










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. vol.25 no.2 Lima abr./jun. 2018
http://dx.doi.org/10.15381/rpb.v25i2.14687
TRABAJOS ORIGINALES
Solapamiento de la dieta de Passer domesticus y Zonotrichia capensis en un agroecosistema de la costa central del Perú
Diet overlap of the invasive House Sparrow and the native Rufous-collared Sparrow in an agroecosystem of Central Coast of Peru
Katherine Menacho 1, Letty Salinas* 1,2 y César Arana 1,3
1 Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos.
2 Departamento de Ornitología, Museo de Historia Natural, Universidad Nacional Mayor de San Marcos.
3 Departamento de Ecología, Museo de Historia Natural, Universidad Nacional Mayor de San Marcos.
Resumen
Se analizó el solapamiento de las dietas de dos especies de gorriones en un agroecosistema de la costa central del Perú, una de las especies es invasora (Passer domesticus), mientras que la otra es nativa (Zonotrichia capensis). El presente trabajo muestra que a pesar de que Z. capensis tiene una dieta más diversa (H’= 2.07) que P. domesticus (H’=1.63), presentan alta similitud entre ellas (Dmax=0.15, p>0.05). El índice de Schoener fue ɸ=0.73 y junto con en el análisis de escalamiento multidimensional no métrico muestran un amplio solapamiento en la dieta de ambos gorriones en los diferentes meses de evaluación. Estos resultados permiten concluir que existe un potencial impacto de la invasión de Passer domesticus sobre la especie nativa Zonotrichia capensis por presentar un marcado solapamiento del alimento.
Palabras clave: Especies invasoras; Ica; nicho trófico; solapamiento de nicho; Aves.
Abstract
We analyse the diet overlap of two species of sparrows in an agroecosystem of the Central Coast of Peru, one of the species is invasive House Sparrow, while the other is native Rufous-collared Sparrow. We show that although native species has a more diverse diet (H'= 2.07), than invasive species (H' = 1.63), they present high similarity between them (Dmax = 0.15, p> 0.05). The Schoener index was ɸ = 0.73 and together with the non-metric multidimensional scaling analysis they show a broad diet overlap of both sparrows in the different months of evaluation. These results allow to conclude that there is a potential impact of the invasion of House Sparrow on the native species Rufous-collared Sparrow by food overlap.
Keywords: Zonotrichia capensis; Passer domesticus; Ica; niche overlap; birds.
Introducción
La competencia que involucra especies invasoras es particularmente importante debido a que puede causar la extinción de especies nativas (Gurnell et al. 2004). Esto es más preocupante, si se toma en cuenta que la introducción no controlada de especies exóticas, es considerada una de las mayores amenazas a la biodiversidad mundial, siendo señalada actualmente como la segunda causa de extinción de especies, después de la destrucción de hábitats (Zaret & Paine 1973, Vitousek 1990, Lambertini et al. 2011).
En el caso de las aves, una de las especies invasoras con mayor éxito es el gorrión Passer domesticus (Linnaeus, 1758), naturalde Europa y el Norte de África, que actualmente ha ampliado su distribución a todos los continentes con excepción de la Antártida (Anderson 2006). Esta ave llegó a Norteamérica entre 1852-1853 y para 1915 ya había colonizado Canadá y Estados Unidos, continuando su avance a través de México, Guatemala, Costa Rica y Panamá (Anderson 2006, MacGregor et al. 2009, Moulton et al. 2010). En Sudamérica existieron múltiples puntos de introducción: en Buenos Aires en 1872, en Santiago de Chile en 1904 y en Río de Janeiro en 1905; desde entonces su presencia y avance ha sido reportada frecuentemente (Baker 1995). Al Perú, llegó proveniente de Chile, donde varios individuos fueron liberados en Santiago en 1904 (Baldini & Pancel 2002, Anderson 2006). Para 1924 alcanzó Antofagasta, en 1940 el límite norte de Chile, y en 1951 fue registrada con frecuencia en Tacna e Ilo; alcanzando luego Lima y avanzando hacia la costa norte (Koepcke 1952, 1964).
