










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista Peruana de Biología
versión On-line ISSN 1727-9933
Rev. peru biol. vol.26 no.2 Lima abr./jun. 2019
http://dx.doi.org/10.15381/rpb.v26i2.16372
TRABAJOS ORIGINALES
Diversidad de mamíferos en el bosque montano de Carpish, Huánuco, Perú
Mammalian diversity of Carpish montane forest, Huánuco, Peru
Víctor Pacheco*,1,2 ORCID iD: https://orcid.org/0000-0002-1005-135X y Maggie Noblecilla1 ORCID iD: https://orcid.org/0000-0001-9770-4247
1 Universidad Nacional Mayor de San Marcos, Museo de Historia Natural, Av. Arenales 1256, Lima, Perú.
2 Universidad Nacional Mayor de San Marcos, Facultad de Ciencias Biológicas, Instituto de Ciencias Biológicas "Antonio Raimondi", Lima, Perú.
Resumen
A pesar de la alta diversidad de mamíferos conocidos para el Perú, son pocos los estudios en las yungas centrales, lo que no permite un adecuado conocimiento de su diversidad. Aquí se da a conocer los resultados de una evaluación de la diversidad de mamíferos de Carpish, Huánuco, un bosque montano en estado crítico de conservación. El estudio se realizó entre 2001 y 2002 con cinco expediciones en las localidades de Carpish de Mayobamba (2730 – 3080 m) y San Pedro de Carpish (1780 – 2400 m). Se emplearon métodos de captura, avistamientos y entrevistas, los que representan un esfuerzo de 28500 trampas noche y 190 redes noche. Registramos 35 especies de mamíferos en el área de estudio. Los órdenes más diversos fueron los murciélagos y roedores (74.3 %), estos incluyen cuatro especies endémicas para el país, cuatro especies en alguna categoría de conservación y el primer registro de la marmosita grácil Gracilinanus aceramarcae para Huánuco. La riqueza y los índices de diversidad (H' y Simpson) para roedores no muestran correlación significativa con respecto a la elevación. El roedor Thomasomys ischyrus y el murciélago Anoura aequatoris fueron las especies con mayor abundancia relativa. Nuestros registros, más los de la literatura y la revisión de colecciones científicas, sustentan la ocurrencia de 45 especies de mamíferos en el bosque de Carpish. Esta riqueza de especies, los endemismos y la acelerada pérdida de bosque son base para recomendar el establecimiento de un área de conservación que garantice la protección de los bosques montanos de Carpish.
Palabras clave: Roedores; Murciélagos; Thoma- somys; Anoura; Yungas centrales.
Abstract
Despite the high diversity of the well-known mammals of Peru, there are few studies in the central yungas, which does not allow an adequate knowledge of their diversity. Here we present the results of an evaluation of mammal diversity of Carpish, Huanuco, a montane forest in a critical state of conservation. The study was carried out between 2001 and 2002 with five expeditions in the localities of Carpish de Mayobamba (2730 – 3080 m) and San Pedro de Carpish (1780 – 2400 m). We used capture methods, sightings and interviews, which represent an effort of 28 500 trap-nights and 190 net-nights. We recorded 35 species of mammals in the study area. The most diverse orders were bats and rodents (74.3%); these include four endemic species for the country, four species in some conservation category and the first record of the gracile opossum Gracilinanus aceramarcae for Huánuco. Species richness and diversity index (H' and Simpson) for rodents show no significant correlation with respect to elevation. The rodent Thomasomys ischyrus and the bat Anoura aequatoris were the species with the highest relative abundance. Our records, plus those from literature and review of scientific collections, support the occurrence of 45 mammal species in the Carpish forest. This species richness, endemism and the accelerated forest loss are the basis for recommending the establishment of a conservation area that guarantees the protection of the Carpish's montane forests.
Keywords: Rodents; Bats; Thomasomys; Anoura; central yungas.
Introducción
La diversidad de mamíferos del Perú es estimada en 559 especies (Pacheco et al. 2018), y varía según las ecorregiones, siendo la región de selva baja la más diversa (Voss & Emmons 1996, Emmons & Feer 1997), seguida por las Yungas (Pacheco et al. 2009). La cordillera de Carpish, en el centro del Perú, Región de Huánuco, alberga uno de los más importantes bosques pluviales montanos en diversidad de plantas y animales en general, pero su diversidad de mamíferos es aún poco conocida.
Son pocos los estudios desarrollados en la Cordillera de Carpish. En el aspecto florístico se han descrito seis formaciones vegetales: estepa de gramíneas con arbustos dispersos, turberas de musgo con arbustos ericoides dispersos, monte esclerófilo, bosque anaranjado de cumbre, bosque del piso nebuloso y selva tropical superior (Beltrán & Salinas 2010). En fauna, se han reportado varias especies de roedores (Gardner & Patton 1976), incluyendo el cariotipo del roedor Chibchanomys orcesi (publicado como Anotomys leander en Gardner 1971); recientemente, una nueva especie de Akodon fue reportada por Jiménez y Pacheco (2016). Entre los anfibios anuros, Lehr y Rodríguez (2017) reportaron siete especies de Phrynopus (Eleutherodactylidae) en la Cordillera de Carpish de las 32 que hay registradas para Perú. Además, Carpish exhibe una alta diversidad de aves, entre las familias más importantes se registran Trochilidae (picaflores o colibrís), Ramphastidae (tucanes, tucanetas), Thraupidae (tangaras), Psittacidae (loros y pericos), Cotingidae (cotingas) y especies endémicas como el tiránido Uromyias agraphia Chapman 1919 y, el picaflor cobrizo Metallura theresiae Simon 1902 (Young & León 1999, Parker & O'Neill 1976).
