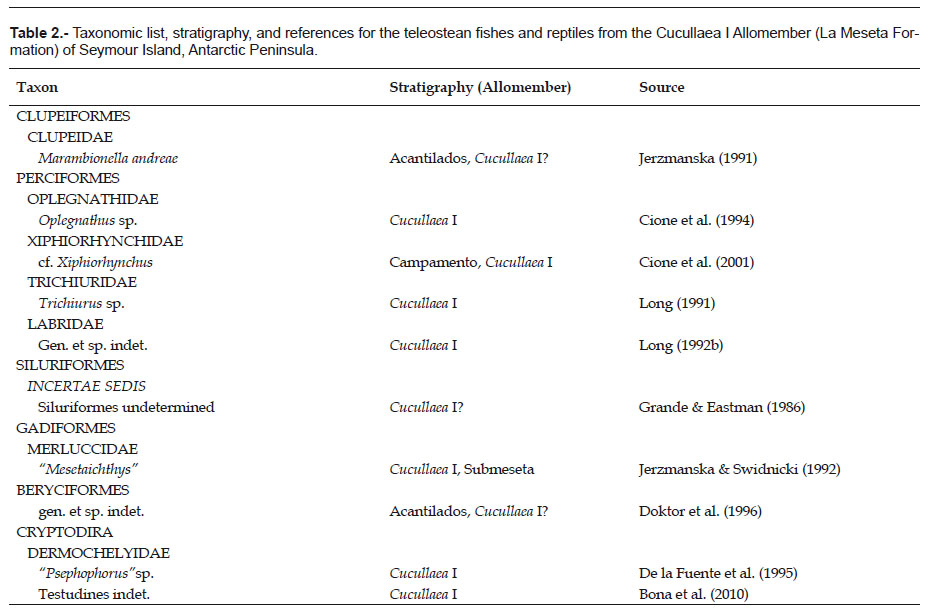
Taxonomic definitions and terminology. The fossil fish material collected consists of isolated remains of selachians and actinopterygians and was obtained by dry sieving of unconsolidated sediments of the Cucullaea
bed (Telm 4). The systematic and nomenclature used here is adopted from Cappetta (1987) and Compagno (1988).For Sphenisciformes Clarke et al. (2003) proposed phylogenetic definitions for higher taxa within the penguin total group. Pansphenisciformes is applied to the clade including all taxa more closely related to Spheniscidae than any other extant avian lineage. Sphenisciformes is applied in a more exclusive sense to the clade including all Pansphenisciformes that share the apomorphic loss of aerial flight. Spheniscidae is applied to the crown clade of penguins, comprising the most recent common ancestor of all living penguin species and its descendants. We use the classification proposed by Simpson (1946, 1971) throughout this paper.
Here, we follow the terminology and sequential stratigraphic interpretation of the revision of Marenssi et al. (1998a, 1998b).
Results
Ypresian diversity of marine/coastal vertebrates in high latitudes
Particularly the Ypresian horizon of the Cucullaea I Allomember contains the bulk of the fossil fish localities of the La Meseta Formation. This fossil fish fauna consists of thousands of selachian teeth and bony fish remains (including teeth, cranial fragments, vertebrae, fin spines, otoliths).