En la actualidad P. domesticus se encuentra distribuido en toda la costa del Perú, tanto en áreas urbanas como rurales (Gonzáles 2004, Salinas 2007, Salinas et al. 2007, Takano & Castro 2007, Angulo et al. 2010, Schulenberg et al. 2010, Pulido et al. 2013, Salinas et al. 2013). Las posibles consecuencias de su introducción en nuestro país, aún no han sido estudiadas, sin embargo, estudios en otros países muestran que la presencia de P. domesticus es capaz de modificar de manera significativa las comunidades de los ecosistemas invadidos, al competir eficientemente por los recursos ocasionando la disminución de poblaciones de aves nativas (Anderson 2006, MacGregor et al. 2009, Feare 2010). Estos resultados podrían estar relacionados con el comportamiento agresivo observado en P. domesticus, que ataca constantemente a las especies nativas en sitios de forrajeo; así como con su gran capacidad de explotar recursos sobre todo en áreas cercanas a poblados y ciudades (Kalinoski 1975, Gowaty 1981, MacGregor et al. 2009).
En particular, Goodall (1946) y Koepcke (1964) consideraron que P. domesticus, podría afectar al gorrión nativo Zonotrichia capensis (Statius Muller, 1776) y a otras aves que se alimentan de semillas, debido a que compartirían parcialmente el nicho, lo que podría causar un desequilibrio en sus poblaciones. A esto se le debe añadir que, las ciudades, poblados y agroecosistemas de la costa peruana ofrecen un ambiente favorable para especies exóticas, en especial a las que se alimentan de semillas (Chace & Walsh 2006), lo que podría otorgar mayores ventajas al gorrión invasor. Passer domesticus tiene un elevado éxito al invadir agro-ecosistemas convirtiéndose en una auténtica plaga, que consume grandes cantidades de semillas de los cultivos y las malezas, además de insectos (Torres-Vila et al. 2015, Singh et al. 2016), afectando la avifauna local (Sax et al. 2000, Anderson 2006). En el presente estudio se evalúa la existencia de solapamiento en la dieta del gorrión invasor P. domesticus y la especie nativa Z. capensis en un agroecosistema de Ica.
Materiales y métodos
Área de Estudio.- El área de estudio se encuentra en el departamento de Ica, en el fundo "La Catalina" (14°8’08.54" S - 75°40’44.46" O, 420 m de altitud) de la Sociedad Agrícola Drokasa (AGROKASA), con una extensión aproximada de 1100 ha (Fig. 1). Este fundo se encuentra dentro del dominio morfoclimático del Desierto del Pacífico, caracterizado por un clima desértico cálido, con temperaturas altas durante el día, humedad relativa generalmente mayor al 60%, precipitaciones poco frecuentes y vientos de moderados a fuertes (Pulido et al. 2007, 2013).Dentro de este fundo se pueden encontrar diferentes tipos de hábitats como los cultivos de espárragos, cultivos de vid, cercos vivos de Acacia cf. horrida y Prosopis pallida; monte
ribereño, y el desierto con árboles y arbustos xerofíticos (Pulido et al. 2007, Salinas et al. 2007).
Identificación de los componentes de la dieta.- Los especímenes de Zonotrichia capensis (n=133) y Passer domesticus (n=113), fueron colectados simultáneamente en los meses de enero, abril y julio de los años 2004 a 2011. Se empleó en cada evento un mismo grupo de redes de niebla para capturar y tomar datos reproductivos y morfológicos de los individuos liberándolos posteriormente, en el presente estudio se utilizaron
los contenidos estomacales sólo de los individuos que murieron accidentalmente en las redes. Los especímenes fueron trasladados al laboratorio donde se extrajeron los tractos digestivos que se fijaron en alcohol etílico 70º (Salinas 2001). El contenido de cada tracto digestivo fue vaciado en placas Petri, y se separaron los diferentes componentes identificables de la dieta (semillas, restos vegetales, piedras, restos de artrópodos, entre otros), con ayuda de una lámpara con lupa.
Para identificar las semillas e insectos, se utilizó un microscopio estereoscópico para tomar fotografías y compararlas con las colecciones de referencia elaboradas con plantas e insectos colectados en el área de estudio. Adicionalmente se consultaron colecciones especializadas digitales del Royal Botanic Garden KEW, Plants Database y Seed Biology.
Análisis de los datos.- Para el análisis de los datos, se obtuvo una lista de los taxa que componen la dieta de cada gorrión. Para cada componente de la dieta se calculó el porcentaje de volumen total (%V) disponiendo cada componente en la forma de un prisma rectangular y midiendo su alto, largo y ancho, también
el porcentaje de abundancia de cada componente (%N) es decir el número de unidades de cada componente dividido por el total de unidades de todos los componentes por cien y la frecuencia porcentual calculada como el porcentaje de contenidos estomacales que contenían cada tipo de componente dietario (%F). Estos valores fueron empleados para calcular el porcentaje del Índice de importancia relativa (Pinkas et al. 1971, George & Hadley 1979, Harrison et al. 1983, Duffy & Jackson 1986): IIRi= (%N + %V) x %F
Además se calculó la diversidad de la dieta con el índice H de Shannon-Wiener usando logaritmo en base 2 (Magurran 2004).