Estos estudios sustentan la importancia del bosque de Carpish, cuya conservación constituiría una adecuada y necesaria representación de las yungas centrales del Perú; dado que, las unidades de conservación que protegen las yungas centrales son pocas, relativamente pequeñas y parciales, como para ser representativas de la diversidad biológica de estos ecosistemas. Entre las unidades de conservación con yungas se tiene al Santuario Nacional Pampa Hermosa en Junín con un área de 115.44 km2 y un rango altitudinal de 2200 m a 3800 m (MINAM 2012a) y al Parque Nacional Tingo María con un área de 47.77 km2 y elevaciones de 680 m a 1600 m (MINAM 2012b). Por otro lado, el Parque Nacional Cordillera Azul se extiende en las regiones de San Martín, Loreto, Huánuco y Ucayali, con un área de 13531.91 km2, de las cuales 781.83 km2 corresponden a la región de Huánuco, limitada mayormente a la ecorregión de selva baja y con elevaciones de 1000 m a 2200 m (MINAG 2003, Vera et al. 2014). El bosque de Carpish, en cambio, tiene una extensión de 1240.25 km2 (GORE Huánuco 2016) y un rango de elevación de 1780 m a 3080 m, que al ser protegido o conservado representaría adecuadamente la biodiversidad de las yungas de la selva central.
Carpish tiene aún extensos bosques con una alta diversidad por lo que sería una buena opción declararla como una nueva área de conservación, sin embargo aún no cuenta con estudios detallados de su biodiversidad, particularmente en relación a la altitud, topografía y dinámica del bosque, información importante para gestión y su conservación (Young & León 1999, Tovar et al. 2010).
Actualmente, Carpish no está incluido en el Sistema Nacional de Áreas Naturales Protegidas por el Estado (SINANPE), pero el Gobierno Regional de Huánuco aprobó la propuesta de Área de Conservación Regional Bosque Montano de Carpish (GORE Huánuco 2016), aunque con un sustento de biodiversidad incompleto.
El presente estudio documenta la diversidad y composición de mamíferos en los bosques montanos de Carpish, en base a expediciones realizadas durante el 2001 y 2002, con los objetivos de: 1) Estimar su diversidad alfa para el área de estudio, 2) determinar los cambios en índices de diversidad y abundancia relativa en una gradiente de elevación y 3) sustentar la conservación del bosque de Carpish basada en los criterios de diversidad, grado de endemismo y presencia de especies amenazadas de mamíferos.
Material y métodos
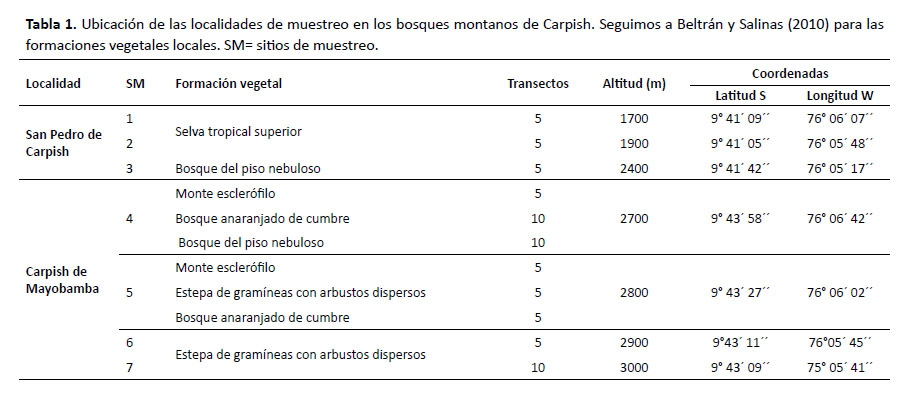
La Cordillera Carpish se ubica en el distrito de Chinchao, provincia de Huánuco, Región de Huánuco, entre las ciudades de Huánuco y Tingo María. Para el presente trabajo se realizaron evaluaciones a diferentes elevaciones desde la parte más baja (1780 m) hasta las zonas más altas (3080 m) (Fig. 1). Se realizaron cinco expediciones a la localidad de Carpish de Mayobamba entre los meses de mayo y noviembre del 2001 (aproximadamente 16 días) y en febrero, mayo y agosto del 2002 (aproximadamente 41 días). En la quinta expedición (agosto 2002) se evaluó también la localidad San Pedro de Carpish (aproximadamente 10 días). Los datos de coordenadas y elevación se especifican en la Tabla 1.
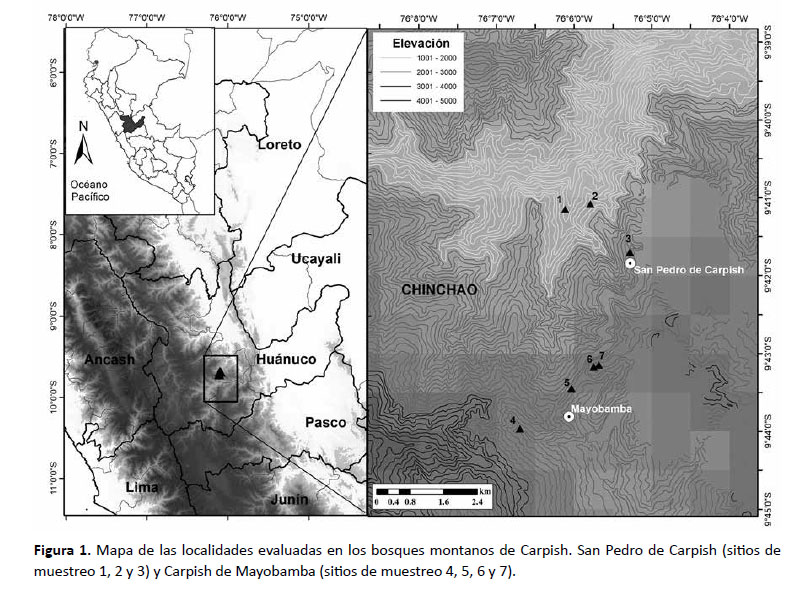
Seguimos a Beltrán y Salinas (2010) para la descripción de las formaciones vegetales locales.
Carpish de Mayobamba (2730 – 3080 m) .Corresponde a las zonas más altas y mejor drenadas del conjunto montañoso de Carpish y donde se establecieron los sitios de muestreo 4, 5, 6 y 7. Incluye el hábitat de bosque montano alto con las formaciones vegetales:
– Estepa de gramíneas con arbustos dispersos (2800 – 2950 m), el suelo es de consistencia esponjosa, formado por el profuso crecimiento de Sphagnum sp. y descomposición de la hojarasca. La vegetación es más bien leñosa, compuesta por ericáceas pequeñas, que ocurren en las pequeñas depresiones que se forman por lo irregular de la superficie del terreno.