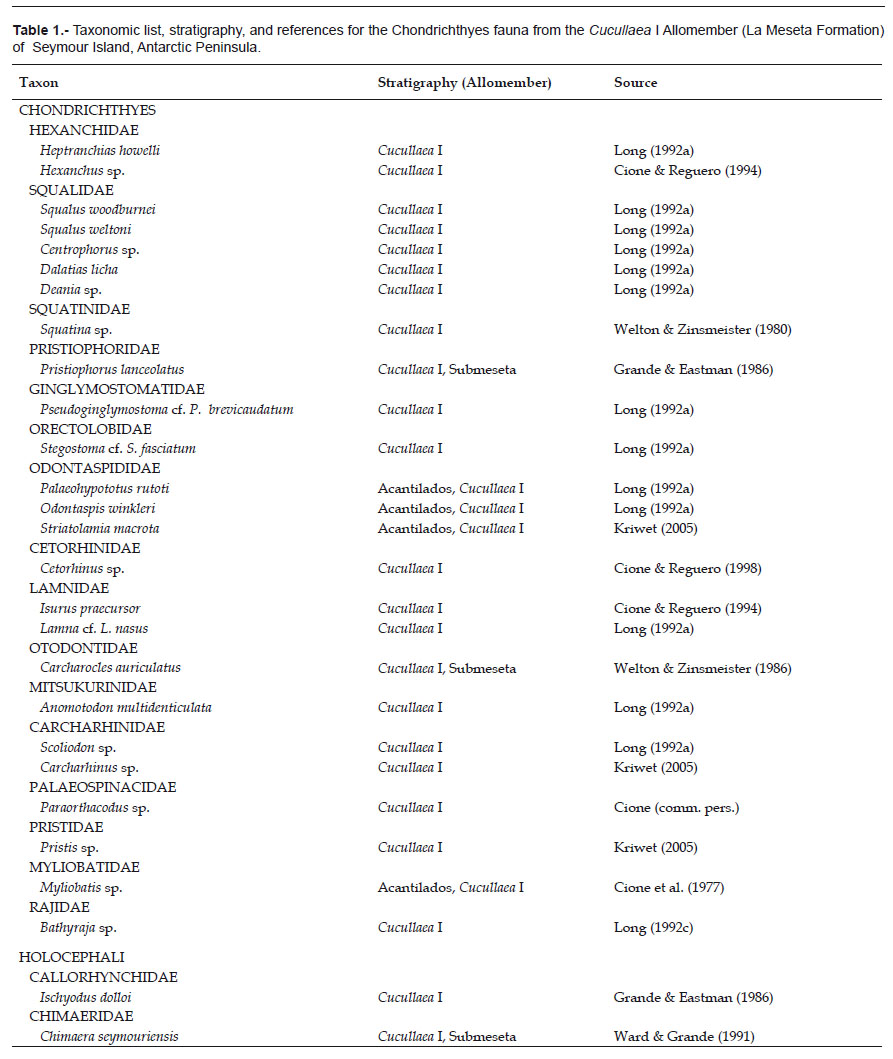
Sharks remains, largely represented by isolated teeth and vertebrae and a few poorly preserved dorsal spine, are extremely abundant and diverse in the coarser sand facies of the Cucullaea I Allomember. Interestingly, the level of diversity from a single locality (DPV 2/84) of the La Meseta Formation is much higher than the level of diversity for most extant cool temperate shark faunas and nearly equal to a present-day tropical shark fauna. At least 21 taxa of sharks between 11 families (Table 1) occur in this horizon. The most abundant (according to tooth number) elamosmobranch taxa are Squatina (37.89%), Pristiophorus (22.45%), Odontaspididae (17.24%), Myliobatis (6.70%), Squalus (6.81%), Rajidae (4.72) and Holocephali (2.68%).
Long (1992a) pointed out that this Antarctic fossil shark assemblage constitutes a very complex assortment of sharks from many different habitats converging on one specific locality.
The rare occurrence of Carcharhinus and Pristis, both taxa confined today to warm-temperate to tropical waters, in the temperate waters of Early Eocene in Antarctica indicates that both were not primary inhabitants but migrated along open trans-equatorial seaways into Southern Hemisphere waters (Kriwet 2005).
The Eocene selachian fauna from Antarctica includes 25 species in 16 families (Table 1). 24 taxa and 15 families come from the Cucullaea I Allomember of the La Meseta Formation of Seymour Island. The majority of taxa belong to sharks while batoids are represented by only three taxa with a very uneven distribution in the sequence. Long & Stilwell (2000) reported the presence of rare selachian teeth from Eocene deposits of Mount Discovery in East Antarctica. This material includes the first record of Galeorhinus for Antarctica.
Most of the fish taxa mentioned here are found in many different overlying localities and horizons of the Cucullaea I Allomember excepting in the upper units that document a sharp decrease in diversity near the boundary Cucullaea II and Submeseta allomembers. In the Submeseta Allomember, there is a dramatic diminution of diversity of those selachians that dominated below, there are no teleost taxa characteristic of warm water (e.g., Labridae, Oplegnathidae, Xiphiorhynchidae), sharks dramatically decreases in diversity and quantity (a few Pristiophorus and odontaspidid teeth) and begin to predominate some sharks (Lamna) and teleosts with species characteristic of colder waters (e.g., gadiforms of the informal genus "Mesetaichthys") are recorded.
Long (1992c) and Case (1992) analyzed the ecology and diversity of the Eocene Seymour selachian fauna and concluded that the selachian fauna represents a cool-temperate fauna with different ecological components including tropical water immigrants (e.g., Pseudoginglymostoma, Stegostoma, Scoliodon).
While the diversity of the elasmobranch fauna in the Cucullaea I Allomember of the La Meseta Formation seems to be abundant and quite diverse, the teleost fishes appear to be low and poor. Teleost fishes are represented by gadiforms, clupeiforms, oplegnathids, siluriforms, perciforms, beryciforms, and chimaeriforms (see Table 2).