Se comparó las dietas de ambos gorriones empleando los valores de volumen de los taxa consumidos, con el Índice de similitud cuantitativo de Morisita – Horn (Horn 1966) y con un análisis de escalamiento multidimensional no métrico donde se consideraron todos ítems de los contenidos estomacales colectados en el mismo grupo de redes en el mismo mes de evaluación (Kenkel & Orlóci 1986).
Para determinar la significancia en las diferencias en los taxa consumidos por ambas especies, se empleó el test de Kolmogorov-Smirnov comparando la abundancia de cada ítem alimentario como ha sido recomendado para comparar comunidades y utilizado en comparación de dietas (Magurran 2004, Pérez & Balta 2011).
El análisis de solapamiento de la dieta se realizó empleando el índice de solapamiento de Schoener (Ojk), el cual es considerado como el más adecuado debido a que no requiere asunciones respecto a la naturaleza del proceso de competencia (Abrams 1980). Los valores del índice varían de 0 a 1, valores cercanos a 0 indican bajo solapamiento, mientras que valores cercanos a 1, indican un mayor solapamiento (Schoener 1968).
Resultados
Composición de la Dieta.- La dieta de los gorriones Zonotrichia capensis y Passer domesticus está compuesta por 27 ítems, 20 tipos de semillas y 7 de insectos (tanto adultos como larvas). Para Z. capensis las semillas representan 84.76% de sus ítems dietarios y los insectos el 15.24%; mientras que para P. domesticus 70.37% y 29.63%, respectivamente. La mayoría de los componentes de la dieta, fueron identificados, especialmente los más abundantes, quedando sin identificación algunos de poca abundancia, debido en gran parte a su avanzado estado de digestión (Tabla 1). Los componentes más importantes en función del %IIR fueron las semillas de Amaranthus sp., Portulaca oleracea, Eleucine indica, Setaria verticillata y los insectos del orden coleóptera (Fig. 2).
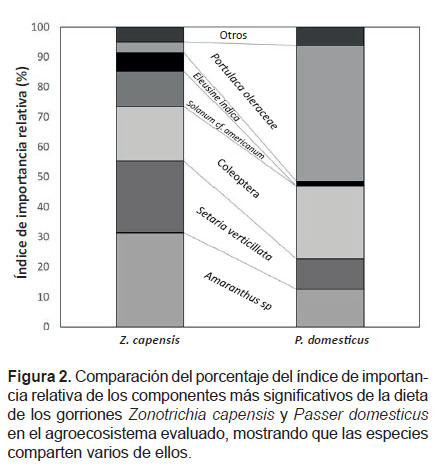
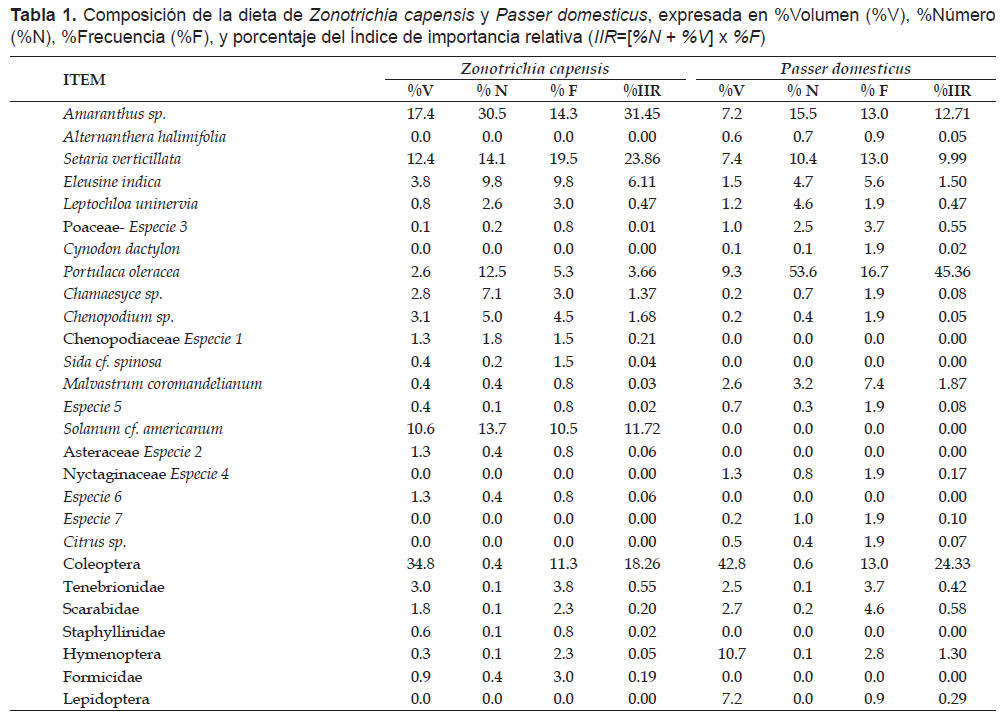
Diversidad y solapamiento de la dieta.- El índice de Shannon-Wiener, mostró que la dieta de Z. capensis (H= 2.07) es más diversa que la de P. domesticus (H= 1.63). El análisis con el índice de similitud cuantitativo de Morisita-Horn, mostró un valor de CH= 0.72, que demuestra el elevado grado de semejanza en la dieta de ambos gorriones. La prueba de Kolmogorov-Smirnov, comprobó que los taxa que componen la dieta de los gorriones y la proporción en la que los consumen son semejantes (Dmax= 0.15, p>0.05).