– Turberas de musgo con dispersos arbustos ericoides (2790 – 2800 m), es una transición entre la estepa de gramíneas y el monte esclerófilo. La vegetación está conformada por elementos de ambas formaciones, además de plantas trepadoras, parásitas e insectívoras, musgos, líquenes, helechos y hierbas.
– Monte esclerófilo (2700 – 2850 m), una de las formaciones vegetales más importantes por su predominancia, ocupando pendientes y colinas. La vegetación es del tipo de bosque enano que alcanza unos 3 m de alto, muy denso que hace difícil ver el suelo o roca madre.
– Bosque anaranjado de cumbre (2700 – 2900 m), esta formación vegetal es reconocida sólo por Foster y Beltrán (1997) y es un ecotono entre el matorral esclerófilo y el bosque nebuloso. El dosel alcanza los 7m de alto, y la vegetación estaría asociada a un substrato de naturaleza ácida.
San Pedro de Carpish (1780 – 2400 m). Corresponde a las zonas más bajas de la Cordillera, la cual está generalmente cubierta de nubes y donde se establecieron los sitios de muestreo 1, 2 y 3. Incluye el hábitat de bosque montano bajo con las formaciones vegetales:
– Bosque del piso nebuloso (2300 – 2750 m), es la más dominante en Carpish y donde se desarrolla gran parte de la diversidad arbórea de todo el bosque. Se extiende hacia las partes altas de Carpish de Mayobamba. El dosel alcanza entre los 10 a 15 m de alto; la vegetación incluye helechos arborescentes, alta diversidad de epífitas (orquídeas, aroideas, bromelias, helechos, líquenes y musgos) que llegan a cubrir casi en su totalidad las ramas y troncos de los árboles. Es también una zona medianamente perturbada por la acción del hombre.
– Selva tropical superior (1700 – 2300 m), es la formación vegetal donde el cambio de bosque montano alto a bajo es abrupto teniendo pocas especies en común. El dosel alcanza de 30 a 35 m de alto y hay escaso desarrollo del sotobosque. La vegetación incluye lianas y epífitos (helechos, bromelias, ciclantáceas, aroideas); el sotobosque contiene arbustos dominantes de Miconia, Palicourea, Piper y Psychotria. Es la formación vegetal más impactada por la actividad humana.
Para esta evaluación se usaron trampas y redes siguiendo la metodología detallada en Pacheco et al. (2007). Se emplearon trampas de tipo golpe, de caja y pitfall, además de redes de niebla. Las trampas fueron instaladas en transectos de 30 estaciones en un hábitat relativamente homogéneo, con un par de trampas por estación, y separadas por aproximadamente 10 m entre estaciones de trampeo. El cebo consistió en una mixtura de avena, mantequilla de maní, vainilla, miel, pasas y diferentes semillas para canarios.
Se emplearon redes de niebla de 12 m o 6 m de largo por 2.5 m para la evaluación de murciélagos, instaladas en diferentes tipos de hábitats. En total se evaluaron 70 transectos para la captura de roedores y marsupiales y 15 redes para murciélagos; el esfuerzo de muestreo total fue de 28500 trampas noche y 190 redes noche (Tabla 2).
Seguimos los lineamientos éticos de la American Society of Mammalogists para la eutanasia de los animales (Sikes et al. 2016). Los especímenes se encuentran depositados en el Departamento de Mastozoología del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos, Lima, Perú (MUSM).
La presencia de mamíferos de mayor tamaño se evaluó a través de entrevistas y registros directos (observaciones, vocalizaciones, etc.) e indirectos (huellas, heces, madrigueras, etc.). Las entrevistas se realizaron con un modelo de ayuda de preguntas preestablecidas además se mostraron imágenes de especies probables en el área (Emmons & Feer 1997, Pacheco et al. 2007). Las observaciones fueron realizadas ad libitum durante la instalación o la revisión de los transectos de mamíferos pequeños, o durante el desplazamiento entre localidades; sin embargo, para los análisis cuantitativos no fueron incluidas las especies registradas por observación o entrevista. Se siguió la nomenclatura taxonómica y nombres comunes empleados en Pacheco et al. (2009), Gardner (2008), Patton et al. (2015) y literatura actualizada.
Se construyó la curva de acumulación de especies para visualizar la tendencia y representatividad del esfuerzo de muestreo empleado (Colwell & Coddington 1994) para mamíferos no voladores y murciélagos; para ello se aleatorizaron los muestreos utilizando los días como unidad de muestreo. Se empleó EstimateS (Colwell 2006) para calcular los índices y Statistica (StatSoft 2011) para graficar la curva.
La abundancia relativa (AR) de las especies fue estimada en base al esfuerzo de captura empleado expresado como trampas noche (TN) para roedores y marsupiales y expresado en redes noche (RN) para murciélagos; así, la AR del primer grupo se expresa como el número de individuos capturados por cada 100 TN y para el segundo grupo, el número de individuos capturados por cada 10 RN (Tabla 2).
La diversidad fue estimada en relación a la riqueza de especies, los índices de Shannon-Wiener, de Simpson y equitatividad (Krebs 1989). Para evaluar la relación entre la abundancia relativa y la diversidad de mamíferos con respecto a la elevación se empleó el coeficiente de correlación de Pearson.
Finalmente, el estado de conservación de las especies fue evaluada considerando la legislación vigente (MINAGRI 2014), la Unión Internacional para la Conservación de la Naturaleza (IUCN 2018) y la Convención Internacional sobre el Comercio de Especies Amenazadas de Fauna y Flora (CITES 2017).
