











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
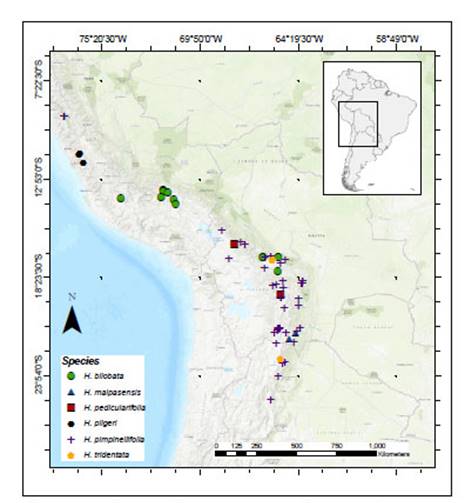
Hypseocharis J. Rémy is a genus endemic to middle to high elevations of the Andes (ca. 2000-4200 m) ranging from Peru to Argentina (Figure 1). In Bolivia it occurs along the eastern Cordillera of the Andes in the Puna, and Pre-puna regions, and in dry valleys between 2500 and 3800 m. It is usually found in open habitats, often on shallow soils in Andean grasslands and between rocks.
The genus was first described by Rémy (1847) and placed in the family Geraniaceae. Knuth (1930) recognized three genera within Oxalidaceae R. Br: Oxalis L., Biophytum DC and Hypseocharis, and he differentiated them based on habit, leaf morphology, the number of stamens and style morphology. Systematics of Hypseocharis have been controversial. Cronquist (1981) considered the genus as a link between the Oxalidaceae and Geraniaceae, while Boesewinkel (1988) analyzed the seed structure and affirmed that the genus is closer related to Geraniaceae than to Oxalidaceae. Subsequent studies on floral anatomy demonstrated that Hypseocharis should be placed in Geraniaceae (Devi, 1991). Price and Palmer (1993) confirmed that placement with molecular data (rbcL-phylogeny) and Palazzesi et al. (2012) retrieved Hypseocharis as sister to core-Geraniaceae and suggested resurrecting the family Hypseocharitaceae Wedd. (Weddell in 1857), based on both the phylogenetic position and the aberrant morphology (especially the capsular fruit).
Currently, six species are recognized in the genus according to the Tropicos website (Missouri Botanical Gardens): Hypseocharis bilobata Killip, H. malpasensis R. Knuth, H. pedicularifolia R. Knuth, H. pimpinellifolia J. Rémy, H. pilgeri R. Knuth, and H. tridentata Griseb. while H. corydalifolia, R. Knuth, H. fiebrigii R. Knuth , H. moschata R. Knuth and H. pimpinellifolia var. punicea Kuntze are considered as synonyms (http://www.tropicos.org, accessed 01.08. 2019). All six accepted species, apart from H. pilgeri, are reported in the Checklist of the Flora of Bolivia (Jørgensen et al. 2014). Macbride (1949) reported four species from Peru: H. bilobata, H. pedicularifolia, H. pilgeri and H. tridentata mainly based on leaflets morphology and number of flowers per peduncle. These authors did not consider the number of stamens, flower color or type of fruit to distinguish the four species.
Slanis and Grau (2001) provided the only recent, critical revision of the genus, focusing on material from Argentina. They proposed the recognition of only two species, namely H. pimpinellifolia and H. tridentata not only considering leaf morphology, but also including number of stamens and type of fruit. Slanis and Grau (2001) also emphasized that the taxonomy of the species in Bolivia and Peru requires critical work and voiced their suspicion that species limits as currently accepted are incorrect. The aim of the present study is to critically revise the genus for the area not covered by Slanis and Grau (2001), i.e., Bolivia and Peru.