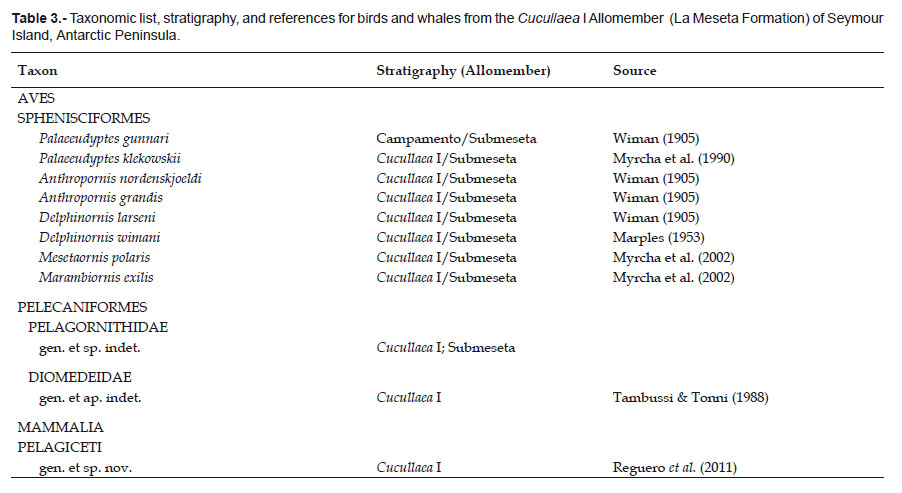
Penguins are by far the most dominant group of coastal/marine birds within this unit. Sphenisciformes are a group of flightless aquatic birds, distributed broadly throughout the Southern Hemisphere. Fossils indicate that stem penguins reached Antarctica by the late Paleocene (Tambussi et al. 2005), South America by the middle Eocene (Clarke et al. 2003), and Australia by the late Eocene (Simpson 1957; Jenkins 1974). Seymour Island Paleogene sequence (Cross Valley and La Meseta formations) has the longest and unique fossil record of basal sphenisciformes, with occurrences ranging from the late Paleocene (Cross Valley Formation) to late Eocene and constitutes the most complete Paleogene stratigraphic record of the group known in the world. The sphenisciform Palaeeudyptes is a taxon with biogeographic significance, is the most widespread Weddellian penguin genus in the Southern Hemisphere during the Eocene (Acosta Hospitaleche & Reguero 2010, 2011).
Stem penguin diversity peaks in the late Eocene by which time these birds are taxonomically diverse, geographically widespread, and abundant at many localities (e.g., Marples 1952; Myrcha et al. 2002; Tambussi et al. 2006).
The highest diversity of Weddellian sphenisciforms is documented in the Anthropornis nordenskjoeldi Biozone (Tambussi et al. 2006) defined within the Priabonian Submeseta Allomember, and this unit was deposited at 34.2 Ma based on 87/86Sr dates (Dingle & Lavelle 1998). Currently, 15 diagnosable sympatric species are recognized from this biozone, a total approaching modern global species-level diversity (19 species).
The first significant radiation of the Weddellian sphenisciforms took place in Ypresian beds of the Cucullaea I Allomember with 8 sympatric species (Table 3). The following eight species were identified in the Ypresian Cucullaea shell bed: Anthropornis nordenskjoeldi Wiman 1905, Anthropornis grandis (Wiman 1905), Palaeeudyptes gunnari (Wiman 1905), Palaeeudyptes klekowskii Myrcha, Tatur & Del Valle 1990, Delphinornis larseni Wiman 1905, Mesetaornis polaris Myrcha et al. 2002; Marambiornis exilis Myrcha et al. 2002, Archaeospheniscus wimani (Marples 1953), whereas the presence of D. arctowskii Myrcha et al. 2002, and D. gracilis Myrcha et al. 2002 are dubiously identified (Fig. 4).
