










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista Peruana de Biología
On-line version ISSN 1727-9933
Rev. peru biol. vol.21 no.3 Lima Dec. 2014
http://dx.doi.org/10.15381/rpb.v21i3.10903
NOTA CIENTÍFICA
Caracterización molecular de papas nativas (Solanum spp.) del distrito de Chungui, Ayacucho, mediante AFLP
Molecular characterization of native potato (Solanum spp.) Chungui, Ayacucho, using AFLP
Juan C. Gonzales Mamani y Gilmar Peña Rojas
Laboratorio de Biología Celular y Molecular – Facultad de Ciencias Biológicas, Universidad Nacional de San Cristóbal de Huamanga, Ayacucho – Perú.
Email Gilmar Peña: gilmar_p@yahoo.com
Email Juan Gonzales: juancarlos.gm20@gmail.com
Resumen
Con el objetivo de evaluar la diversidad genética de Solanum spp. del distrito de Chungui, La Mar, Ayacucho, se colectaron 25 morfotipos de papas nativas los cuales fueron introducidos y micropropagados en el medio de cultivo Murashigue Skoog (1962). La extracción de ADN se procedió utilizando el método CTAB modificado a partir de hojas de 3 semanas cultivo, se logró buena calidad y cantidad de ADN para emplear el AFLP. La digestión enzimática del ADN se realizó utilizando EcoRI y MseI. Se emplearon tres combinaciones de primers AFLP con tres nucleótidos selectivos, originándose un total de 85 bandas claramente diferenciables, de las cuales 63 fueron polimórficas. La combinación E37 – M50 fue la más informativa obteniendo un índice de contenido polimórfico de 0.43. La lectura de la presencia o ausencia de bandas polimórficas de los morfotipos evaluados empleando el coeficiente de similitud Simple Matching y el análisis de agrupamiento según el algoritmo UPGMA originó un dendograma con un índice de correlación cofenética de r= 0.7. El dendograma con un coeficiente de similitud de 0.64 agrupó a los morfotipos de papas nativas en 4 grupos genéticos y no se encontró morfotipos duplicados a pesar de tener algunas características morfológicas muy semejantes. Estos resultados demuestran el alto poder informativo de los marcadores AFLP en el análisis de la diversidad genética de papas nativas.
Palabras clave: AFLP; diversidad genética; papas nativas; Solanum spp.; caracterización.
Abstract
Genetic diversity of 25 morphotypes of native potatoes Solanum spp. from Chungui (La Mar, Ayacucho) were assess. Morphotypes collected were micropropagated in Murashigue Skoog medium (1962). DNA extraction proceeded using the CTAB method modified from 3 weeks leaves crop, good quality and quantity of DNA was able to use the AFLP. Enzymatic digestion of the DNA was performed using EcoRI and MseI. Three combinations of AFLP primers with three selective nucleotides were used, resulting in a total of 85 clearly discernable bands, of which 63 were polymorphic. The combination E37 – M50 showed the most informative polymorphic index content of 0.43. The presence/absence of polymorphic bands was evaluated using the Simple Matching coefficient similarity and clustering analysis using the UPGMA. The dendrogram produced had a cophenetic correlation coefficient r= 0.7. At the level 0.64 of Simple Matching coefficient similarity, the dendrogram grouped the morphotypes of native potatoes in 4 genetic groups, it not found duplicated morphotypes, despite having some morphotypes very similar. Our results would be showing the highly informative power of AFLP markers for the analysis of genetic diversity of native potatoes.
Keywords: AFLP; genetic diversity; native potato; Solanum spp.; characterization.
