










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. vol.21 no.3 Lima dic. 2014
http://dx.doi.org/10.15381/rpb.v21i3.10896
TRABAJOS ORIGINALES
Comunidad de avispas Ophioninae (Hymenoptera: Ichneumonidae) en el bosque nublado Monteseco, Cajamarca, Perú
Ophioninae (Hymenoptera: Ichneumonidae) wasp community in the cloudy forest Monteseco, Cajamarca, Peru
Evelyn Sánchez1,2, Mabel Alvarado3,1 y Juan Grados1
1 Departamento de Entomología, Museo de Historia Natural, Av. Arenales 1256 Jesús María, Lima 14, Perú.
2 Departamento de Entomologia, Universidade Federal de Viçosa, Av. P. H. Rolfs S/N, Campus Universitário, Viçosa, Minas Gerais, 36570-900, Brasil.
3 Division of Entomology, Natural History Museum, and Department of Ecology & Evolutionary Biology, 1501 Crestline Drive – Suite 140, University of Kansas, Lawrence, Kansas, 66045, USA.
Email Evelyn Sánchez: eyss13@hotmail.com
Email Mabel Alvarado: mag2199@hotmail.com
Email Juan Grados: gradosjuan@hotmail.com
Resumen
Se describe la composición de especies de la subfamilia Ophioninae (Hymenoptera: Ichneumonidae) a lo largo de una gradiente altitudinal en el bosque nublado Monteseco, Cajamarca, Perú; colectados en los años 2009 y 2010. Se encontraron 18 especies en tres géneros de Ophioninae: Alophophion, Enicospilus y Ophion. Cinco especies son registradas por primera vez para Perú: Ophion polyhymniae Gauld, 1988, Enicospilus cubensis (Norton, 1863), E. guatemalensis (Cameron, 1886), E. cressoni Hooker, 1912 y E. mexicanus (Cresson, 1874). La composición de la subfamilia Ophioninae varía según la elevación. La mayor riqueza de Ophioninae se encontró a 2150 m (S=11) y la menor a 3116 m (S=3). Enicospilus es más diverso en zonas de baja a mediana altitud, Ophion es más diverso en zonas de mediana a elevada altitud y Alophophion ocurre predominantemente a elevada altitud.
Palabras claves: Ophioninae; Neotrópico; especies; diversidad; altitud.
Abstract
We describe the species composition of the subfamily Ophioninae (Hymenoptera: Ichneumonidae) along an altitudinal gradient in the cloudy forest Monteseco, Cajamarca, Peru collected in 2009 and 2010. Eighteen species were recorded in three genera of Ophioninae: Alophophion, Enicospilus y Ophion. Five species are recorded for the first time in Peru: Ophion polyhymniae Gauld, 1988; Enicospilus cubensis (Norton, 1863); E. guatemalensis (Cameron, 1886); E. cressoni Hooker, 1912 y E. mexicanus (Cresson, 1874). Subfamily composition varies with the elevation. The highest species richness (S=11) was found at 2150 m and the lowest (S=3) at 3116 m. Enicospilus is more diverse from low to mid elevation, Ophion from mid to high elevation and Alophophion occurs predominantly at high elevation.
Keywords: Ophioninae; Neotropics; species; diversity; elevation.
Introducción
La subfamilia Ophioninae presenta 33 géneros y aproximadamente 1025 especies en todo el mundo (Yu et al. 2005). En la región Neotropical se encuentran 12 géneros (Gauld 1988) y 240 especies (Fernández-Triana 2005, Yu et al. 2005). En Perú se han registrado 49 especies (Rodríguez-Berrío et al. 2009, Alvarado 2014) distribuidas en siete géneros (Rodríguez-Berrío et al. 2009, Alvarado et al. 2010), de los cuales los géneros Janzophion, Simophion y Ophiogastrella presentan tan solo la ocurrencia del género (Alvarado et al. 2010). El género Alophophion está representado por 17 especies (Alvarado 2014), Thyreodon por cuatro especies, Enicospilus por 26 especies y Ophion únicamente por O. flavidus Brullé, 1846 (Yu et al. 2005, Rodríguez-Berrío et al. 2009).