Material and methods
A total of 105 specimens mainly from the Herbario Nacional de Bolivia (La Paz, LPB), the Herbario de la Universidad Nacional Mayor de San Marcos (Lima, USM), Field Museum of Natural History (Chicago, F) and Muséum National d'Histoire Naturelle (France, P) were critically revised. Additional material was seen in BOLV, HUT, MO, P, US, BONN, F, B, K, GH, NHA, CORD, SI, LIL (herbarium acronyms follow Thiers 2019). Seeds collected in Bolivia (Ortuño 1361) were germinated in Bonn University Botanic Gardens and taken into cultivation and a population of the putative Peruvian endemic "H. pilgeri" (Peru, Department Lima) was studied in the field.
Vegetative and reproductive morphological characters were qualified and quantified from a subset of specimens from Bolivia representing all character combination used to delimit putative species based on key from Flora of Peru (Macbride, 1949) and classification key of Hypseocharis in Argentina (Slanis & Grau, 2001). Specimens were assigned to taxa based on the technical characters used in the literature and based on a comparison to type specimens, where available. Hypseocharis is generally poorly represented in herbaria, probably due to its very short flowering time, high elevation habitat and the technical problems of preparing good specimens due to the rosulate habit and very deep-seated roots. Only specimens showing all relevant characters were included in the analysis.
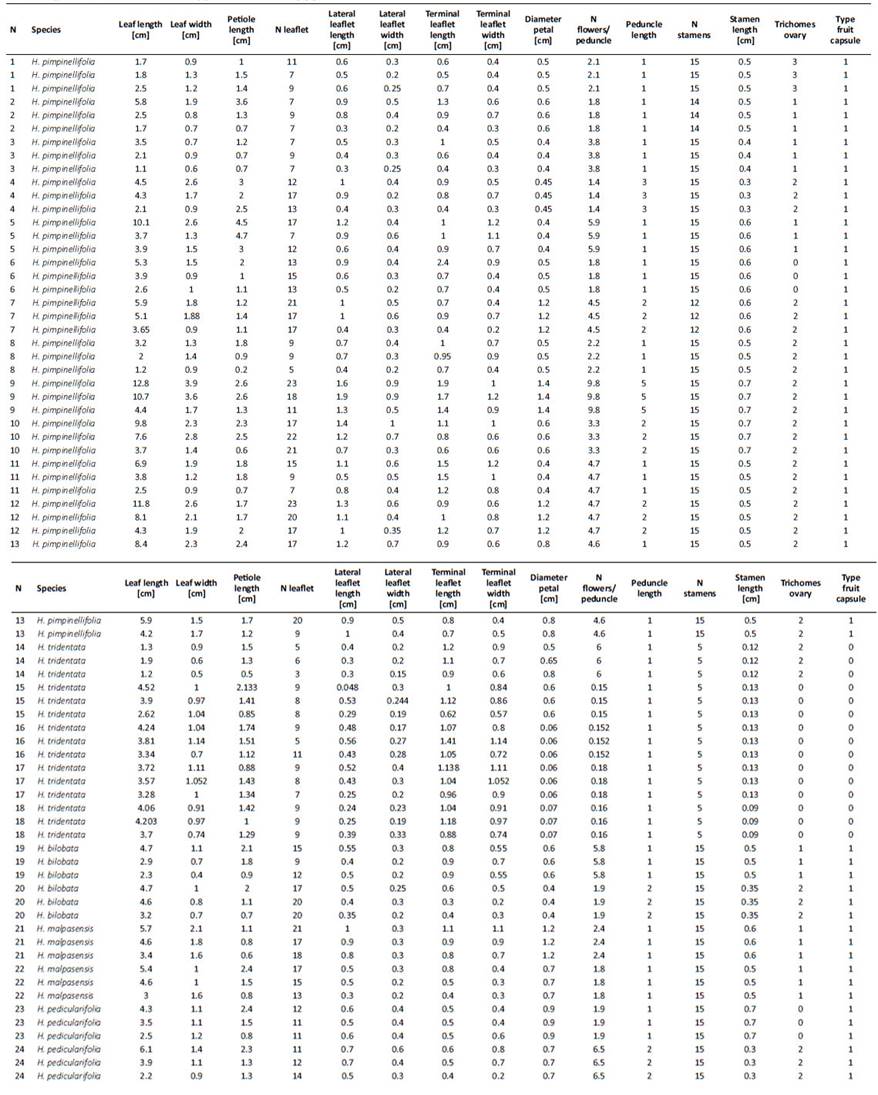
Qualitative characters were described according to The Cambridge Illustrated Glossary of Botanical Terms (Hickey, King, & King, 2000). Three leaves per specimen were selected according to size (big, medium and small) and one of them (generally the largest one) was used to characterize the leaflets (width and length measurements). From flowers, petal diameter, number of flowers per peduncle, number and length of stamens were measured, and we included types of trichomes present inthe ovary as well. Finally, type of fruit was characterizedaccording to Slanis and Grau (2001) into capsule or schizocarp (Supp. Table A1). The morphological