Introducción
La mayor diversidad genética de Solanum tuberosum "papa" silvestre y cultivada se encuentra principalmente en la zona temperada y en las partes altas de los andes (Valverde 2007). La papa es una planta perenne de la familia de las solanáceas (Struik 2007), se originó en los andes y fue introducida a la dieta europea en el Siglo XVI (Rocha et al. 2010), es un cultivo muy importante en todo el mundo y es esencial para la seguridad alimentaria global. Se propaga generalmente por clonación, es altamente heterocigota y autotetraploide; es el cuarto alimento cultivado en el mundo después del trigo, el arroz y el maíz (The Potato Genome Consortium 2011, Maga 1994, Gilsenan et al. 2010). En el año 2007, más 325 millones de toneladas de papas se producían en todo el mundo y era consumida en todo el planeta alrededor de 75 a 95 kg per capita al año (Fontes 2005) y desde el punto de vista nutricional, constituye un alimento altamente energético, debido a su alto contenido de carbohidratos, vitaminas y minerales (FAO 2008), constituye la base de la alimentación del poblador peruano.
En el Banco de Germoplasma del Centro Internacional de la Papa, se custodian más de cinco mil variedades cultivadas de papa, de las cuales aproximadamente 3500 son papas nativas provenientes de nueve países de América latina (CIP 2004). De éstas, casi 2000 son del Perú (Gómez et al. 2006). En Perú, las papas nativas cubren el 80% del área papera total del país (Castillo 2004).
El AFLP (Amplified fragment length polymorphism, polimorfismos en la longitud de los fragmentos amplificados) se ha utilizado para estudiar variación genética de cepas o especies estrechamente relacionadas, principalmente de plantas, hongos, y bacterias, realizando el mapeo genético de las poblaciones en estudio. Además se ha utilizado para el análisis de la diversidad genética de papas en diferentes regiones del mundo (Fang et al. 2011, Akkale et al. 2010, Nunziata et al. 2010, Esfahani 2009, Solano et al. 2007, Spooner et al.2005).
El presente trabajo pretende evaluar la diversidad genética de las papas nativas del distrito de Chungui, mediante marcador molecular AFLP.
Materiales y métodos
Material biológico.- Las papas nativas utilizadas fueron colectadas en la localidad de Sillaccasa - Unión Libertad de Rumichaca (13°1113.65"S; 73°3728.42"W) a una altitud promedio de 3660 m sobre el nivel del mar, en el distrito de Chungui, provincia La Mar, departamento de Ayacucho. En total se analizaron 25 morfotipos de papas nativas que fueron sembrados en el vivero de la de la Universidad Nacional de San Cristóbal de Huamanga utilizando para ello sustrato conteniendo tierra agrícola, tierra negra y arena en proporción de 3:2:1 respectivamente. Se logró introducir in vitro los primeros brotes de la planta para la micropropagación utilizando el medio de cultivo Murashigue y Skoog (1962).
Extracción de ADN.- La extracción de ADN se realizó empleando el método CTAB (CIP 1997) modificado, para lo cual se tomaron las hojas y tallos de las plántulas jóvenes 4 a 6 semanas del cultivo in vitro. La calidad y la cantidad de ADN fue verificada por el método de electroforesis en geles de agarosa al 1% (p/v), para la carga de las muestras al gel de agarosa se empleó 1µL de ADN genómico el cual fue diluido con 9 µL del colorante de carga SALB 1X. Como tampón de corrida se utilizó TBE 1X obtenidos de la dilución del TBE 10X. Para el cálculo de la concentración de ADN de las muestras se comparó con la intensidad de la primera banda del estándar del ADN Lambda (λ) digerida con la endonucleasa de restricción PstI de una concentración de 280 ng/µL. Las muestras de ADN fueron diluidas en agua Milli Q libre de nucleasas hasta alcanzar una concentración de 50 ng/µL.
Técnica de AFLP
Digestión del ADN genómico.- 400 ng de ADN genómico fueron digeridos enzimáticamente con 5U de EcoRI (Fermentas) y con 1U de MseI (Invitrogen), 4 µL de buffer de restricción 5X. La mezcla de reacción se incubó a 37 ºC toda la noche para asegurar una digestión completa, transcurrido este tiempo fue verificado el producto de la digestión empleando geles de agarosa al 1% (p/v) observándose un barrido del ADN, luego el producto de la digestión se llevó a -20 ºC para su conservación y posterior empleo.