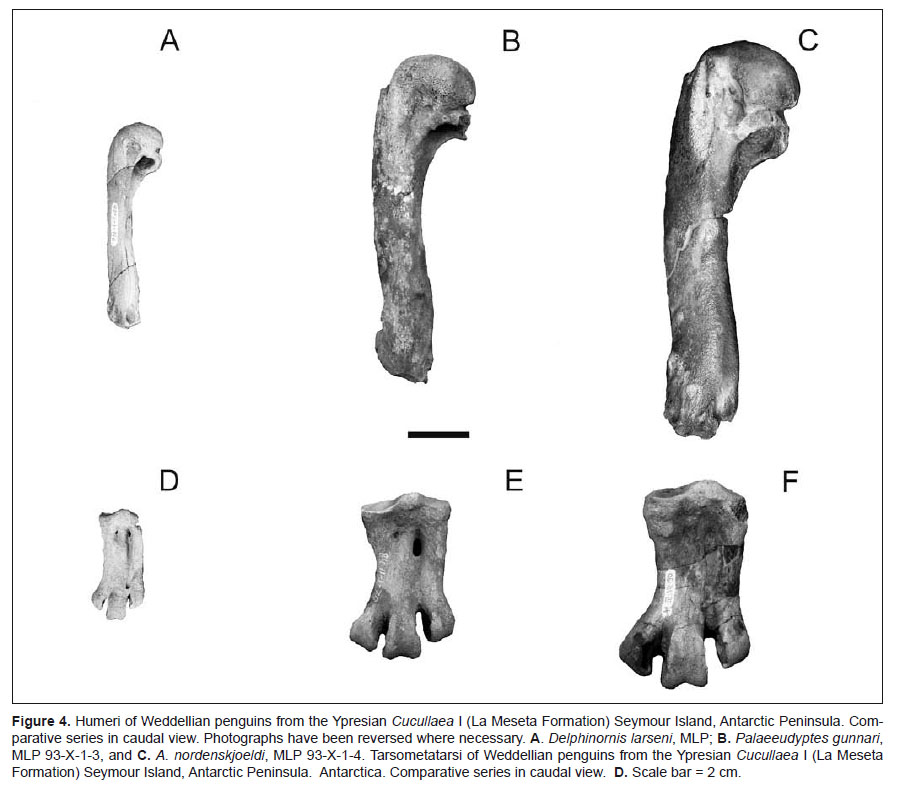
Although late Paleocene–middle Eocene sampling of the penguin record is not complete, stratigraphically calibrated, specific-level phylogenies (i.e. Ksepka & Clarke 2010; Clarke et al. 2010) indicate that an important radiation took place during this interval in Antarctica; at minimum, five clades of sphenisciforms diverged by the early Eocene (Psepka & Clarke 2010).
The underlying unit (Telm 3, late Early Eocene Campamento Allomember, 52 Ma) yielded small abundance of penguins with a low diversity, only one large species is recorded, Palaeeudyptes gunnari (Jadwiszczak 2006a, 2006b). The FAD (First Appearance Datum) of Palaeeudyptes gunnari is located within the Ypresian Campamento Allomember (La Meseta Formation) (Fig. 2).
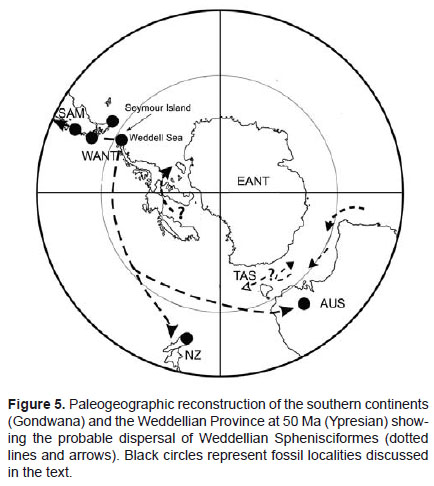
This Early Eocene sphenisciformes diversity supports several separate dispersals of giant penguins from Antarctica to lower latitudes: Australia (late Eocene, paleolatitude ~ 33°S, Jenkins 1974), New Zealand (late Eocene/early Oligocene, paleolatitude ~ 45°S, Simpson 1971), Argentina (middle Eocene, paleolatitude ~ 54°S, Clarke et al. 2003), Chile (middle to late Eocene, paleolatitude ~ 52°S, Sallaberry et al. 2010), and Peru (middle to late Eocene, paleolatitude ~14°S, Clarke et al. 2010) regions during greenhouse earth conditions (Fig. 5).
Whale remains occur sporadically throughout the La Meseta Formation (Cucullaea I and Submeseta allomembers). Archaeocetes from the La Meseta Formation of Seymour Island, Antarctica (Borsuk-Bialynicka 1988; Fostowicz-Frelik 2003), are in their majority based on unspecific postcranial material. Recently, Reguero et al. (2011) reported an incomplete jaw with teeth of a new basilosaurid archaeocete recovered from the Cucullaea shell bank. This discovery is significant as documented the earliest occurrence of a basilosaurid in Antarctica. The mandible bears five alveolus and two cheek teeth. The crown of one tooth, probably P2, is triangular and laterally compressed with multiple accessory denticles (3) and wear facet with a distinctive basal cingulid (Fig. 6).