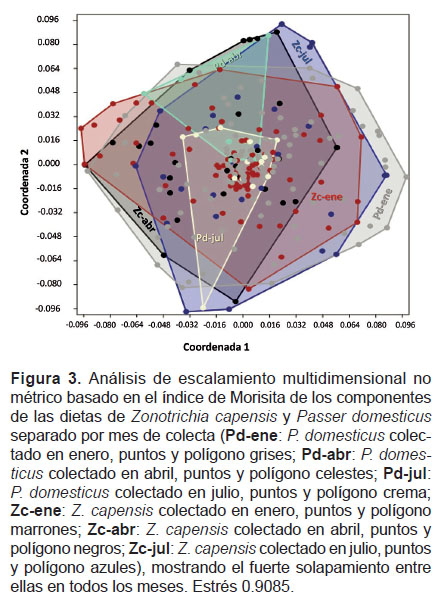
Finalmente, la prueba de solapamiento con el índice de solapamiento de Schoener (Ojk= 0.73), mostró que existe un elevado grado de solapamiento. Esto mismo se observa al realizar el análisis de escalamiento multidimensional no métrico (Fig. 3) donde las dietas de las dos especies en los diferentes meses de evaluación se ordenan en prácticamente un mismo espacio y siempre se sobreponen una con la otra.
Discusión
Composición de la dieta.-El análisis inicial de la dieta de Z. capensis y P. domesticus muestra que éstas se componen mayoritariamente de semillas de malezas de los campos de cultivo y una cantidad menor de insectos. Estos datos son comparables a los obtenidos en la reserva "El Charmolán" en Colombia, donde la dieta de Z. capensis estuvo compuesta en más del 70% por semillas y un porcentaje cercano al 20% de insectos (Cabrera et al. 2011). Otro estudio realizado en la Quebrada de La Plata en Chile, muestras que la dieta, estuvo compuesta de semillas que representaron entre 78% y 99% del total de alimentos consumidos, manteniendo en alrededor del 4.5% la proporción de insectos consumidos (López 1995).
Sin embargo, cuando Z. capensis se encuentra en otro tipo de ecosistemas, estos porcentajes varían, por ejemplo, en la isla Carabajal en Argentina, su dieta se compuso casi en la misma proporción de insectos y semillas (Alessio et al. 2005), y en la restinga del Parque Nacional Jurubatiba del Brasil, llegó a alimentarse en mayor proporción de insectos (62%), mientras que las semillas fueron sólo el 38% (Souza et al. 2008).
En cuanto a la dieta de P. domesticus, en lugares donde se encuentra naturalmente distribuido, por ejemplo, en los campos de cultivo de la Península Ibérica, las semillas representaron el 95%; mientras que los insectos sólo el 5% (Alonso 1985); y en el río Jarama-Madrid, su dieta se compuso de 75% de semillas y 25% de insectos (Sánchez 1986). En áreas donde P. domesticus es introducido, también muestra preferencia por las semillas; así en Punjab-India, el porcentaje de semillas fue 90%, mientras que los insectos fueron apenas 5% (Saini & Dhindsa 1991), igualmente en Jodhpur–India, las semillas representaron entre 70% y 90% de su dieta, y los insectos entre 30% y 10%, dependiendo de las estaciones (Rana & Idris 1987). Todas estas investigaciones coinciden en reportar la preferencia de P. domesticus por las semillas, resultados que coinciden con los reportados en el presente estudio.