data matrixwas analyzed with Principal Component Analysis (PCA) in24 individuals in total, using the open source software R(R Core Team, 2019) with a previous logarithm transformation to account for differences in unit measurements. Taxonomic descriptions were complemented by the inclusion of specimens from Peru, Bolivia and Argentina.
Results and discussion
A total of 24 Bolivian specimens were quantitatively analyzed corresponding to 5 "species" as described by Knuth (1908): Hypseocharis pimpinellifolia (n=13), H. bilobata (n=2), H. malpasensis (n=2), H. pedicularifolia (n=2) and H. tridentata (n=5) (Supp. Table A1).
Three types of leaves: pinnate, bipinnate and bipinna te with ternate leaflets (Supp. Figure A1) and two typesof trichomes located on the ovary were identified: simple and glandular ones (Figure 6G). A summary of the main reproductive and vegetative characters is presented in Table 1. Hypseocharis pimpinellifolia presented a high variabilityin leaf type and flower color; while H. tridentata differed from the remaining specimens in the number of stamensand type of fruit (Table 1). Principal Component Analysisshowed similar results based on a combination of quantitative vegetative and reproductive characters, depicting aclear separation of H. tridentata (Figure 2). The first fivecomponents accounted for 90% of the total variation. The first component was mainly explained by peduncle length; while the second component was mainly explained by leaf length and number of leaflets (Table 2, Figure 2).
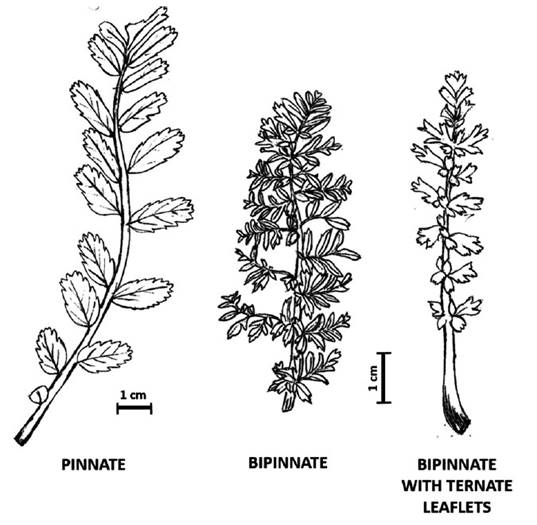
Supplement Figure A1. Types of leaves of the genus Hypseocharis in Bolivia and Peru. A: Terán J 535 (HNB); B: R Ehrich 63 (LPB); C: Salomón Pérez 26 (HNB). Drawings by S. Lozada-Gobilard.
Table 1 Summary of reproductive and vegetative variables measured in Hypseocharis specimens from Bolivia. Hyphenated species names assigned to technical characters used in the literature (Sch: Schizocarp, VC: Ventricidal Capsule; P: Pinnate, BP: Bipinnate, BPtl: Bipinnate with ternate leaflets; W: White, Y: Yellow, O: Orange, R: Red; G: Glandular, S: Simple).