Resultados
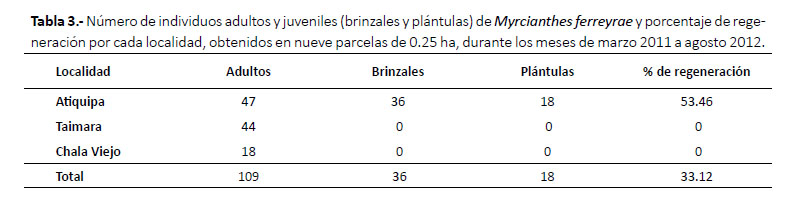
Se registraron 897 especímenes y cuatro observaciones de mamíferos medianos a grandes, lo que resulta en la presencia de 35 especies de mamíferos nativos en los bosques montanos de Carpish. Estos incluyen 5 didelfimorfios, 1 primate, 11 roedores, 15 murciélagos, 2 carnívoros y 1 artiodáctilo (Tabla 3, Apéndice 1).
Los murciélagos con 15 especies (42.9%) y los roedores con 11 especies (31.4%) resultaron los órdenes más diversos y dentro de ellos, las familias Phyllostomidae (34.3%) y Cricetidae (22.9 %), respectivamente, fueron las mejor representadas. Por otro lado, los roedores se agruparon en cuatro familias, siendo así los más diversos. Se sigue aquí a Jiménez y Pacheco (2016) al reconocer a Akodon kotosh como especie válida distribuida en las localidades evaluadas. Entre los murciélagos, los géneros Anoura y Sturnira fueron los más diversos (Apéndice 1).
Algunas de las especies registradas constituyen nuevos registros regionales o proporcionan nueva información sobre la distribución de la especie; por ejemplo, los ejemplares de Monodelphis gardneri, capturados en San Pedro de Carpish son los primeros registros para el departamento de Huánuco (Solari et al. 2012). El marsupial Marmosops impavidus fue capturado en las dos localidades. Los marsupiales Gracilinanus aceramarcae y Marmosops bishopi son especies muy raras y fueron capturadas en Carpish de Mayobamba y San Pedro de Carpish respectivamente. Los roedores Thomasomys aureus y Thomasomys notatus, capturados en Carpish de Mayobamba, resultan ser los primeros registros para el bosque de Carpish. Los murciélagos Eptesicus andinus,
La correlación del número de especies de roedores con la elevación no fue estadísticamente significativa para roedores (r= -0.37; p= 0.46) ni para murciélagos (r= -0.80; p= 0.06).
En Carpish de Mayobamba se registró la mayor riqueza con 28 especies a 2700 m, predominando murciélagos y roedores; mientras que en San Pedro de Carpish, la mayor riqueza (13 especies) fue registrada a 1900 m (Apéndice 1, Tabla 3).
Otros registros importantes incluyen, el roedor Akodon kotosh, endémico de Carpish. También, se registran especies endémicas para Perú como Thomasomys ischyrus, T. kalinowskii y T. notatus (Pacheco et al. 2009). Encontramos también que en San Pedro de Carpish se encuentran especies no compartidas con Carpish de Mayobamba (i.e., Carollia brevicauda, Desmodus rotundus, Marmosops bishopi, Monodelphis gardneri); mientras que lo inverso también ocurre con otras especies (i.e., Eptesicus andinus, Myotis keaysi, Thomasomys aureus). Probablemente, con mayor esfuerzo de muestreo, estas especies exhiban un mayor rango de distribución.
El número de especies registradas para mamíferos no voladores (Fig. 2) significó el 92.02% de las especies esperadas (Sesp= 13.04), lo que describe un muestreo bastante completo con los métodos de captura de roedores y marsupiales; mientras que el número de especies registradas para murciélagos (Fig. 3) significó el 80.60% de las especies esperadas (Sesp= 18.61), lo que describe un muestreo bastante bueno con los métodos de captura empleados para murciélagos.
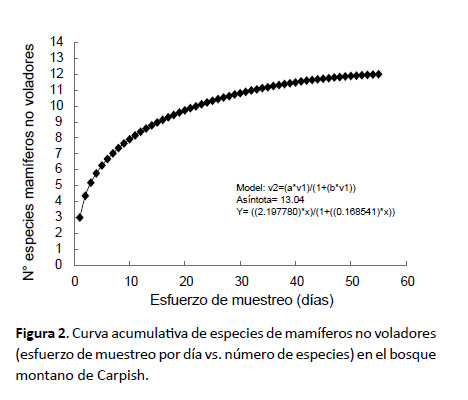
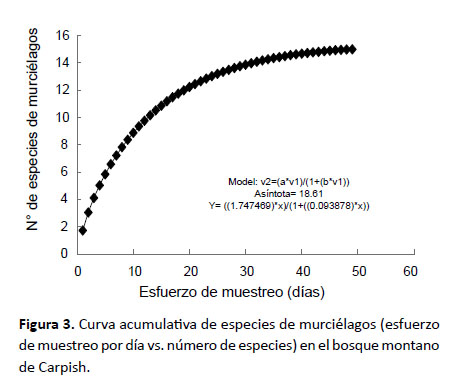
En relación con la estacionalidad, durante la época húmeda, se registraron los murciélagos Eptesicus andinus, Myotis keaysi, Platyrrhinus nigellus y Sturnira oporaphilum; los mismos que en época seca estuvieron ausentes, ocurriendo más bien Carollia brevicauda y Desmodus rotundus. Los roedores thomasominos y akodontinos no siguen un patrón estacional de distribución ya que estuvieron presentes en ambas estaciones. Los marsupiales Marmosops bishopi y Monodelphis gardneri fueron registrados sólo en época seca en las zonas más bajas del bosque de Carpish (1785 m). Es posible que con mayor esfuerzo de muestreo estas diferencias de presencia estacional disminuyan.
Abundancia relativa
La abundancia relativa (AR) es analizada para el grupo de roedores y marsupiales (Tabla 4) y murciélagos en Carpish de Mayobamba, a 2900 m, se registró la mayor AR; mientras que en San Pedro de Carpish, a 1700 m, se registró uno de los valores de AR más bajos. Para los murciélagos, en San Pedro de Carpish (1900 m) se regis- (2700 m) se registró uno de los valores AR más bajos; en este caso las AR más altas estarían relacionadas con hábitats de elevaciones bajas. El análisis de correlación para ambos grupos evidencia falta de correlación con la elevación.