Los Ophioninae son generalmente grandes (la longitud del ala anterior fluctúa entre 14 y 22 mm), esbeltos y de color castaño-amarillentos. Son particularmente distintivos por las características de su venación alar. El ala anterior carece de areolet y de vena 2rs-m; vena 3rs-m presente, ubicada muy distalmente de 2m-cu y, la celda discosubmarginal frecuentemente con un área anterior glabra o de pocos pelos denominada fenestra (Gauld & Lanfranco 1987, Gauld 1988, Lee & Kim 2002).
La mayoría de Ophioninae son endoparásitos koinobiontes y solitarios, principalmente de larvas de Lepidoptera de las familias Noctuidae, Lasiocampidae, Lymantriidae, Saturniidae, Geometridae, Arctiidae y Sphingidae (Carlson 1979). Varias especies de Ophioninae son parasitoides comunes de plagas de lepidópteros en bosques y agroecosistemas (Rohlfs & Mack 1983, Fritz et al. 1986) por lo que el mejor entendimiento de estas avispas facilitaría utilizarlas como controladores biológicos (Gauld 1988). Además, los Ophioninae pueden ser utilizados en estudios biogeográficos y ecológicos ya que a diferencia de muchos Ichneumonidae, gran número de especímenes pueden ser colectados efectivamente con trampas de luz de vapor de mercurio (Gauld & Lanfranco 1987) debido a que estas avispas son generalmente activas durante la noche (Gauld 1988).
En vista de la importancia ecológica y taxonómica de esta subfamilia, este trabajo busca caracterizar la composición de especies de Ophioninae presentes en el bosque nublado Monteseco, departamento de Cajamarca, Perú.
Materiales y métodos
El estudio se realizó en el bosque nublado Monteseco, uno de los 23 bosques relictos de la Vertiente Occidental de los Andes de Perú (Weigend et al. 2006), caracterizado por la presencia de bosques nublados montanos y pajonales de altura o jalca (SERNANP 2011). Por encima de los 1000 m es posible encontrar abundante vegetación, siendo la zona más densa entre los 1500 m - 2000 m. En este rango se encuentran acantilados cubiertos por bosques húmedos, mantenidos por las fuertes lluvias estacionales y por la nubosidad continua que cubren la parte occidental por varios meses del año (Sagástegui & Dillon, 1991).
Las colectas se llevaron a cabo en los años 2009 y 2010. En el 2009 se realizaron dos evaluaciones, la primera en el mes de abril y la segunda en octubre. Los puntos altitudinales fueron los siguientes: 1195 m (en este punto altitudinal no se colectó en la primera evaluación), 1615 m, 2150 m y 2841 m. En el 2010 solo se hizo una tercera evaluación en el mes de mayo, colectándose en los mismos puntos altitudinales del 2009, añadiendo el punto a 3116 m (Tabla 1).
El material en estudio fue colectado con trampas de luz. Como fuente luminosa se usó una lámpara de luz mixta de 250 voltios y como fuente de poder se utilizaron generadores portátiles. Todas las colectas se realizaron durante la fase de luna nueva. El esfuerzo de muestreo fue de seis horas por noche, desde las 18:00 hasta las 00:00 horas. En cada elevación se colectó por tres noches consecutivas.
Las imágenes de los especímenes son montajes compuestos de fotografías tomados con una cámara digital D1x Nikon, conectada a un lente microscópico de larga distancia Infinity K-2 o con la cámara digital Canon EOS T3, acoplada a un adaptador Clearshot 600 Digital. Las fotografías fueron integradas en una sola imagen con el programa Combine ZP (Hadley, 2006). Se editaron con el programa Adobe Photoshop CS5 y las láminas fueron preparadas con el programa Adobe Illustrator CS5.
Para la determinación hasta el nivel de género se siguieron las claves y descripciones propuestas por Gauld y Lanfranco (1987), Gauld (1985) y Townes (1969); para la identificación a nivel de especie, las claves propuestas por Gauld (1988). La identificación de algunas especies fue confirmada mediante la comparación de los especímenes con el material examinado por Gauld, depositados en el Museo de Historia Natural de Londres, Inglaterra (BMNH).