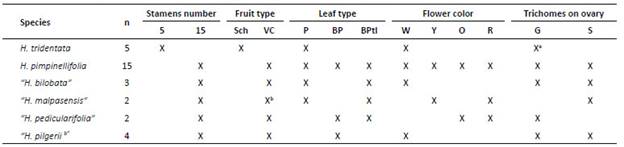
a Data taken from Slanis and Grau 2001
b Weigend (pers. obs.)
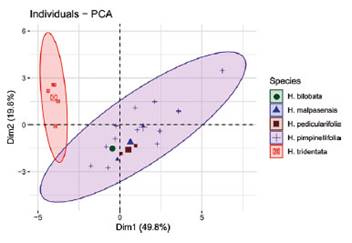
Figure 2 Principal Component Analysis (PCA) based on vegetative and reproductive traits from the five "species" of Hypseocharis in Bolivia. Species names assigned based on technical characters as used in the literature. There is no clear separation of any of the species apart from H. tridentata. First component explain 49.8% of the total variation and separate individuals mainly based on peduncle length while second component explains 19.8% and is based on vegetative characters of leaf length and number of leaflets. See details in Table 2.
Table 2 Proportional contribution of variables for the first five PCA components. Vegetative characters mainly explain by the second and fifth components, while reproductive ones are explained by components one, three and four. Numbers in bold show the highest contribution to the correspondent component.
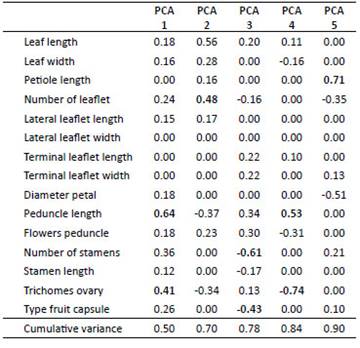
Hypseocharis tridentata is not as common and widespread as H. pimpinellifolia (Figure 1), but it is morphologically highly differentiated with terminal leaflets distinctively larger than the lateral ones, with a tridentate apex or emarginate with a medial mucro or tooth (Figure 4, Figure 5, Figure 6; Macbride 1949; Slanis & Grau 2001). Hypseocharis tridentata also differs in both flower and fruit morphology (Table 1): It differs from H. pimpinellifolia by 5 stamens per flower (versus 15) and a schizocarpic fruit with five dehiscent mericarps and 1- few seeds per locule versus a ventricidal capsule (Slanis & Grau 2001, Figure 3). The only specimen of H. tridentata present in the LPB Herbarium plus four specimens from BOLV Herbarium corresponded to the published descriptions of the fruit (Figure 4).
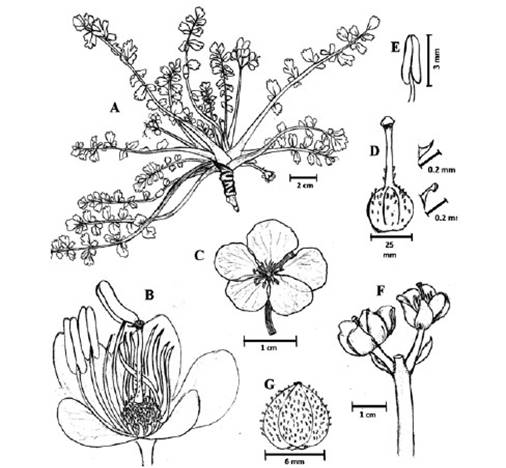
Figure 3 Hypseocharis pimpinellifolia A. Plant B. Flower C. Frontal view of the flower with imbricate arrangement of the petals D. Ovary with simple trichomes on the style and glandular ones on the ovary E. Stamen F. Scapiform peduncle G. Fruit. A: Tejeda WT18, LPB; B, G: Caro-Rojas 1564, LPB; C: Huaylla, H & R. Lozano 1175, HSB; D, E: M. Huanca et al. 2916, MO; F: Yanapa 274, LPB. Drawings by S. Lozada-Gobilard.