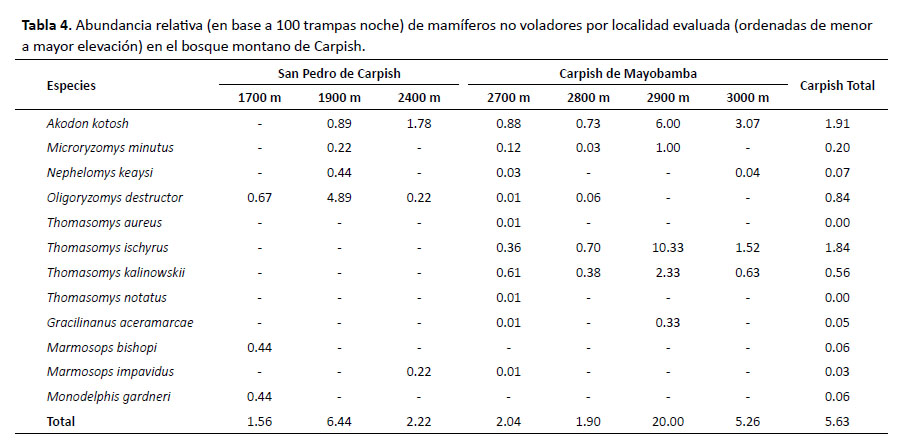
En Carpish de Mayobamba, Thomasomys ischyrus fue la especie más abundante, mientras que Oligoryzomys destructor lo fue en San Pedro de Carpish (Tabla 4).
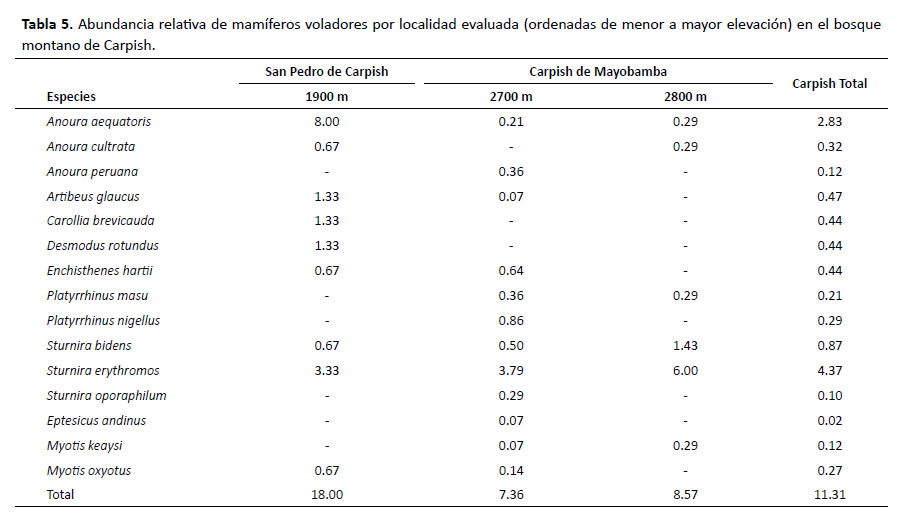
Por otro lado, en San Pedro de Carpish, Anoura aequatoris fue la especie más abundante y la que registró la más alta AR en toda el área evaluada; en Carpish de Mayobamba, Sturnira erythromos fue las más abundante (Tabla 5).
En Carpish de Mayobamba fueron avistados el mono coto Alouatta seniculus, el marsupial Didelphis pernigra y el mustélido Eira barbara. Mediante entrevistas se registraron los roedores Cuniculus taczanowskii, Dasyprocta sp. y el venado "sachacabra" Pudu mephistophiles. La presencia de este pequeño venado fue mencionada por Grimwood (1969) y confirmada por Pedro Hocking (comm. pers.). Este último lo observó en el año 1970 y colectó un ejemplar en la localidad cercana del Bosque Zapatogocha, en Acomayo (capital del distrito de Chinchao) en 1972 y refiere que actualmente es posible encontrar sus caminos en las partes más altas del bosque de Carpish.
En San Pedro de Carpish se registraron avistamientos de ardillas de Hadrosciurus sp.; la especie puede corresponder a un voucher del Louisiana State University, Museum of Zoology representando a Hadrosciurus pyrrhinus, colectado en Carpish (LSUMZ 18437).
Índices de diversidad
Los valores obtenidos con los índices de ShannonWiener para mamíferos no voladores (Tabla 6) muestran la mayor diversidad en Carpish de Mayobamba a 2700 m (H'= 1.38) y la menor en San Pedro de Carpish a 2400 m (H'= 0.64). Por otro lado, con el índice de Simpson, se registra la diversidad más baja en localidades intermedias como San Pedro de Carpish a 1900 m y 2400 m (1 – D= 0.40 y 1 – D= 0.34 respectivamente). El análisis del coeficiente de correlación de Pearson mostró que no existe una correlación significativa de los valores de H' y 1 – D con respecto a la elevación. Por otro lado, la equidad (J') fue más alta en San Pedro de Carpish (1700 m) lo cual indica que este lugar está cerca de alcanzar su máxima diversidad.