Ligación de adaptadores.- La ligación de los adaptadores se realizó con 2U de T4 ADN ligasa (Invitrogen), 50 pmol de MseI adapter y 5 pmol del adapter EcoRI y agua milli Q libre de nucleasas, el mix así preparado se incubó a temperatura ambiente (25 ºC) toda la noche.
Reacción de pre amplificación (00/00).- La concentración final del mix para la amplificación pre selectiva consistió de 5 µL de ADN digerido y ligado diluido (2:7) con buffer PCR 1X, 250 µM DNTP (Promega), 50 ng de EcoRI+0 (Invitro gen), 50 ng de MseI (Invitro gen), 1U de taq polimerasa (Promega) y 1.5 mM de MgCl2 (Promega), llevando al programa de termociclador consistente en 25 ciclos 1er ciclo (72 °C/2 min), 2° ciclo (94 °C/4 min), 3° - 24° ciclo (94 °C/30 s, 56 °C/ 60 s), y finalmente de 72 °C/5 min. La verificación de la calidad del proceso consistió en la observación de los pre amplificados a través de geles de agarosa al 1%(p/v), observándose la presencia de un barrido más corto a manera de una banda gruesa tenue.
Amplificación selectiva (+3/+3).- Se emplearon combinaciones de iniciadores: EcoRI +3 – ACT MseI +3 CAG (E38 – M49); EcoRI +3 – ACA, MseI +3 – CAG (E35 – M49); EcoRI +3 – ACG- MseI +3 – CAT (E37 – M50). Para ello se empleó 0.75U de Taq polymerasa Promega (5U/µL), 0.3mM de DNTPs, 100ng de EcoRI+3 y 15ng de MseI +3. El producto de la amplificación selectiva se analizó mediante electroforesis en geles de agarosa al 1% (p/v), a 80 voltios durante 15 a 20 minutos, para ello se tomó 4 µL del producto de la amplificación selectiva con 6 µL del colorante SALB1X, observándose la presencia de banda.
Análisis de agrupamiento.- El análisis de agrupamiento se realizó mediante el método de UPGMA o método de agrupamiento de pares no ponderados usando la media aritmética. El cálculo del índice de similitud, análisis de agrupamiento y su grado de correlación fueron realizados con la ayuda del software NTSYS-pc versión 2.10p (Applied Bioestatistic Inc., Setauket, Nueva York, EE.UU)
Resultados y discusión
Uso de los marcadores moleculares AFLP en estudios de diversidad genética.- El presente trabajo constituye el primer estudio de diversidad genética de papas nativas usando marcadores AFLP en las comunidades campesinas del distrito de Chungui. Si bien, los marcadores morfológicos permiten estudiar y diferenciar los cultivos de papa, los marcadores moleculares pueden diferenciarlos con mayor exactitud (Onamu et al. 2012).
En el estudio evolutivo filogenético de papas realizado por Raker y Sponner (2002), así como también en el estudio de la diversidad genética de papas nativas realizadas por Soto (2006), ambos empleando marcadores moleculares microsatélites, no fue posible diferenciar grupos homogéneos de especies y tampoco hubo una buena diferenciación por zona ni por ploidía, razón por la cual Soto (2006) sugirió el empleo de marcadores moleculares de tipo AFLPs para un estudio más detallado.