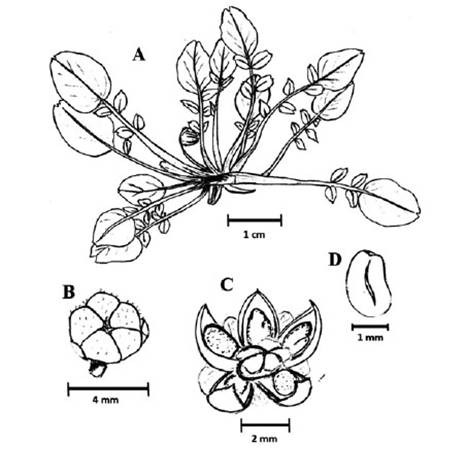
Figure 4 Hypseocharis tridentata A. Plant B. Schizocarp closed fruit. C. Opened fruit showing 5 mericarps 1-seeded D. Seed A-D: R. Baar 256, LPB. Drawings by S. Lozada-Gobilard.

Figure 5 Proposed division of species of the genus Hypseocharis (species names assigned based on technical characters as used in the literature). A-F Hypseocharis pimpinellifolia. Synonyms of H. pimpinellifolia: H. pilgeri (A), H. pedicularifolia (D), H. malpasensis (E), H. bilobata (F). G H.tridentata. A: H. "pilgeri" (Graciela Vilcapoma Segovia & al. 3257, USM); B: H. pimpinellifolia (Caro-Rojas 1564, LPB); C: H. pimpinellifolia (Evert Thomas ET131, LPB); D: H. "pedicularifolia" (R. Ehrich 63, LPB); E: H. "malpasensis" (Beck St. G. 26091, LPB); F: H. "bilobata" (Beck St. G. 16849, LPB); G: H. tridentata (R. Baar 256, LPB). Photos M. Weigend, USM (A), S. Lozada-Gobilard, LPB (B-G).

Figure 6 Comparison of the proposed species of the genus Hypseocharis based on vegetative and reproductive traits. A-G Hypseocharis pimpinellifolia. Synonyms of H. pimpinellifolia: H. pilgeri ( C-E), H. bilobata (F). H Hypseocharis tridentata. H. pimpinellifolia presents a high diversity, leaves can be pinnate (A), bipinnate (B, D) or bipinnate with ternate leaflets (F) (See Supplementary Figure A1 for detailed drawings). Flowers possess 15 stamens with separate (E) or close petals (A, B, F), orange (A) or white (B, E F), but also can be yellow or red. Ovary possess simple (s), glandular (g) or both types of trichomes (G). On the other hand, leaves of Hypseocharis tridentata are pinnate with the terminal leaflet distinguished larger than the rest and flowers are white with 5 stamens only (G). Photos: Teresa Ortuño (A), Alfredo Grau (B) M. Weigend (C-F), S. Lozada-Gobilard (G). Used with permission from AA. M. Anton and F. O. Zuloaga (directors), Flora Argentina. 28th November 2019 http://www. floraargentina.edu.ar (H).
Additionally, a comparison of cultivated material of "H. pimpinellifolia" from La Paz (Ortuño 1361) with an observation of living plants of "H. pilgeri" at Canta (Depto. Lima, Weigend & Peña Ramos 9715) showed no differences whatsoever in leaf dissection, flower morphology or indument (Figure 6C-E). A comparison of additional material from Lima clearly confirmed that H. pilgeri cannot be differentiated from H. pimpinellifolia (Figure 5A-F).