This archaeocete represents the earliest record of whales in Antarctica (Ypresian, 49.5 Ma), and confirms the presence of this primitive group of whales (Basilosauridae) in the middle levels of La Meseta Formation. This specimen is the first inquestionable archaeocete described from Antarctica and is referred to a new taxon.
Discussion
During the Eocene, notwithstanding that some circulation could exist in Drake Passage area (Wrenn and Hart 1992), there was not a well developed Antarctic Circumpolar Current and consequently no author proposed the existence of the equivalent to Humboldt and Malvinas currents (Cione et al. 2007). During the Eocene, the Pacific coast would have been located at paleolatitude nearly equivalent present-day latitude, as there has been essentially no latitudinal translation of the area since the late Cretaceous. Cold-water upwelling along the western coast appears to have been in place by the Late Cretaceous or early Tertiary, and this "proto-Humboldt" current may have influenced low-latitude penguin diversity by cycling cold, nutrient-rich water into the ecosystem (Clarke et al. 2007).
The Eocene selachian fauna from Antarctica includes 25 species in 16 families; 24 taxa and 15 families come from the Cucullaea I Allomember of La Meseta Formation of Seymour Island. The majority of taxa belong to sharks while batoids are represented by only three taxa with a very uneven distribution in the sequence.
The Ypresian Cucullaea shell bed (Telm 4, basal horizon of the Cucullaea I Allomember) of La Meseta Formation is highly fossiliferous and yielded a remarkably diverse assemblage of plants, invertebrates and vertebrates that have provided the most detailed record of high-latitude southern Eocene organisms to date (e.g., Reguero et al. 2007). Nearly all phyla commonly preserved in the fossil record have been described from the unit (Schweitzer et al. 2005). Both continental and marine organisms indicate that the environment was completely different to that present in Antarctica today (Reguero & Marenssi 2010).
The shark fauna from the Ypresian Cucullaea shell bed (Telm 4, basal horizon of the Cucullaea I Allomember) of La Meseta Formation is one of the most diverse and abundant Early Eocene temperate neoselachian assemblages known from the Southern Hemisphere. So far, 22 selachian taxa within 14 families (including two batoids) have been reported from different localities on Seymour Island and from different levels of the Cucullaea I Allomember within the La Meseta Formation (Cione & Reguero, 1994, 1998; Long, 1992a, b, c; Welton & Zinsmeister, 1980). All these taxa have the oldest record in the Cucullaea I shell bed (Telm 4).
Today four penguin species: Aptenodytes forsteri (Emperor penguin), Pygoscelis adeliae (Adelie penguin), Pygoscelis antarctica (Chinstrap penguin), and Pygoscelis papua (Gentoo penguin) live and nest on Antarctica and sub-Antarctic islands. The phylogeny of the Sphenisciformes reveals that important radiations of small and large Weddellian penguins took place during the Late Paleocene/Early Eocene in Antarctica. The stratigraphical record of the sphenisciformes from Seymour Island is almost continuous from the Late Paleocene to Late Eocene and provides strong evidence on the evolution of this group in the Paleogene. One of the events clearly documented in this Paleogene sequence is the first radiation of this group that occurred in the Cucullaea shell bed (Telm 4) of the La Meseta Formation dated in a range of 49 to 52 Ma (Ypresian). The diversity of sphenisciformes increases from one in the Paleocene, one in the Valle de la Focas and Acantilados allomembers, probably two in the Campamento Allomember to nine species in the Cucullaea I Allomember.
They are broadly considered to be cool-adapted and the crown clade (Spheniscidae) seems to have the origin in association with the abrupt latest Eocene–Oligocene global cooling (~34 Ma) and to undergo a major radiation and range expansion to low latitudes only during later Neogene cooling (Baker et al. 2006). However, there is no fossil evidence to support a crown radiation in the Late Eocene concomitant with the initiation of the circum-Antarctic current, initial onset of Cenozoic global cooling, or at the proposed extinction of giant penguins.
The new gen. et sp. of archaeocete recorded in the Cucullaea shell bank represents the earliest record of whales fully aquatic (Ypresian, 49.5 Ma), and confirms the presence of this primitive group of whales (Basilosauridae) in West Antarctica.