El poder discriminatorio del empleo de marcadores moleculares AFLP reside en la generación de un gran número de bandas por cada combinación de iniciadores, puesto que permite amplificar muchas regiones del genoma de una especie como menciona Vos et al. (1995). El estudio de la diversidad genética mediante AFLP, superan complicaciones de clasificación mediante características fenotípicas, por el hecho que los marcadores moleculares se basan directamente en el análisis del ADN, los cuales son de utilidad no sólo en la caracterización de genotipos, sino también en estudios de similitud y distancia genética tal como menciona Bonamico (2004). Aunque Koskinen (2004) señala la importancia del número de bandas para la confiabilidad de los resultados en estudios de diversidad genética empleando los marcadores moleculares.
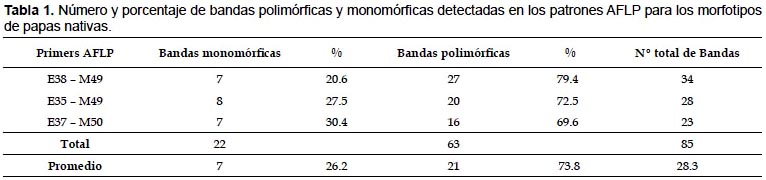
En el presente estudio, empleando la combinaciones de 3 iniciadores se generó un total de 85 bandas claramente diferenciables, de las cuales 63 fueron polimórficas las mismas que representan el 73.8%, estos resultados son similares a los obtenidos por Akkale et al. (2010). Sin embargo, la combinación de iniciadores E37 – M50 (Fig. 1) generó un total de 16 bandas polimórficas en comparación a las reportadas por Kim et al. (1998) quienes encontaron 64 bandas polimórficas; esta diferencia podría explicarse debido al gran número de especies con diferentes ploidias, más aun tratándose de Solanum spp. compuesta por 8 especies con diferentes ploidías (CIP -FEDECCH 2006). De otro lado, no se descarta que la diferencia del número de bandas generadas por una misma combinación de iniciadores puedan variar de una población a otra, así como Griffits et al. (2002) señalan que la composición genética de la población está relacionada con los procesos básicos, por lo tanto se debe examinar los cambios en la composición de la población debidos a la inmigración de individuos procedentes de otras poblaciones.

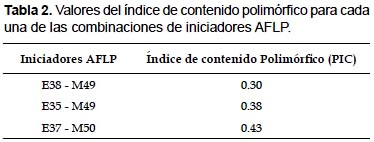
Índice de contenido polimórfico (PIC).- De la Tablas 1 y 2, la combinación de iniciadores E37-M50 mostraron 23 bandas, de las cuales 69% fueron polimórficas y un valor de PIC 0.43, la combinación E35 – M49 generaron 28 bandas de las cuales 72% fueron polimórficas y con un valor de PIC de 0.38, la combinación de iniciadores E38 – M49 generó 34 bandas de las cuales 79% fueron polimórficas con un valor de PIC de 0.3, por lo tanto, la combinación E37 – M50 demostró ser más informativo para detectar la variabilidad presente en los morfotipos analizados de papas nativas del distrito de Chungui. Los valores de PIC encontrado no tienen relación con el número de bandas encontrado por Esfahani et al. (2009), en contraste con lo encontrado por Bonamico et al. (2004) y Alagón y Rosas (2008), quienes mencionan que el aumento del valor de PIC es proporcional al aumento del número de bandas polimórficas.