Los presentes resultados muestran que la dieta de Z. capensis, está principalmente compuesta por semillas de malezas de las familias Amaranthaceae, Poaceae, Solanaceae y Portulacaceae, mientras que en los insectos destaca el orden Coleoptera. Resultados comparables fueron encontrados en estudios realizados en Colombia (Cabrera et al. 2011) y Chile (López 1995), donde se reportaron un marcado predominio de la familia Poaceae entre las semillas y del orden Coleoptera entre los insectos. Por otra parte, De la Peña & Pensier (2003) reportan el consumo de semillas pertenecientes a las familias Chenopodiaceae y Malvaceae y los géneros Chenopodium, Sida y Eleusine. Nuevamente si comparamos los resultados de este estudio con los encontrados en ecosistemas con mayor complejidad vegetal como la restinga brasileña, se observan diferencias notables en la dieta de Z. capensis, que prefiere alimentarse de las semillas de las familias Lauraceae, Burseraceae, Clusiaceae y Humiriaceae (Souza et al. 2008).
En el caso de P. domesticus los ítems de mayor importancia en su dieta son las semillas de las familias Portulacaceae, Amaranthaceae, Poaceae y Malvacea e insectos del orden Coleoptera. Esta composición es consistente con la reportada para este gorrión en ambientes cultivados de India (Saini & Dhindsa 1991, Rana & Idris 1987) y Estados Unidos (Gavett & Wakeley 1986), donde también destacan las semillas de las familias Poaceae y Amaranthaceae, y los insectos del orden Coleoptera. Estudios realizados en España muestran que P. domesticus consume semillas de los géneros Chenopodium (Chenopodiaceae) y Portulaca (Portulacaceae) e insectos de los órdenes Coleoptera e Hymenoptera (Sánchez 1986, Alonso 1985), taxones también encontrados en esta investigación.
Diversidad y solapamiento de la dieta.- Los resultados de diversidad, muestran que la dieta de Z. capensis (H’= 2.07) es más diversa que la dieta de P. domesticus (H’= 1.63). Esta mayor diversidad de alimentos puede deberse a que siendo Z. capensis una especie nativa, aprovecharía recursos poco abundantes, mientras que el gorrión invasor, que sólo habita estos ecosistemas las últimas décadas aprovecha sólo los recursos más abundantes. La dieta de P. domesticus analizada en su área de distribución natural mostró valores de diversidad H’= 2.06, en un área de cultivos cercana a Coria (Alonso 1985) y H’= 3.16 en las inmediaciones de Madrid (Sánchez 1986), que son muy superiores a los encontrados en la presente investigación.
En áreas donde Passer domesticus es una especie introducida, la diversidad de su dieta baja, como en los agroecosistemas de Punjab-India, donde el índice de diversidad fue H’= 0.75 (Saini & Dhindsa 1991). Esto podría deberse a que su dieta oportunista consiste en buscar recursos alimenticios que aporten la mayor tasa de energía, en el menor tiempo de búsqueda (Anderson 2006, Greenberg & Mettke-Hofmann 2001).
Existen menos datos de diversidad de la dieta de Z. capensis. En la isla Carbajal, provincia de Santa Fe-Argentina reportaron un valor H’= 2.89, siendo este superior al encontrado en esta investigación (Alessio et al. 2005).
El índice de Morisita – Horn (CH= 0.72) muestra alta similitud en la dieta de Z. capensis y P. domesticus. Adicionalmente se conoce por De la Peña (2011) que el comportamiento alimenticio de ambas especies es semejante. Ambas recolectan alimentos en el suelo, estrato herbáceo, ramas y follaje hasta los cuatro metros, con la diferencia que P. domesticus caza al vuelo los insectos.
Nuestros datos muestran un solapamiento de la dieta de ambos gorriones lo que significa un peligro potencial para las poblaciones del nativo Z. capensis si se incrementa la población del invasor P. domesticus.