Acknowledgments
We especially acknowledge the Instituto Antártico Argentino and Fuerza Aérea Argentina, which provided logistical support for our participation in the Antarctic fieldwork and the Agencia Nacional de Promoción Científica y Tecnológica (PICT, PICTO), Consejo Nacional de Investigaciones Científicas y Técnicas, and Universidad Nacional de La Plata for partial financial support for this project.
Literature cited
Acosta Hospitaleche C. & M. Reguero. 2010. First articulated skeleton of Palaeeudyptes gunnari from the late Eocene of Seymour Island (= Marambio) (Antarctica). Antarctic Science 22(3): 189-298. [ Links ]
Acosta Hospitaleche C. & M. Reguero. 2011. Evolution and biogeography of Paleogene Weddellian penguins (Aves: Sphenisciformes) of the James Ross Basin, Antarctic Peninsula. 11th International Symposium on Antarctic Earth Sciences (ISAES), Edinburgh, Scotland. [ Links ]
Borsuk-Bialynicka M. 1988. New remains of Archaeoceti from Paleogene of Antarctica. Polish Polar Research 9: 437–445. [ Links ]
Cappetta H. 1987. Chondrichthyes II. Mesozoic and Cenozoic Elamosbranchii. In: H.-P. Schultze ed. Handbook of Paleoichthyology Vol. 3B. Stuttgart and New York, Gustav Fischer Verlag. Pp. 1-193. [ Links ]
Case J.A. 1992. Evidence from fossil vertebrates for a rich Eocene Antarctic marine environment. Antarctic Research Series 56: 119-130. [ Links ]
Cione A.L., R. Del Valle, C. Rinaldi & E. Tonni. 1977. Nota preliminar sobre los pingiiinos y tiburones del Terciario inferior de la isla Vicecomodoro Marambio, Antártida. Contribuciones Cientificas del Instifuto Antártico Argentino 213: 3-21.
Cione A.L. & M. Reguero. 1994. New records of the sharks Isurus and Hexanchus from the Eocene of Seymour Island, Antarctica. Proceedings of the Geologist´s Association 105: 1-14. [ Links ]
Cione A.L. & M. Reguero. 1998. A middle Eocene basking shark (Lamniformes, Cetorhinidae) from Antarctica. Antarctic Science 10: 83-88. [ Links ]
Cione A.L., M.A. Reguero & C. Acosta Hospitaleche. 2007. Did the continent and sea have different temperatures in the northern Antarctic Peninsula during the middle Eocene? Revista de la Asociación Geológica Argentina 62(4): 586-596. [ Links ]
Clarke J.A., E.B. Olivero & P. Puerta. 2003. Description of the earliest fossil penguin from South America and first Paleogene vertebrate locality of Tierra del Fuego, Argentina. American Museum Novitates 3423: 1–18. [ Links ]
Clarke J.A., et al. 2007. Paleogene equatorial penguins challenge the proposed relationship between biogeography, diversity, and Cenozoic climate change. Proceedings of the National Academy of Sciences 104: 11545–11550. [ Links ]
Clarke, J.A., D.T. Ksepka, R. Salas-Gismondi, A.J. Altamirano, M. Shawkey, L. d’Alba, J. Vinther, T.J. de Vries, P. Baby. 2010. Fossil Evidence for Evolution of the Shape and Color of Penguin Feathers. Science 330: 954-957. [ Links ]
Compagno L.J.V 1988. Sharks of the order Carcharhiniformes. Princeton University, NJ, 486 pp. [ Links ]
Del Valle R.A., D.H. Elliot & D.I.M. Macdonald. 1992. Sedimentary basins on the east flank of the Antarctic Peninsula: proposed nomenclature. Antarctic Science 4:477-478. [ Links ]
Dingle R. & M. Lavelle. 1998. Antarctic Peninsula cryosphere: early Oligocene (c. 30 Ma) initiation and a revised glacial chronology. J. Geol. Soc. Lond. 155: 433-437. [ Links ]
Dutton A.L., K. Lohmann & W.J. Zinsmeister. 2002. Stable isotope and minor element proxies for Eocene climate of Seymour Island, Antarctica. Paleoceanography 17(2): 1–13. [ Links ]
Elliot D.H. & T.A. Trautman. 1982. Lower Tertiary strata on Seymour Island, Antarctic Peninsula, In: C. Craddock ed. Antarctic Geoscience, University of Wisconsin Press. Madison. Pp. 287-297. [ Links ]