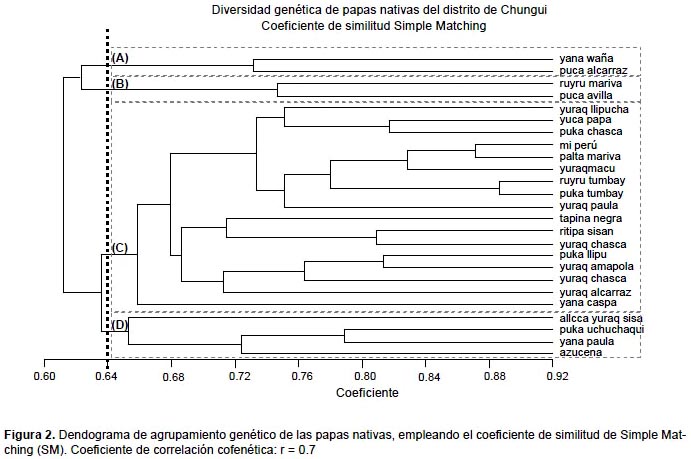
Análisis de similitud entre la matriz básica de datos y el dendograma mediante el coeficiente cofenético "r".- La elección del coeficiente de similitud es una parte crucial para la estimación del nivel de relación entre los genotipos. Como se puede apreciar en la Figura 2, el valor del coeficiente de correlación cofenética "r" fue de 0.7; lo cual indica que el dendograma así obtenido presenta poca distorsión con respecto a los datos de la matriz original, según Rohlf (1970) este valor de "r" corresponde a un nivel de relación adecuado.
Análisis de agrupamiento de la diversidad genética.- Varios investigaciones han reportado baja correlación entre marcadores morfológicos y moleculares para varios cultivos (Karuri et al. 2010, Koehler-Santos et al. 2003, Ferriol et al. 2004). En nuestro estudio, utilizando el AFLP logró una alta correspondencia en la identificación de nombres locales por parte de los agricultores conservacionistas de la comunidad de Chungui. Esta precisión en la clasificación nominal de papas nativas por los campesinos conservacionistas también fue encontrada en la comunidad de Chahuaytire, por Quiros et al. (1990), Zimmerer (1991) y Rojas (2007).
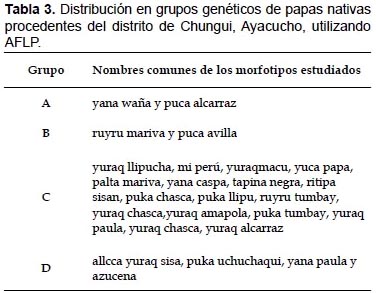
Con un coeficiente de similitud de SM de 0.65 se obtuvo cuatro grupos genéticos: grupo "A" (yana waña y puca alcarraz); grupo "B" (ruyru mariva y puca avilla); grupo "C" (yuraq llipucha, yuca papa, yuraq chasca, miperú, palta mariva, yuraqmacu, ruyru tumbay, puka tumbay, yuraq paula, tapina negra, ritipa sisan, puka chasca, puca llipu, yuraqamapola, yuraq chasca, yuraq alcarraz, yana caspa) y grupo "D" (allqa yuraq sisa, puca uchuchaqui, yanapaula y azucena).
La similitud genética de los morfotipos de papas nativas estuvieron entre 0.61 y 0.89, con una similitud media de 0.75, lo cual representa a una mayor similitud genética a lo reportado por Soto (2006), quien reporta una media de similitud genética de 0.62, este mayor nivel de similitud se debería a que el estudio de Soto (2006) hace referencia a papas nativas de diferentes departamentos.
En la Tabla 3, los morfotipos yana waña, puca alcarraz del grupo "A" y las accesiones ruyru mariva y puca avilla del grupo "B" son de pulpa pigmentadas al 100% pero las accesiones del grupo "C" no coinciden con esta relación, en el grupo "C" se encuentran papas pigmentadas y no pigmentadas, de igual manera no existe ninguna relación con los otros caracteres descritos en la caracterización morfológica. Mediante el análisis AFLP se determinó que no existe duplicado, a pesar que pueden o no presentar caracteres fenotípicos muy similares, puesto que el genotipo pudiera estar enmascarado por el ambiente como señala Griffiths et al. (2002).
Los marcadores moleculares no siempre pueden coincidir con los caracteres morfológicos puesto que las enzimas de restricción empleadas para el análisis AFLP no discriminan genes, las secuencias de reconocimiento de MseI y EcoRI no necesariamente pueden estar ubicadas en los genes, además las características morfológicas como la pigmentación, la forma del cuerpo están determinados por muchos genes que participan en complejas rutas metabólicas Griffiths (2002) este hecho es confirmado por Solano (2007) quien en su estudio para determinar la relación existente entre los datos morfológicos y los datos analizados usando marcadores moleculares AFLP obtuvo un bajo nivel de correlación (r: –0.09).