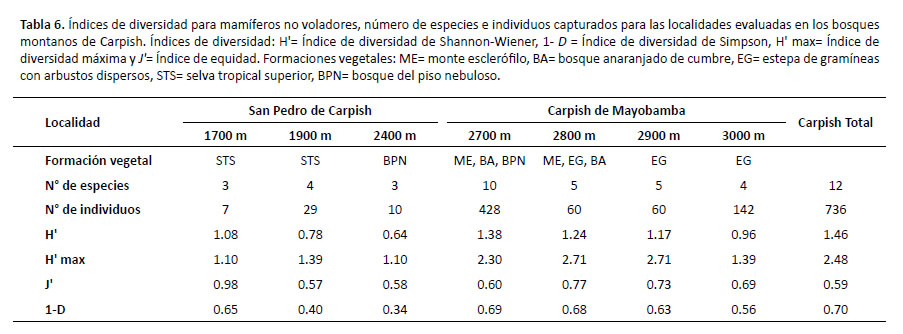
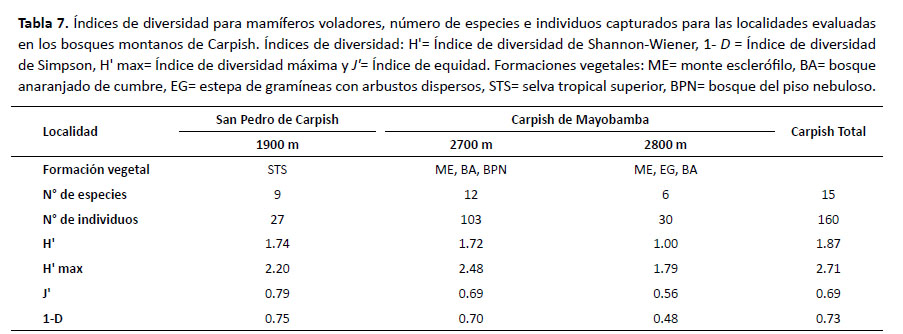
Para mamíferos voladores se observó que en San Pedro de Carpish (1900 m) se registró la mayor diversidad (H'= 1.74) y el valor más alto (1 – D= 0.75) con el índice de Simpson; mientras que la menor diversidad (H'= 1.00) se obtuvo en Carpish de Mayobamba (2800 m) así como el valor más bajo (1 – D= 0.48) con el índice de dominancia de Simpson. En relación al índice de equidad (J') se observa que fue moderadamente alta en San Pedro de Carpish (1900 m). Para mamíferos voladores no se aplicó el coeficiente de correlación de Pearson debido a que no se muestreó en todas las elevaciones (Tabla 7).
Especies de interés en conservación
Nuestros resultados muestran cuatro especies de mamíferos que están registradas como amenazadas en la legislación peruana (MINAGRI 2014): Alouatta seniculus, Thomasomys ischyrus, T. kalinowskii y Pudu mephistophiles; todas ellas en la categoría "vulnerable"; adicionalmente, P. mephistophiles se encuentra registrada también como "vulnerable"en la lista CITES (2017) y en la de la IUCN (2018). Por otro lado, los roedores Akodon kotosh y T. notatus, si bien no están bajo ninguna categoría de amenaza, son endémicos para el país. Así mismo, T. ischyrus y T. kalinowskii aunque son relativamente comunes localmente, son endémicos de Perú.
Discusión
Los bosques montanos centrales no cuentan con estudios detallados de diversidad de mamíferos, los cuales son importantes para sustentar los esfuerzos de conservación en la zona. Los resultados del presente trabajo apoyan la necesidad de conservar este bosque como un área representativa de los bosques montanos o yungas del centro del Perú, ecosistemas que están siendo destruidos o fragmentados rápidamente, sin que existan alternativas viables para su conservación, motivo por el que son consideradas una de las 18 ecorregiones terrestres prioritarias a nivel mundial con respecto a conservación (Mittermeier et al. 1999).
La importancia biogeográfica de Carpish se demuestra por los nuevos registros de este estudio; el marsupial Monodelphis gardneri, los roedores Thomasomys aureus y T. notatus. Así mismo, es importante destacar los registros de Gracilinanus aceramarcae, Marmosops bishopi, Eptesicus andinus, Anoura cultrata y Artibeus glaucus, especies consideradas raras por la literatura (Pacheco et al. 2009, Gardner 2008, Patton et al. 2015), pero que fueron registradas en Carpish.
Aunque Carpish de Mayobamba (2730 m a 3080 m) muestra en general una mayor riqueza de especies de mamíferos comparado a San Pedro de Carpish (1780 m a 2400 m), aún es prematuro sugerir la presencia de un patrón relacionado a la elevación como fue encontrado en otras Yungas peruanas (Pacheco et al. 2011), ya que también puede deberse a la mayor deforestación en las partes bajas.
Concordando con Pacheco (2002), en Carpish los roedores y murciélagos (74.3%) son los grupos más diversos, siendo mayor la riqueza de especies de murciélagos filostómidos, lo cual estuvo relacionado al método de captura con redes a nivel de sotobosque, por lo que se recomienda el uso de métodos adicionales como redes de dosel, búsqueda de refugios y grabaciones de ecolocación, para complementar la diversidad de especies de murciélagos (Voss & Emmons 1996).
Según los índices de H' y 1 – D, la diversidad es mayor en San Pedro de Carpish para murciélagos, mientras que en Carpish de Mayobamba es mayor para los roedores. Por lo tanto, en general ambas localidades son ricas y diversas. Los valores bajos para murciélagos en Carpish de Mayobamba pudieron deberse a que no se evaluó toda la gradiente, además que algunas evaluaciones coincidieron con los días de luna llena, días no recomendados para la captura de murciélagos (Flores 2008) ya que éstos prefieren las noches oscuras debido a que presentan un fenómeno conocido como fobia lunar (Fenton 1991).
Aunque, Pacheco et al. (2009) señalaron la presencia de Gracilinanus aceramarcae a lo largo de las Yungas, esta era solo conocida para Puno, Cusco, Piura y San Martín; en el presente estudio se amplía la distribución de la especie para Huánuco entre los 2770 y 2920 m.
Los murciélagos pequeños como Sturnira, por lo general, tienden a prosperar en áreas perturbadas (Montaño et al. 2015). Este es el caso en Carpish, debido a que registramos alta diversidad y abundancia de ese género.
Nuestros resultados indican que aún existe continuidad de bosque desde las zonas bajas a altas, dado por las especies registradas a lo largo de la gradiente evaluada en Carpish, por ejemplo A. kotosh y S. erythromos. La conservación de Carpish debería por lo tanto incluir una gradiente de elevación que favorezca la dispersión de especies.
Se concluye que los bosques montanos de Carpish albergan al menos 45 especies de mamíferos, que resulta de incluir las 35 especies aquí registradas y las especies en reportes previos por Gardner y Carter (1972), Wetzel (1985), Jenkins y Barnett (1997) y Sunquist y Sunquist (2009). Se observa que esta diversidad es mayor a las 17 especies registradas en un rango de 1800 a 2750 m en la cuenca del río Apurímac (Pacheco et al. 2007), probablemente por el mayor esfuerzo de captura empleado en Carpish.