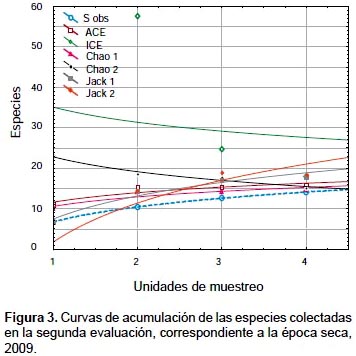
Para analizar el esfuerzo de muestreo realizado se utilizaron los siguientes estimadores de riqueza no paramétricos: Chao 1, Chao 2, Jackknife de primer orden, Jackknife de segundo orden, ACE (Abundance-based Coverage Estimator) y ICE (Incidence-based Coverage Estimator), todos obtenidos mediante el programa EstimateS (Version 8.2.0). Debido a que el esfuerzo de muestreo realizado en los diferentes puntos altitudinales fue heterogéneo, solamente se emplearon estos estimadores para la segunda y tercera evaluación. Además, ya que solo en la tercera evaluación se colectó a 3116 m, las estimaciones de riqueza en ese punto altitudinal no fueron consideradas.
Los valores de estimadores de riqueza se exportaron al programa STATISTICA 7.0 para la construcción de las curvas de acumulación, de acuerdo a lo propuesto por Jiménez-Valverde & Hortal (2003). Las curvas fueron ajustadas al modelo de Clench, según Soberón & Llorente (2003).
Resultados y discusión
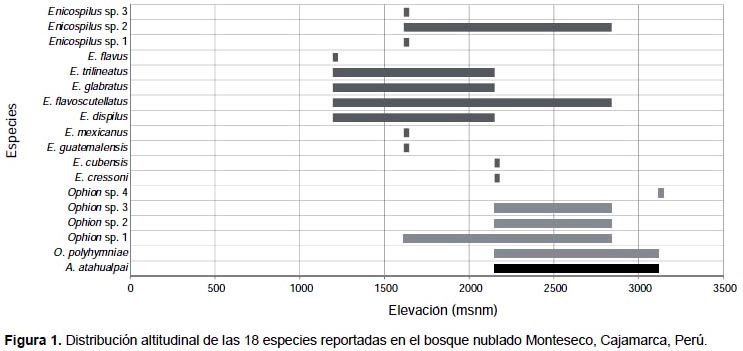
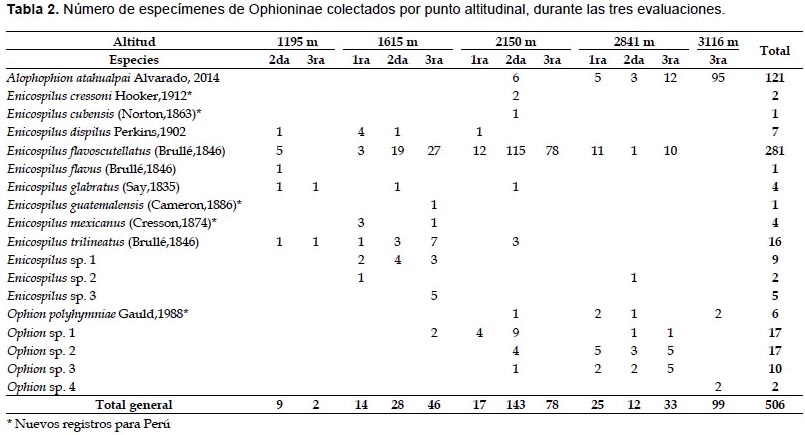
Se registraron tres géneros pertenecientes a la subfamilia Ophioninae: Alophophion, Enicospilus y Ophion. Se registraron cinco especies pertenecientes al género Ophion, 12 especies pertenecientes al género Enicospilus y solo una especie perteneciente al género Alophophion (Fig. 1, Tabla 2). Del total de especies registradas (Tabla 2) en el presente trabajo, seis han sido registradas previamente para Perú (Rodríguez-Berrío et al. 2009, Alvarado 2014); cinco son nuevos registros para Perú (O. polyhymniae, E. cubensis, E. cressoni, E. guatemalensis y E. mexicanus) (Fig. 5B-F) y siete taxa han quedado como morfoespecies: cuatro pertenecientes al género Ophion y tres a Enicospilus.