Fostowicz-Frelik L. 2003. An enigmatic whale tooth from the Upper Eocene of Seymour Island, Antarctica. Polish Polar Research 24: 13–28. [ Links ]
Ivany L.C., S. Van Simaeys, E.W. Domack & S.D. Samson. 2006. Evidence for an earliest Oligocene ice sheet on the Antarctic Peninsula. Geology 34(5): 377–380. [ Links ]
Jadwiszczak P. 2006a. Eocene penguins of Seymour Island, Antarctica: taxonomy. Polish Polar Research 27: 3–62. [ Links ]
Jadwiszczak P. 2006b. Eocene penguins of Seymour Island, Antarctica: the earliest record, taxonomic problems and some evolutionary considerations. Polish Polar Research 27: 287–302. [ Links ]
Jenkins R.J.F. 1974. A new giant penguin from the Eocene of Australia. Palaeontology 17: 291–310. [ Links ]
Jianjun L. & Z. Shuonan. 1994. New Materials of bird ichnites from Fildes Peninsula, King George Island of Antarctica and their biogeographic significance. In: Y. Shen ed. Stratigraphy and palaeontology of Fildes Peninsula, King George Island, Antarctica. State Antarctic Commitee, Monograph 3. Pp. 239-249. Science Press, Beijin. [ Links ]
Kriwet J. 2005. Additions to the Eocene selachian fauna of Antarctica with comments on Antarctic selachian diversity. Journal of Vertebrate Paleontology 25(1):1-7. [ Links ]
Ksepka, D. T. & J. A. Clarke. 2010. The basal penguin (Aves: Sphenisciformes)Perudyptes devriesi and a phylogenetic evaluation of the penguin fossil record. Bulletin of the American Museum of Natural History 337:1–77. [ Links ]
Long D.J. 1992a. The shark fauna from La Meseta Formation (Eocene), Seymour Island, Antarctic Peninsula. Journal of Vertebrate Paleontology 12:1-32. [ Links ]
Long D.J. 1992b. An Eocene wrasse (Perciformes; Labridae) from Seymour Island. Antarctic Science 4:235-237. [ Links ]
Long D.J. 1992c. Paleoecology of Eocene Antarctic sharks, In: J.P. Kennett and D.A. Warnke eds. The Antarctic paleoenvironment: a perspective on global change, Antarctic Research Series 56. Pp. 131-139. [ Links ]
Long D.J. & J.D. Stilwell. 2000. Fish remains from the Eocene of Mount Discovery, East Antarctica. Antarctic Research Series 76: 349-353. [ Links ]
Marenssi S.A., S.N. Santillana & C.A. Rinaldi. 1998a. Paleoambientes sedimentarios de la Aloformación La Meseta (Eoceno), Isla Marambio (Seymour), Antártida. Instituto Antártico Argentino, Contribución 464: 1-51. [ Links ]
Marenssi S.A., S.N. Santillana & C.A. Rinaldi. 1998b. Stratigraphy of the La Meseta Formation (Eocene), Marambio (Seymour) Island, Antarctica. In: S. Casadio ed. Paleógeno de América del Sur y de la Península Antártica, Asociación Paleontológica Argentina, Publicación Especial 5. Pp. 137-146, Buenos Aires. [ Links ]
Marples B.J. 1952. Early Tertiary penguins of New Zealand. New Zealand Geological Survey, Palaeontological Bulletin 20: 1–66. [ Links ]
Montes M., F. Nozal, S. Santillana, F. Tortosa, E. Beamud & S. Marenssi. 2010. Integrate stratigraphy of the Upper Paleocene-Eocene strata of Marambio (Seymour) Island, Antarctic Peninsula. XXXI SCAR, Open Science Conference, Buenos Aires, Argentina. [ Links ]
Myrcha A., A. Tatur & R. del Valle. 1990. A new species of fossil penguin from Seymour Island, West Antarctica. Alcheringa 14: 195–205. [ Links ]
Myrcha A., P. Jadwiszczak, C. Tambussi, J. Noriega, A. Gazdzicki, A. Tatur & R. del Valle. 2002. Taxonomic revision of Eocene Antarctic penguins based on tarsometatarsal morphology. Polish Polar Research 23: 5–46. [ Links ]
Reguero M.A., S.A. Marenssi & S.N. Santillana. 2002. Antarctic Peninsula and Patagonia Paleogene terrestrial environments: biotic and biogeographic relationships. Palaeogeography, Palaeoclimatology, Palaeoecology 179: 189-210. [ Links ]
Reguero M.A. & Z. Gasparini. 2007. Late Cretaceous–Early Tertiary marine and terrestrial vertebrates from James Ross Basin, Antarctic Peninsula: a review. In: J. Rabassa and M.L. Borla eds. Antarctic Peninsula & Tierra del Fuego: 100 years of Swedish-Argentine scientific cooperation at the end of the world, London, Taylor & Francis. Pp. 55-76. [ Links ]