Se recomienda desarrollar una zonificación ecológica y un plan de conservación in situ para el bosque de Carpish, el cual involucre a los pobladores del área, autoridades e investigadores, con el objetivo de favorecer el desarrollo de estos grupos de interés y garantizar la conservación y mejor gestión del bosque.
Agradecimientos:
Muchas personas participaron en el trabajo de campo, pero merecen especial reconocimieto por su participación y esfuerzo en el campo: Heidi Quintana, Alicia Vásquez, Marina Villalobos, Dyana La Rosa, Paul Velazco y Lili Arias. Nuestros agradecimientos a Richard Cadenillas por su asesoría en el aspecto estadístico y a Pamela Sánchez-Vendizú por la elaboración del mapa de los sitios de muestreo. Igualmente, nuestro agradecimiento a Diana Silva por sus valiosos comentarios. Al Ministerio de Agricultura y Riego por facilitarnos las autorizaciones correspondientes. Esta investigación fue financiada por CONCYTEC con el fondo concursable "Santiago Antúnez de Mayolo" 2001 Proyecto 078 "Protección de la biodiversidad en bosques montanos fragmentados y propuesta para conservar el bosque de Carpish, Huánuco".
Rol de los autores:
VP que diseñó la investigación, colectó los especímenes, escribió y revisó críticamente el manuscrito y consiguió el financiamiento para el estudio. MN que realizó los análisis estadísticos y participó en el diseño y preparación del manuscrito.
Conflicto de intereses:
Los autores no incurren en conflictos de intereses.
Fuentes de financiamiento:
Esta investigación fue financiada por CONCYTEC con el fondo concursable "Santiago Antúnez de Mayolo" 2001 Proyecto 078 "Protección de la biodiversidad en bosques montanos fragmentados y propuesta para conservar el bosque de Carpish, Huánuco".
Aspectos éticos / legales:
Seguimos los lineamientos éticos de la American Society of Mammalo- gists para la eutanasia de los animales (Sikes et al. 2016).
Con autorización No 013-2001-INRENA-J-DGAPFS. Servivio Nacional Forestal y de Fauna Silvestre (SERFOR) del Ministerio de Agricultura y Riego permisos de recolección No. 084-2012-AG-DGFFS-DGEFFS 0148-2013-AG-DGFFS-DGEFFS
Citación:
Pacheco V. & M. Noblecilla. 2019. Diversidad de mamíferos en el bosque montano de Carpish, Huánuco, Perú. Revista peruana de biología 26(2): 217 226 (Julio 2019). doi: http://dx.doi.org/10.15381/rpb.v26i2.16372
Literatura citada
Beltrán H. & I. Salinas. 2010. Flora vascular y vegetación de los Bosques Montanos Húmedos de Carpish (Huánuco Perú). Arnaldoa 17 (1): 107 130. ISSN: 1815 – 8242. [ Links ]
Colwell R.K. & J.A. Coddington. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London B 345: 101– 118. https://doi.org/10.1098/rstb.1994.0091 [ Links ]
Colwell R.K. 2006. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User's Guide), Version 8. [ Links ]
CITES 2017. (en línea). Apéndices I, II y III. <www.cites.org/ esp/app/appendices.php>. Acceso 12/10/18. [ Links ]
Emmons L.H. & F. Feer. 1997. Neotropical rainforest mammals, a field guide. 2nd edn., The University of Chicago Press, Chicago. Pp. 307. https://doi.org/10.2307/1383232 [ Links ]
Fenton M. B. 1991. Just bats. University of Toronto Press, Toronto, Buffalo. Pp. 165. [ Links ]
Flores M. 2008. Estructura de las comunidades de murciélagos en un gradiente ambiental en la reserva de la biosfera y tierra comunitaria de origen Pilón Lajas, Bolivia. Mastozoología Neotropical 15(2): 309-322. ISSN 0327-9383 [ Links ]
Foster R. & H. Beltrán.1997. Vegetación y flora de la Cordillera del Cóndor. In T. S. Schulenberg & K. Awbrey (eds.) The Cordillera del Cóndor region of Ecuador and Peru: A biological assessment. Conservation International Rapid Assessment Program Working Papers 7: 45-54 [ Links ]
Gardner A.L. 1971. Karyotypes of two rodents from Peru, with a description of the highest diploid number recorded for a mammal. Experientia 26: 1088-1089. https://doi.org/10.1007/BF02138898 [ Links ]
Gardner A.L. & D.C. Carter. 1972. A new stenodermine bat (Phyllostomatidae) from Peru. Occas. Papers Mus., Texas Tech Univ. 2:1–4. https://doi.org/10.5962/bhl.title.156587. [ Links ]
Gardner A.L. & J.L. Patton. 1976. Karyotypic variation in oryzomyine rodents (Cricetinae) with comments on chromosomal evolution in the Neotropical cricetine complex. Occasional papers of the Museum of Zoology. Louisiana State University 49: 1-48. [ Links ]
Gardner A.L. (editor). 2008 (2007). Mammals of South America, Volume 1. Marsupials, Xenarthrans, Shrews, and Bats. United States of America: The University of Chicago Press. doi.org/10.7208/chicago/9780226282428.001.0001. [ Links ]
GORE Huánuco. 2016. Aprueban la propuesta del área de conservación regional "Bosque Montano de Carpish" y su expediente técnico. Ordenanza Regional Nº 0472016-CR-GRH. 19 de junio de 2016. El Peruano Normas Legales: 590035-590038. [ Links ]
Grimwood, I.R. 1969. Notes on the distribution and status of some Peruvian mammals 1968. Special Publication, American Committee for International Wild Life Protection 21:1-86. [ Links ]
IUCN 2018. (en línea). IUCN Red list of threatened species. Version 2018.1. <www.iucnredlist.org>. Acceso 03/11/2018. [ Links ]