Los taxa que no fueron identificados al nivel de especie, podrían tratarse de especies nuevas para la ciencia y no pudieron ser identificadas con la información actual, debido a que la principal fuente de consulta fueron las claves propuestas por Gauld (1988) que buscan identificar la fauna de América Central y no la de América del Sur. A pesar de eso, Gauld (1988) incluyó los datos de ocurrencia de las especies en otros países tales como Perú, siendo ésta la principal fuente de información para la región. La fauna de América del Sur ha sido poco estudiada, pocas especies han sido descritas o las descripciones de las existentes son insuficientes para discernir entre ellas, tal como fue registrado por Alvarado (2014) en la revisión de Alophophion. Los géneros Ophion y Enicospilus no cuentan con revisiones en América del Sur lo que limita la identificación hasta especie.
La composición de la subfamilia Ophioninae varió según la elevación (Fig. 1 y 2, Tabla 2). La menor abundancia se obtuvo a 1195 m (N=11), donde solo se registraron especies de Enicospilus. A 1615 m se registraron 10 especies: nueve especies pertenecientes al género Enicospilus y solo una especie a Ophion. A 2150 m se encontraron 11 especies: seis pertenecientes al género Enicospilus, cuatro a Ophion y una de Alophophion – A. atahualpai Alvarado, 2014 (Fig. 5A). En esta elevación se pudo evidenciar la marcada dominancia de E. flavoscutellatus (Brullé, 1846) (N=205), representando el 86.13% de los individuos colectados en ese punto altitudinal. A 2841 m la riqueza fue menor con respecto al anterior punto altitudinal, se colectaron siete especies: A. atahualpai, dos especies de Enicospilus y cuatro de Ophion; a esta altitud se pudo evidenciar que el número de especímenes de E. flavoscutellatus (N=22) fue menor al registrado a 2150 m. Solo a 2150 m y 2841 m se colectaron los tres géneros de Ophioninae. A 3116 m se colectaron tres especies: A. atahualpai y dos especies de Ophion; siendo considerablemente más abundante la especie A. atahualpai (N=95), representando el 95.96% de la comunidad en ese punto altitudinal.
El género Enicospilus estuvo presente desde 1195 m hasta 2841 m y ausente a 3116 m (Fig. 1, Tabla 2). Estos resultados son similares a los registrados en Costa Rica por Gauld y Lanfranco (1987), donde ninguna especie de este género fue colectada a más de 3000 m de altitud; ellos también mencionan que este género presenta alta diversidad entre 300 m y 1500 m en las zonas húmedas del Neotrópico. En el presente estudio no se realizaron colectas en puntos altitudinales menores a 1195 m, sin embargo, el mayor número de especies (S=9) se registró a 1615 m (Tabla 2), siendo ésta una elevación ligeramente mayor a la sugerida por Gauld y Lanfranco (1987).
En el presente estudio la mayoría de especies fueron encontradas sobre los 2100 m, registrándose 11 especies a 2150 m, siete especies a 2841 m y tres especies a 3116 m (Fig. 1); a diferencia de lo encontrado en América Central por Gauld (1988), donde solo se registró a O. polyhymniae habitando sobre los 2700 m. Además, el presente estudio reporta para el género Ophion cuatro especies a 2150 m, cuatro especies a 2841 m y dos especies a 3116 m (Fig. 1, Tabla 2), evidenciándose una mayor diversidad de este género a puntos altitudinales más elevados que los encontrados en América Central por Gauld (1988).
Como se puede observar en la Figura 1, existen especies que fueron colectadas en solo un punto altitudinal, mientras que otras fueron registradas en un amplio rango de elevación. Enicospilus flavoscutellatus es la especie que presentó un mayor rango distribución a lo largo de la gradiente altitudinal, habitando desde 1195 m hasta 2841 m, presentó además un pico de abundancia a 2150 m (Fig. 2). La abundancia de esta especie decrece mientras aumenta la altitud, resultados similares fueron reportados por Gauld (1988). Nuestro estudio no encontró diferencias en cuanto a la distribución altitudinal de las especies (Tabla 2) a diferencia de lo reportado por Gauld (1988).
Alophophion es el único género de Ophioninae que está restringido para América del Sur (Gauld & Lanfranco 1987, Gauld 1988). Gauld y Lanfranco (1987) sugirieron que este género tendría como límite de distribución septentrional alrededor de los 25°S; sin embargo en el presente estudio fue colectado en los 6°S (Tabla 1). Según Alvarado (2014), el género Alophophion ocurre desde el sur de Chile y Argentina hasta Ecuador. La especie registrada en el presente estudio también ocurre en Ecuador, siendo la especie con distribución más septentrional del género (Alvarado 2014). Esta especie fue colectada desde los 2150 m hasta los 3116 m pero es considerablemente más abundante a 3116 m (Tabla 2, Fig. 2).