Agradecimientos
Al Vicerrectorado de Investigación y Posgrado de la Universidad Nacional Mayor de San Marcos, por el financiamiento de los Proyectos de Investigación 071001161, 071001291, 081001291, 091001171, 101001271, 101001281, 111001221, 111001115, 121001161, 121001191. A la empresa AGROKASA por las ffacilidades brindadas en su fundo. Al SERFOR por los permisos de investigación (RD 0263-2014-MINAGRIDGFFS/DGEFFS, RD 0267-2014-MINAGRI-DGFFS/ DGEFFS, Autorización 051-2005-INRENA-IFFS-DCB). A los revisores anónimos por contribuir a mejorar el manuscrito. A Víctor Pulido, Joyce del Pino y Alejandra Arana por su ayuda en el trabajo de campo.
Literatura citada
Abrams P. 1980. Some comments on measuring niche overlap. Ecology61(1):44-49. https://doi.org/10.2307/1937153 [ Links ]
Alessio V., A. Beltzer, R. Lajmanovich, et al. 2005. Ecología alimentariade algunas especies de Passeriformes (Furnariidae, Tyrannidae, Icteridae y Emberizidae): consideraciones sobre algunosaspectos del nicho ecológico. En: Aceñolaza, F., ed. Temas debiodiversidad del Litoral Fluvial Argentino II. INSUGEO Magna Publicaciones. Tucumán. Pp. 441-482. [ Links ]
Alonso J.C. 1985. La alimentación del gorrión común Passer domesticus en áreas de cultivo de regadío extremeñas. Ardeola32:405-408. [ Links ]
Anderson T.R. 2006. Biology of the ubiquitous House Sparrow:From genes to population. Oxford University Press. New York. 560 pp. [ Links ]
Angulo F., T.S. Schulenberg & E. Puse. 2010. Las aves de los humedalesde Eten, Lambayeque, Perú. Ecología Aplicada 9(2):71-81. https://doi.org/10.21704/rea.v9i1-2.397 [ Links ]
Baker M. 1995. Environmental component of latitudinal clutch-size variation in house sparrows (Passer domesticus). The Auk 112(1):249-252. https://doi.org/10.2307/4088786 [ Links ]
Baldini A. & L. Pancel. 2002. Agentes de daño en el bosque nativo. Editorial Universitaria. Santiago de Chile. 408 pp. [ Links ]
Cabrera A., Y.C. Ortiz & J.J. Calderón. 2011. Dieta de Semilleros en un Bosque Subandino. Revista de Ciencias - Universidad de Nariño 1(1):1-8. [ Links ]
Chace J.F. & J.J. Walsh. 2006. Urban effects on native avifauna: a review. Landscape and Urban Planning 74(1):46-69. https://doi.org/10.1016/j.landurpblan.2004.08.007 [ Links ]
De La Peña M.R. & J.F. Pensier. 2003. Contribución de la flora en los hábitos alimentarios de las aves en un bosque del centro de la provincia de Santa Fe, Argentina. Ornitología Neotropical14:499-513. [ Links ]
De La Peña M.R. 2011. Observaciones de campo en la alimentación de las aves. FAVE 15(1):1-88. https://doi.org/10.14409/fave.v15i1.3108 [ Links ]
Duffy D.C. & S. Jackson. 1986. Diet studies of seabirds: a review of methods. Colonial waterbirds 9(1):1-17. https://doi.org/10.2307/1521138 [ Links ]
Feare C. 2010. Invasive bird eradication from tropical oceanic islands. En Genovesi P. & R. Scalera, eds. Aliens: The Invasive Species Bulletin Newsletter of the IUCN/SSC Invasive Species Specialist Group N° 30. Roma. Pp. 12-19. [ Links ]
Gavett A.P. & J.S. Wakeley. 1986. Diets of House Sparrows in urban and rural habitats. Wilson Bulletin 98:137-144. [ Links ]
George E.L. & W.L. Hadley. 1979. Food and habitat partitioning between rock bass (Ambloplites rupestris) and smallmouth bass(Micropterus dolomievi) young of the year. Transactions of the American Fisheries Society 108(3):253-261. https://doi.org/10.1577/1548-8659(1979)108<253:fahpbr>2.0.co;2 [ Links ]
Goodall J.D., A.W. Johnson & R.A. Philippi. 1946. Las aves de Chile:su conocimiento y sus costumbres. Tomo 1. Platt Establecimientos Gráficos S.A. Buenos Aires. 358 pp. [ Links ]
Gowaty P.A. 1981. Aggression of breeding eastern bluebirds (Sialia sialis) toward their mates and models of intra- and interspecificintruders. Animal Behaviour 29(4): 1013-1027. https://doi.org/10.1016/S0003-3472(81)80055-3 [ Links ]