Jenkins P.D. & A.A. Barnett. 1997. A new species of water mouse, of the genus Chibchanomys (Rodentia, Muridae, Sigmodontinae) from Ecuador. Bulletin of The Natural History Museum, London (Zoology) 63:123-28. [ Links ]
Jiménez C.F. & V. Pacheco. 2016. A new species of grass mouse, genus Akodon Meyen, 1833 (Rodentia, Sigmodontinae), from the central Peruvian Yungas. Therya 7(3): 449-464. https://doi.org/10.12933/therya-16-336 [ Links ]
Krebs C. J. 1989. Ecological methodology. Harper Collins. New York. 654pp. [ Links ]
Lehr, E. & D. Rodríguez. 2017. Two new species of Andes Frogs (Craugastoridae: Phrynopus) from the Cordillera de Carpish in central Perú. Salamandra 53(3): 327-338. [ Links ]
MINAG (Ministerio de Agricultura). 2003. Plan Maestro del Parque Nacional Cordillera Azul 2003 2008. Lima-Perú. 273 pp. [ Links ]
MINAM (Ministerio del Ambiente). 2012a. Plan Maestro del Santuario Nacional Pampa Hermosa 2012 2017. Lima-Perú. 55 pp. [ Links ]
MINAM (Ministerio del Ambiente). 2012b. Plan Maestro del Parque Nacional Tingo María 2012 2017. Lima-Perú. 54 Pp. [ Links ]
MINAGRI (Ministerio de Agricultura y Riego). 2014. Decreto Supremo Nº 004-2014-MINAGRI. El Peruano Normas Legales: 520497 520504. [ Links ]
Mittermeier R.A., N. Myers, P. Robles Gil & C.G. Mittermeier. 1999. Biodiversidad Amenazada. Las Ecorregiones terrestres prioritarias del Mundo. CEMEX, México, 431 pp. [ Links ]
Montaño-Centellas, F. M. I. Moya, L. F. Aguirre, C. R. Galeón, O. Palabral, R. Hurtado, I. Galarza y J. Tordoya. 2015. Community and species-level responses of phyllostomid bats to a disturbance gradient in the tropical Andes. Acta Oecologica 62: 10-17. https://doi.org/10.1016/j.actao.2014.11.002. [ Links ]
Pacheco V. 2002. Diversidad y Conservación de los Mamíferos Peruanos. En Ecología y Conservación de los mamíferos de Latinoamérica. (G. Ceballos y J. Simonetti, Eds). Fondo de Cultura Económica, México, D. F. [ Links ]
Pacheco V., B. Inche & W. Wust. 2018. Mamíferos del Perú. Grupo La República Publicaciones S.A., Lima. 120 pp. [ Links ]
Pacheco V., E. Salas, L. Cairampoma, et al. 2007. Contribución al conocimiento de la diversidad y conservación de los mamíferos en la cuenca del río Apurímac, Perú. Revista Peruana de Biología 14(2), 169-180. doi:10.15381/rpb.v14i2.1722. [ Links ]
Pacheco V., R. Cadenillas, E. Salas, et al. 2009. Diversidad y endemismo de los mamíferos del Perú. Revista Peruana de Biología 16(1): 5-32. https://doi.org/10.15381/rpb.v16i1.111. [ Links ]
Pacheco V., G. Márquez, E. Salas & O. Centty. 2011. Diversidad de mamíferos en la cuenca media del río Tambopata, Puno, Perú. Revista Peruana de Biología 18(2): 231 – 244. https://doi.org/10.15381/rpb.v18i2.246. [ Links ]
Parker, T.A. & J.P. O’Neill. 1976. Introduction to bird finding in Perú: Part II. The Carpish Pass region of the Eastern Andes along the central highway. Birding 20: 205–216.
Patton J.L., U. Pardiñas, & G. D’Elía .2015. Mammals of South America, Volume 2. Rodents. United States of America: The University of Chicago Press. https://doi. org/10.7208/chicago/9780226169606.001.0001.
Sikes, R.S., & The Animal Care and Use Committee of the American Society Of Mammalogists. 2016. 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. Journal of Mammalogy 97(3): 663–688. https://doi.org/10.1093/jmammal/gyw078. [ Links ]
Solari S., V. Pacheco, E. Vivar & L.H. Emmons. 2012. A new species of Monodelphis (Mammalia: Didelphimorphia: Didelphidae) from the montane forests of central Perú. Proceedings of the Biological Society of Washington 125(3): 295–307. https://doi.org/10.2988/11-33.1 [ Links ]
StatSoft Inc. 2011. Statistica (Data analytic tool). Versión 10. [ Links ] Sunquist M.E. & F.C. Sunquist. 2009. Familia Felidae (Cats).
In: D. E. Wilson y R. A. Mittermeier, eds. Handbook of the mammals of the world. Vol. 1. Carnivores. Lynx Edicions. Barcelona. Pp. 146-147. https://doi.org/10.1007/bf03193176.
Tovar A. (editor). 2010. Yungas Peruanas Bosques montanos de la vertiente oriental de los Andes del Perú: Una perspectiva ecorregional de conservación. Lima. Centro de Datos para la Conservación de la Universidad Nacional Agraria La Molina. [ Links ]
Vera A. (editor). 2014. Parque Nacional Cordillera Azul Joya Natural para la Humanidad. Lima. Centro de Conservación, Investigación y Manejo de Áreas Naturales – Cordillera Azul (CIMA-Cordillera Azul). [ Links ]
Voss R.S. & L. H. Emmons. 1996. Mammalian diversity in Neotropical Lowland rainforests: A preliminary assessment. Bulletin of the American Museum of Natural History 230: 18-22. [ Links ]
Wetzel, R.M. 1985. Taxonomy and distribution of armadillos, Dasypodidae. In: G. G. Montgomery, ed. The evolution and ecology of armadillos, sloths, and vermilinguas. Pp. 23–48. Washington, DC: The Smithsonian Institution Press, 10 (unnumbered) +451 pp. [ Links ]
Young K. & B. León. 1999. Peru’s humid eastern montane forests. DIVA, Technical report 5: 1-97.
Correspondencia:
* Autor para correspondencia
Víctor Pacheco: vpachecot@unmsm.edu.pe
Maggie Noblecilla: mcnoblecilla@gmail.com
Presentado: 31/12/2018
Aceptado: 21/02/2019
Publicado online: 06/07/2019