Reguero M.A. & S.A. Marenssi. 2010. Paleogene climatic and biotic events in the terrestrial record of the Antarctic Peninsula: An overview. In: R. Madden, A.A. Carlini, M.G. Vucetich and R. Kay eds. The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia. Cambridge University Press. Pp. 383-397. [ Links ]
Reguero M.A., C. Tambussi, T. Mörs, M. Buono, S. Marenssi, S. Santillana. 2011. Vertebrates from the basal horizons (Ypresian to Lutetian) of the Cucullaea I Allomember, La Meseta formation, Seymour (Marambio) Island, Antarctica. 11th International Symposium on Antarctic Earth Sciences (ISAES), Edinburgh, Scotland. [ Links ]
Sadler P. 1988. Geometry and stratification of uppermost Cretaceous and Paleogene units on Seymour Island, northern Antarctic Peninsula. In: R.M. Feldmann and M.O. Woodburne eds. Geology and Paleontology of Seymour Island, Antarctic Peninsula, Geological Society of America, Memoir 169. Pp. 303-320. Boulder. [ Links ]
Sallaberry M.A., R.E. Yury-Yáñez, R.A. Otero, S. Soto-Acuña & T. Torres G. 2010. Eocene birds from the western margin of southernmost South America. Journal of Paleontology 84(6): 1061–1070 [ Links ]
Schweitzer C., R. Feldmann, S. Marenssi & D.A. Waugh. 2005. Remarkably preserved annelid worms from the La Meseta Formation (Eocene), Seymour Island, Antarctica. Palaeontology 48: 1-13. [ Links ]
Simpson G.G. 1946. Fossil penguins. Bulletin of the American Museum of Natural History 87(1): 1–99. [ Links ]
Simpson G.G. 1957. Australian fossil penguins, with remarks on penguin evolution and distribution. Records of the South Australian Museum 13: 51–70. [ Links ]
Simpson G.G. 1971. Review of fossil penguins from Seymour Island. Proceedings of the Royal Society of London B Biological Sciences 178: 357–387. [ Links ]
Tambussi P. & E.P. Tonni. 1988. Un Diomedeidae (Aves, Procellariformes)del Eoceno tardio de la AnCtida. In: J.C. Quiroga an A.L. Cione eds. 5th Jornadas Argentinas de Paleontología de Vertebrados, Abstracts, La Plata, Universidad de La Plata, p. 4. [ Links ]
Tambussi C.P., C.I. Acosta Hospitaleche, M.A. Reguero & S.A. Marenssi. 2006. Late Eocene penguins from West Antarctica: systematics and biostratigraphy. In: J.E. Francis, D. Pirrie and J.A. Crame eds. Cretaceous-Tertiary high-latitude palaeoenvironments, James Ross Basin, Antarctica. Geological Society of London, Special Publications 258. London. Pp. 145–161. [ Links ]
Welton B.J. & W.J. Zinsmeister. 1980. Eocene neoselachians from La Meseta Formation, Seymour Island, Antarctic Peninsula. Contributions in Science, Museum Los Angeles County 329: 1-10. [ Links ]
Wiman C.1905. Über die alttertiaren Vertebraten der Seymourinsel. Wiss. Ergebnisse der Schwedishen Süspolar-Expedition 1901–1903 3: II–6. [ Links ]
Wrenn J.H. & G.F. Hart. 1988. Paleogene dinoflagellates cyst biostratigraphy of Seymour Island, Antarctica. In: R.M. Feldmann and M.O. Woodburne eds. Geology and Paleontology of Seymour Island, Antarctic Peninsula, Geological Society of America, Memoir 169. Boulder. Pp. 321-447. [ Links ]
Zinsmeister W.J. 1979. Biogeographic significance of the late Mesozoic and early Tertiary molluscan faunas of Seymour Island (Antarctic Peninsula) to the final breakup of Gondwanaland. In: J. Gray and A. Boucot eds. Historical biogeography,plate tectonics and the changing environment, Oregon State University. Pp. 349-355. [ Links ]
Zinsmeister W.J. 1982. Late Cretaceous-Early Tertiary molluscan biogeography of southern Circum-Pacific. Journal of Paleontology 56: 84-102. [ Links ]
Email: regui@fcnym.unlp.ed.ar
Presentado: 01/04/2012
Aceptado: 20/11/2012
Publicado online: 15/01/2013