El cultivo de las papas nativas presenta una alta diversidad genética tanto en flores, como en tubérculos que varían en intensidad, combinaciones y tonalidades de colores. Esta diversidad morfológica fue ratificada por el análisis molecular de las 25 morfotipos estudiados, al respecto Gómez y Roca (2008) mencionan que esta diversidad se debe en gran parte a que las papas nativas crecen en condiciones agroecológicas muy particulares, se suma también la cosmovisión del agricultor de conservar y sembrar la diversidad en diferentes pisos ecológicos. Esfahani et al. (2009) menciona que la alta variación genética de papas por regiones tiene muchas causas, entre ellas su alta heterocigocidad de especies tetraploides con un gran número de genes.
La Figura 2 nos muestra que a un coeficiente de similitud de 0.64 los morfotipos de papas nativas se dividieron en 4 grupos genéticos (A, B, C y D). Existen morfotipos con mayor similitud genética (0.88) que corresponde a los morfotipos ruyru tumbay y puca tumbay. Debido a esta gran variabilidad genética, las papas cultivadas son altamente heterocigotos: el cruzamiento aún dentro de la misma variedad, produce una progenie altamente variable. Por esta razón, la pureza varietal solo podría mantenerse por reproducción asexual o vegetativa (Arce 2002).
Agradecimientos
Nuestro agradecimiento al Laboratorio de Biología Molecular del INIA – Lima por su apoyo en la realización de los análisis.
Literatura citada
Akkale C, Yildirim Z, Yildirim MB, Kaya C, Öztürk G. & B. Tanyolaç. 2010. Assessing genetic diversity of some potato Solanum tuberosum L.) genotypes grown in Turkey by using AFLP marker technique. Turkish Journal of Field Crops 15(1):73-78.
Alagón T. & R. Rosas. 2008. Caracterización Molecular de Mashua (Tropelum tuberosum) de las comunidades campesinas de Cusco y Huanuco. Arequipa. Tesis de pregrado del programa profesional de ingeniería Biotecnológica de la Universidad Católica de Santa María.
Arce F. 2002. El cultivo de la patata. Ed. Mundi-Prensa. Madrid.
Bonamico N., J. Aisassa, M. Ibañez, M. Di Renzo, D. Diaz. & J. Salemo. 2004. Caracterización y clasificación de híbridos simples de maíz con marcadores SSR. INTA, Argentina RIA, 33(2): 129-144.
Castillo A. 2004. Raíces y tubérculos peruanos, una herencia milenaria para el futuro. INITIRIMAY. Boletín virtual N° 17.
CIP (Centro Internacional de la Papa). 2004. Exitosa Restauración de Papa Nativa. <http://www.cipotato.org/news/pressreleases/espanol/papanatv.htm>. Acceso 10/8/2013.
CIP (Centro Internacional de la Papa). 1997. Protocolos de laboratorio de biología molecular-Tipificación genética. En: Ghislain, M., Zhang, D. & Herrera, M. R (edit). Departamento de Recursos Genéticos. Manual de Capacitación CIP. Lima, Perú. pp. 30.
CIP-FEDECCH (Centro Internacional de la Papa-Federacion Departamental de Comunidades Campesinas de Huancavelica). 2006. Catálogo de variedades de papa nativa de Huancavelica, Perú. Centro Internacional de la Papa. Lima. pp: 206 <http://cipotato.org/wp-content/uploads/2014/08/003524.pdf> Acceso 10/8/2013.
Fang W., L. Fangdi, W Jian, Z Yun & S. Haihong. 2011. Genetic Diversity of the Selected 64 Potato Germplasms Revealed by AFLP Markers. <http://biopublisher.ca/index.php/mpb/article/html/132/>. Acceso 11/06/2013.