Greenberg R. & C. Mettke-Hofmann. 2001. Ecological aspects of neophobia and neophilia in birds. Current Ornithology16:119-178. https://doi.org/10.1007/978-1-4615-12110_3 [ Links ]
Gurnell J., L.A. Wauters, P.W. Lurz, et al. 2004. Alien species and interspecific competition: effects of introduced easterngrey squirrels on red squirrel population dynamics. Journal of Animal Ecology 73(1):26-35. https://doi.org/10.1111/j.1365-2656.2004.00791.x [ Links ]
Harrison C.S., T.S. Hida & M.P. Seki. 1983. Hawaiian seabird feedingecology. Wildlife Society. California. 71pp. [ Links ]
Horn H.S. 1966. Measurement of "Overlap" in comparative ecologicalstudies. The American Naturalist 100:419-424. https://doi.org/10.1086/282436 [ Links ]
Kalinoski R. 1975. Intra and interspecific aggression in House Finchesand House Sparrows. Condor 77:375-384. https://doi.org/10.2307/1366086 [ Links ]
Kenkel N. C. & L. Orloci. 1986. Applying Metric and Nonmetric Multidimensional Scaling to Ecological Studies:Some New Results. Ecology 67:919–928. https://doi.org/10.2307/1939814 [ Links ]
Koepcke M. 1952. El Gorrión Europeo en el Perú. Revista Mar del Sur 22:63-72. [ Links ]
Koepcke M. 1964. Las Aves del Departamento de Lima. GráficaMorson. Lima. 156 pp. [ Links ]
Lambertini M., J. Leape, J. Marton-Lefevre, et al. 2011. Invasives: A Major Conservation Threat. Science 333(6041): 404–405. https://doi.org/10.1126/science.333.6041.404-b [ Links ]
López M.V. 1995. Dieta de Zonotrichia capensis (Emberizidae) yDiuca diuca (Fringillidae): efecto de la variación estacional delos recursos tróficos y la riqueza de aves granívoras en Chile central. Revista Chilena de Historia Natural 68:321-331. [ Links ]
MacGregor I., L. Morales, J. Quesada & J.E. Schondube. 2009. Relationship between the presence of House Sparrow (Passer domesticus) and Neotropical bird community structureand diversity. Biological Invasions 12:87-96. https://doi.org/10.1007/s10530-009-9432-5 [ Links ]
Magurran A. 2004. Measuring Biological Diversity. Blackwell Publishing. Oxford. 264 pp. [ Links ]
Moulton M., W. Cropper, M. Avery & L. Moulton. 2010. The earliestHouse Sparrow introductions to North America. Biological Invasions 12:2955-2958. https://doi.org/10.1007/s10530010-9692-0 [ Links ]
Pérez J. & K. Balta. 2011. Ecología de Phyllodactylus angustidigitus y P. gerrhopygus (Squamata: Phyllodactylidae) de la Reserva Nacional de Paracas, Perú. Revista Peruana de Biología 18(2):217-223. https://doi.org/10.15381/rpb.v18i2.232 [ Links ]
Pinkas L., M.S. Oliphant & I.L.K. Iverson. 1971. Food habits ofalbacore Bluefin tuna and bonito in California waters. State of California, Dept. of Fish and Game. California. 105 pp. [ Links ]
Plants Database. 2014. Fact Sheets & Plant Guides. (En línea). <https://plants.usda.gov/java/factSheet> Acceso 10/06/2013. [ Links ]
Pulido V., L. Salinas & C. Arana. 2007. Aves en el desierto de Ica. La experiencia en AGROKASA. Editorial Biblos. Lima. 323 pp. [ Links ]
Pulido V., L. Salinas & C. Arana. 2013. Aves en el desierto de la costa central del Perú. Editorial Barrón. Lima. 460 pp. [ Links ]
Rana D.B. & M. Idris. 1987. Food habits of the House Sparrow Passerdomesticus indicus in an arid environment. Japanese Journalof Ornithology 35:125-128. [ Links ]
Royal Botanic Garden KEW. Seed Information Database - SID. (En línea). < http://data.kew.org/sid/> Acceso 10/06/2013. [ Links ]
Saini H.K. & M.S. Dhindsa. 1991. Diet of the house sparrow in an intensively cultivated area. Japanese Journal of Ornithology 39:93-100. [ Links ]