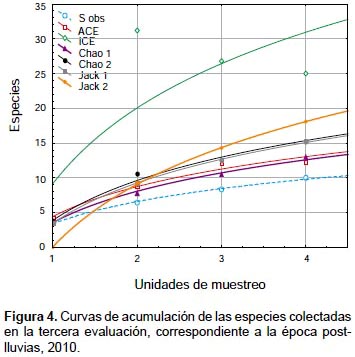

Las curvas de acumulación sugieren que durante la segunda evaluación (Fig. 3), correspondiente a la época seca, se colectó entre 75.19% (ACE) y 79.15% (Chao 1) de la riqueza estimada. En la tercera evaluación (Figura 4), correspondiente a la época de post-lluvias, se colectó entre el 21.92% (Chao 1) y 39.82% (ACE) de la riqueza estimada, lo que sugeriría que la estacionalidad estaría influenciando la ocurrencia de las especies de Ophioninae. Además, se evidencia para las dos evaluaciones que los mejores estimadores de riqueza fueron ACE y Chao 1, los cuales se basan en la abundancia de las especies colectadas.
Es de esperar que la composición de especies sea considerablemente distinta en ambientes más áridos de la vertiente occidental de los Andes. Por ejemplo, el género Enicospilus presenta pocas especies en ambientes con marcada estación seca y desiertos (Gauld 1985) mientras que Alophophion es muy diverso en ese tipo de ambientes (Alvarado 2014). No existen estudios en las zonas áridas de la vertiente occidental de los Andes, por lo que no se puede comparar con la comunidad de Ophioninae de los bosques húmedos de montaña como el bosque nublado Monteseco.
La distribución de los géneros Enicospilus y Ophion a lo largo de la gradiente altitudinal en el bosque nublado Monteseco es consistente con lo encontrado por Gauld (1988) y Gauld y Lanfranco (1987) para bosques húmedos de montaña, donde el género Enicospilus es más diverso en zonas de baja a mediana altitud y Ophion es más diverso en zonas de mediana a elevada altitud.
Agradecimientos
Queremos agradecer a Conservation International (CI) y a la Asociación Peruana para la Conservación de la Naturaleza (APECO) por la subvención a Juan Grados para llevar a cabo la investigación sobre Lepidópteros Neotropicales en el bosque nublado Monteseco. Adicionalmente, queremos agradecer a dos revisores anónimos por sus observaciones.
Literatura citada
Alvarado, M. 2014. Revision of the South American wasp genus Alophophion Cushman, 1947 (Hymenoptera: Ichneumonidae: Ophioninae). Revista Peruana de Biología, 21(1), 003–060. http://dx.doi.org/10.15381/rpb.v21i1.8245.
Alvarado M., A. Rodríguez-Berrío & S. Bordera. 2010. Colección de Ichneumonidae (Hymenoptera) del Museo de Entomología "Klaus Raven B." y del Museo de Historia Natural de la UNMSM, Perú. En: Toledo, V. H. Corona, A. M., Flores, A. P., Tovar, E, Coronado, J. M. Ruíz-Cancino, E (Editores). II Taller Internacional de Recursos Naturales. Red de Cuerpos Académicos, México. 107 pp.
Carlson R. 1979. Family Ichneumonidae. In: Krombein, K., Hurd, P., Smith, D. & Burks, B (Ed), Catalog of Hymenoptera in America north of Mexico, Volume 1, Smithsonian Institution, Washington, D.C.1198 pp.
Colwell R.K. & J.A. Coddington. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society, London, Series B, 345: 101-118.
Fernández-Triana J.L. 2005. The taxonomy and biogeography of Cuban Ophioninae (Hymenoptera: Ichneumonidae). Zootaxa, 1007: 1-60.
Fritz G.N., A.P. Frater, J.C. Owens, E.W. Huddleston & D.B. Richman. 1986. Parasitoids of Hemileuca oliviae (Lepidoptera: Saturniidae) in Chihuahua, Mexico. Annals of the Entomological Society of America, 79: 686-690.