Esfahani S.T., B. Shiran, and G. Balali 2009. AFLP markers for the assessment of genetic diversity in european and North American potato varieties cultivated in Iran. Crop Breeding and Applied Biotechnology 9: 75-86. http://dx.doi.org/10.12702/1984-7033.v09n01a11.
FAO. 2008. Potato World: World Potato Production. Available from: <http://www.fao.org/potato-2008/en/world/index.html>. Acceso 29⁄10⁄2012>.
Ferriol M., B. Pico, D. Fernandez & F. Nuez. 2004. Molecular diversity of germplasm collection of squash (Cucurbita moschata) deter- mined by SRAP and AFLP markers. Crop Sci. 44:653-664.
Fontes F. 2005. Cultura da batata. In Fontes PCR (ed.) Olericultura teoria e prática. Editora Universidade Federal de Viçosa, Viçosa, p.322- 343.
Gilsenan C., R. Burke & C. Barry-Ryan. 2010. A study of the physicochemical and sensory properties of organic and conventional potatoes (Solanum tuberosum) before and after baking. International Journal of Food Science & Technology 45(3): 475-481.
Gómez R. 2006. Guía para las caracterizaciones morfológicas básicas en colecciones de papas nativas. En manual para caracterización in situ de cultivos nativos. Instituto Nacional de Investigación y Extensión Agraria – INIEA. pp. 26-50.
Gomez R. & W. Roca. 2008. Papas nativas del Perú (Año internacional de la papa 2008). Catálogo de variedades y usos gastronómicos. Ministerio de Agricultura, Lima.
Griffiths F., H. Miller, T. Suzuki, C. Lewontin, & M. Gelbart. 2002. Genética. Séptima Edición. Edit. Mc Graw Hill Interamericana.
Karuri H W., E. M. Ateka, R. Amata, A. B. Nyende, A. W. Muigai, E. Mwasame & S. T. Gichuki. 2010. Evaluating diversity among Kenyan sweet potato genotypes using morphological and SSR markers. Int. J. Agric. Biol. 12:33-38.
Kim J.H., H. Joung, H.Y. Kim & Y.P. Lim. 1998. Estimation of Genetic Variation and Relationship in Potato (Solanum Tuberosum L.) Cultivars Using AFLP Markers. American Journal of Potato Research 75 (2): 107-12. http://dx.doi.org/10.1007/BF02883885.
Koehler-Santos P., A. L. Dornelles & L. B. de Freitas. 2003. Characterization of mandarin citrus germplasm from southern Brazil by morphological and molecular analysis. Pesq. Agropec. Bras. 38:797-806.
Koskinen M.T., H. Hirnoven, P.A. Landry & C. Primmer. 2004. The benefits of increasing the number of microsatellites used in genetic population studies: an empirical perspectiva. Hereditas 141:61-67.
Maga J. 1994. Potato flavour. Food Review International, 10, 1–48.
Murashige T. & F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15(3): 473-497. http://dx.doi.org/10.1111/j.1399-3054.1962.tb08052.x.
Nunziata A., V. Ruggieri, N. Greco, L. Frusciante & A. Barone. 2010. Genetic Diversity within Wild Potato Species (Solanum spp.) Revealed by AFLP and SCAR Markers. American Journal of Plant Sciences, 1: 95-103.
Onamu R., J. Legaria, J. Sahagún, J. Rodríguez & N. Pérez. 2012. Análisis de marcadores morfológicos y moleculares en papa (Solanum tuberosum L.). Revista Fitotecnia Mexicana, vol. 35, núm. 4, octubre-diciembre, 2012, pp. 267-277.
Quiros C., S. Brush, D. Douches, K. Zimmerer & G. Huestis. 1990. Biochemical and folk assessment of variability of Andean cultivated potatoes. Economic Botany 44(2): 254-266.