Salinas L. 2001. Aves dispersoras de semillas de plantas leñosas en el bosque de Zárate, Lima. Tesis Bióloga con mención en Zoología. EAP Ciencias Biológicas, UNMSM. Lima, 158 pp. http://sisbib-03.unmsm.edu.pe/cgi-bin/koha/opac-detail.pl?biblionumber=74638# [ Links ]
Salinas L. 2007. Avifauna de Tacna en la ruta del censo de suri Pterocnemia pennata. INRENA. Lima. 25 pp. [ Links ]
Salinas L., C. Arana & V. Pulido. 2007. Diversidad, abundancia y conservación de aves en un agroecosistema del desiertode Ica, Perú. Revista Peruana de Biología 13(3):155-167. https://doi.org/10.15381/rpb.v13i3.2333 [ Links ]
Salinas L., C. Arana & A. Tello. 2013. Aves de El Olivar y otros parquesde Lima. Hatch Ediciones. Lima. 118 pp. [ Links ]
Sánchez F.J. 1986. Sobre la alimentación de los gorriones molinero y común (Passer montanus L. y P. domesticus L.), en inviernoy primavera. Ardeola 33:17-33. [ Links ]
Sax D. F. & J.H. Brown. 2000. The paradox of invasion. Global Ecology and Biogeography 9(5): 363-371. https://doi.org/10.1046/j.1365-2699.2000.00217.x [ Links ]
Schoener T.W. 1968. The Anolis lizards of Bimini: Resource partitioning in a complex fauna. Ecology 49:704-726. https://doi.org/10.2307/1935534 [ Links ]
Schulenberg T.S., D.F. Stotz, D.F. Lane, et al. 2010. Aves del Perú. Princeton Unity Press. Chicago. 656 pp. [ Links ]
Seed Biology. Seed ID Workshop. (En línea). < https://www.oardc.ohio-state.edu/seedid/> Acceso 10/06/2013 [ Links ]
Singh R., P. Kour & S. Koul. 2016. Comparative study of food habits and seasonal variations in the food of adult House Sparrow Passer domesticus indicus in some rural and urban areas of Jammu Province. International Journal of Advanced Research 4(7):951–958. https://doi.org/10.21474/ijar01/901 [ Links ]
Souza V., B.A. Loiselle & Alves M.A. 2008. Birds foraging for fruits and insects in shrubby restinga vegetation, southeastern Brazil. Biota Neotropica 8(4):21-31. https://doi.org/10.1590/S1676-06032008000400001 [ Links ]
Takano F. & N. Castro. 2007. Avifauna en el campus de la UniversidadNacional Agraria la Molina (UNALM), Lima – Perú. Ecología Aplicada 6(1,2):149-154. https://doi.org/10.21704/rea.v6i1-2.351 [ Links ]
Torres-Vila L., J. Ferrero-García, D. Martín-Vertedor, et al. 2015. Sparrow plagues in Extremadura (Western Spain) overfour centuries (1501–1900): A Spatio-temporal analysisof records from historical archives. Ardeola 62(1):19–33. https://doi.org/10.13157/arla.62.1.2015.19 [ Links ]
Vitousek P. 1990. Biological Invasions and Ecosystem Processes:Towards an Integration of Population Biology andEcosystem Studies. Oikos 57(1): 7-13. https://doi.org/10.2307/3565731 [ Links ]
Zaret T.M. & R.T. Paine. 1973. Species introduction in a tropical lake. Science 182(4111):449-455. https://doi.org/10.1126/science.182.4111.449 [ Links ]
Fuentes de financiamiento: Proyectos de Investigación VRIPUNMSM 071001161, 071001291, 081001291, 091001171, 101001271, 101001281, 111001221, 111001115, 121001161, 121001191.
Permisos de colecta: Servisio Nacional Forestal y de Fauna Silvestre (SERFOR) del Ministerio de Agricultura y Riego - permisos de investigación RD 0263-2014-MINAGRI-DGFFS/DGEFFS, RD 0267-2014-MINAGRI-DGFFS/DGEFFS, Autorización 051-2005-INRENAIFFS-DCB.
Información sobre los autores: KM realizó los análisis e identificación de la mayoría de las muestras y preparó el primer borrador del manuscrito, CA y LS realizaron el diseño del muestreo y las evaluaciones de campo, participaron en el análisis de la información y en la elaboración del manuscrito.
Los autores no incurren en conflictos de intereses.
*Autor de correspondencia
Email Katherine Menacho: katymj_1008@hotmail.com
Email Letty Salinas: lsalinass@unmsm.edu.pe
Email César Arana: caranab@unmsm.edu.pe
Presentado: 09/06/2017
Aceptado: 28/02/2018
Publicado online: 30/05/2018