Gauld I.D. 1985. The phylogeny, classification and evolution of parasitic wasps of the subfamily Ophioninae (Ichneumonidae). British Museum (Natural History).
Gauld I.D. 1987. Some factors affecting the composition of tropical ichneumonid faunas. Biological Journal of the Linnean Society, 30: 299-312. http://dx.doi.org/10.1111/j.1095-8312.1987.tb00304.x.
Gauld I.D. 1988. A survey of the Ophioninae (Hymenoptera: Ichneumonidae) of tropical Mesoamerica with special reference to the fauna of Costa Rica. Bull. Brit. Mus. (Natur. Hist.) Entomol. 57: 1-309.
Gauld I.D. & D. Lanfranco. 1987. Los géneros de Ophioninae de Centro y Sudamérica. Revista Biología Tropical, 35: 257-267.
Food and Drug Administrations (FDA). 2001. Fish and Fishery Products Hazards and Controls Guidance. Third Edition June 2001. <http://www.cfsan.fda.gov/~comm/haccp4.html> (acceso 24/12/07).
Hadley A. 2006. (en línea). Combine ZP, Image Stacking Software. <http://www.hadleyweb.pwp.blueyonder.co.uk/CZP/News.htm>. Acceso 25/01/2013.
Jiménez-Valverde A. & J. Hortal. 2003. Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología, 8: 151-161.
Lee J.W. & K.B. Kim. 2002. Taxonomical Review of the Subfamily Ophioninae (Hymenoptera: Ichneumonidae) I. Genus Stauropoctonus Brauns. Korean journal of entomology, 32: 81-86. http://dx.doi.org/10.1111/j.1748-5967.2002.tb00015.x.
Magurran A. 1988. Ecological diversity and its measurement. Princeton University Press, New Jersey, 179 pp. http://dx.doi.org/10.1007/978-94-015-7358-0.
Rodríguez-Berrío A., S. Bordera, I. Sääksjärvi. 2009. Checklist of Peruvian Ichneumonidae (Insecta, Hymenoptera). Zootaxa, 2303: 1-44.
Rohlfs W.M. & T.P. Mack. 1983. Effect of parasitization by Ophion flavidus Brullé (Hymenoptera: Ichneumonidae) on consumption and utilization of a pinto bean diet by fall armyworm (Lepidoptera: Noctuidae). Environmental Entomology, 12: 1257-1259.
Sääksjärvi I.E., S. Haataja, S. Neuvonen, I.D. Gauld, R. Jussila, J. Salo, A.M. Burgos. 2004. High local species richness of parasitic wasps (Hymenoptera: Ichneumonidae; Pimplinae and Rhyssinae) from the lowland rainforests of Peruvian Amazonia. Ecological Entomology, 29: 735–743. http://dx.doi.org/10.1111/j.0307-6946.2004.00656.x.
Sagástegui A. & Dillon M. 1991. Inventario preliminar de la Flora del Bosque Monteseco. Arnaldoa, 1(1):35-52.
Servicio Nacional de Áreas Naturales protegidas por el Estado (SERNANP). (en línea). Decreto Supremo de categorización definitiva de la Zona Reservada Udima en Refugio de Vida Silvestre "Bosques Nublados de Udima" Nº 020-2011-MINAM, Lima. <http://www.sernanp.gob.pe/sernanp/contenido.jsp?ID=77&C=ANP>. Acceso: 07/10/2014.
Soberón J. & J. Llorente. 1993. The use of species accumulation functions for the prediction of species richness. Conservation biology, 7: 480-488. http://dx.doi.org/10.1111/10.1046/j.1523-1739.1993.07030480.x.
Townes H. 1969. The genera of Ichneumonidae, Part 1. Memoirs of the American Entomological Institute, 11: 1-300.
Weigend M., N. Dostert & E. Rodríguez. 2006. Bosque relictos de los andes peruanos: Perpectivas económicas. Botánica económica de los Andes Centrales: 130-145.
Yu D.S., K. Van Achterberg & K. Horstmann. 2005. World Ichneumonoidea 2004. Taxonomy, biology, morphology and distribution. CD/DVD, Taxapad, Vancouver, Canada. (http://www.taxapad.com).
Presentado: 05/02/2014
Aceptado: 06/10/2014
Publicado online: 30/12/2014