Raker C. & D. Spooner. 2002. Chilean tetraploid cultivated potato, Solanum tuberosum, is distinct from the Andean populations: Microsatellite data. Crop Sci. 42:1451-1458.
Rocha E., L. Paiva, H. de Carvalho & C. Guimarães. 2010. Molecular characterization and genetic diversity of potato cultivars using SSR and RAPD markers. Crop Breeding & Applied Biotechnology [serial on the Internet]. (2010, Sep), [cited February 10, 2011]; 10(3): 204-210.
Rohlf F. 1970. NTSYS pc: Numerical Taxonomy and Multivariate Analysis System, version 201. Departament of Ecology and Evolution State University of New York.
Rojas P.D. 2007 Análisis de la diversidad genética de papas nativas (Solanum sec. Petota) de la comunidad de Chahuaytire, integrante del Parque de la Papa (Pisaq-Cusco), y de las papas nativas repatriadas por el Centro Internacional de la Papa usando marcadores microsatélites. Tesis, Biologo, Facultad de Ciencias Biológicas Universidad Nacional Mayor de San Marcos. Tesis de pregrado de la carrera profesional de biología de la UNMSM.
Solano J., D. Morales & L. Anabalón. 2007. Molecular description and similarity relationship among native germoplasma potatoes (Solanum tuberosum ssp. Tuberosum L.) using morphological data and AFLP markers. Electronic Journal of Biotechnology 10(3):436-443. http://dx.doi.org/10.2225/vol10-issue3-fulltext-14.
Soto T.J. 2006. Análisis de la diversidad genética de papa nativa (Solanum spp.) de los departamentos de Ayacucho, Cajamarca, Cuzco, Huancavelica y Puno-Perú, mediante el uso de marcadores moleculares microsatélites. Tesis, Biologo, mención Biología Celular y Molecular. Facultad de Ciencias Biológicas Universidad Nacional Mayor de San Marcos. <http://cybertesis.unmsm.edu.pe/bitstream/cybertesis/806/1/soto_tj.pdf>. Acceso 12/05/2012.
Spooner D., K. McLean, G. Ransay, R. Waugh, & G. Bryan. 2005 A single domestication for potato based on multilocus amplified fragment length polymorphism genotyping. Proceedings National Academy of Sciences (PNAS) (USA). ISSN 0027-8424. 2005. 102(41): 14694-14699.
The Potato Genome Sequencing Consortium. 2011. Genome Sequence and Analysis of the Tuber Crop Potato. Nature 475 (7355): 189-95. doi:10.1038/nature10158.
Struik P. 2007. Above-ground and below-ground plant development. In: Potato Biology and Biotechnology- Advances and Perspectives (edited by D. Vreugdenhil, J. Bradshaw, C. Gebhardt, F. Govers, D.L. MacKerron, M.A. Taylor & H.A. Ross). Pp. 219–236. Oxford: Elsevier.
Valverde R. 2007. Mapeo genético y detección de QTLS en especies de Solanum. Agronomía Costarricense [serial on the Internet]. (2007, July), [cited March 25, 2011]; 31(2): 31-47. Available from: Fuente Académica.
Vos P., R. Hogers, M. Bleeker, M. Reijans, T. van de Lee, M. Hornes, A. Friters, et al. 1995. AFLP: A New Technique for DNA Fingerprinting. Nucleic Acids Research 23 (21): 4407-4414. doi:10.1093/nar/23.21.4407.
Zimmerer K. 1991. Managing diversity in potato and maize fields of the Peruvian Andes. J. Ethnobiol. 11(1): 23-49.
Fuentes de financiamiento:
La presente investigación fue financiada por el Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica del Perú (CONCYTEC) mediante la Subvención PROCYT Nº 233-2008-CONCYTEC-OAJ.
Presentado: 03/07/2014
Aceptado: 15/12/2014
Publicado online: 30/12/2014