Conclusion
Overall, we confirm and expand the observations by Slanis and Grau (2001) from Argentina: H. pimpinellifolia is a highly variable species with regards to details of leaf dissection and flower colour, but the recognition of additional taxa is not warranted. All specimens seen from Argentina to northern Peru belong to a morphological continuum and are best recognized as belonging to a single species. Bolivian H. malpasensis, H. pedicularifolia, and H. bilobata and Peruvian H. pilgerii are therefore synonymized with H. pimpinellifolia. Hypseocharis pimpinellifolia and H. tridentata are recognized as the only two species in the genus. There are some differences in leaf dissection and flower color, which - at face value - might warrant the recognition of intraspecific entities. However, there is no clear geographical pattern in the distribution of different "morphs" and the differentiating characters do not appear to be at all combined in any meaningful way, as the statistical analysis shows.
Key to the species of Hypseocharis Leaves pinnate, up to 2 cm long, terminal leaflet much larger than lateral leaflet; flowers with 5 stamens, petals white; fruit a schizocarp with five 1-seeded mericarps H. tridentata
Leaves pinnate, bipinnate or bipinnate with ternate leaflets, (1-)5-13 cm long, terminal leaflet marginally larger than lateral leaflets; flowers with (12-)15 stamens, petals white, yellow, orange or red; fruit a ventricidal capsule H. pimpinellifolia
Formal taxonomy
Hypseocharis pimpinellifolia J. Rémy -- Ann. Sci. Nat., Bot. sér. 3, 8: 239. 1847 [Oct 1847].
Type:-- BOLIVIA, Santa Cruz, Prov. Valle Grande, in dry grassland of the high Andes by the Río Grande, not far from Puccara, at 3000 m, D´Orbigny 1137 (P00835307!).
= Hypseocharis bilobata Killip -- J. Wash. Acad. Sci. 16: 570. 1926. Holotype:-- PERU, Cuzco, 3000-3600 m, F. L. Herrera s.n. (US00101084!).
= Hypseocharis corydalifolia R. Knuth -- Bot. Jahrb. Syst. 41(3): 173. 1908. Holotype:-- ARGENTINA, Prov. Catamarca, near Capillitas, anno 1878, Schickendantz 306 (B†; isotype: CORD00002963!).
= Hypseocharis fiebrigii R.Knuth -- Bot. Jahrb. Syst. 41(3): 173. 1908. Holotype:-- BOLIVIA, Cuesta Vieja, at 3100 m, Fiebrig 3296 (B†).
= Hypseocharis malpasensis R. Knuth -- Repert. spec. nov. Regni Veg. 29: 219. 1931. Lectotype (here designated):-- BOLIVIA, Chuquisaca, Prov. Zudañes Cuesta de Malpaso, Cordillera de Sombrero, 3100 m, anno 1927, C. Troll 797 (B 10 0002189!, isotypes: 2 specimens; holotype: M 0172361!).
= Hypseocharis moschata R. Knuth --Meded. Rijks-Herb. 27: 67. 1915. Type:-- BOLIVIA, on short-grassed alpine meadows of the Sunchalkammes, ca. 2900 m, 04.1911, T. Herzog 2013 (Z-000017708!; L.2008782!; S10-22464).- Knuth described part of that collection as moschata, but assigned another part of Herzog 2013 (L.2008781!) to fiebrigii.
= Hypseocharis pedicularifolia R. Knuth -- Bot. Jahrb. Syst. 41(3) 173. 1908. Lectotype (here designated):-- BOLIVIA [Depto. Tarija], Prov. Cercado, Puna Patanca, at 3700 m, on the gently sloping edge of the plateau, Calderillo, around 3000 m, on the valley plain between grasses, Fiebrig 2626 (F 218086!; B 10 0279796!, isotypes: K000073728!, MO 2125259!, GH 00043695!).
Type Specimens
= Hypseocharis pilgeri R. Knuth -- Bot. Jahrb. Syst. xli. 174 (1908). Holotype:-- PERU [Depto. Lima, Prov. Huarochiri], between Norquinia and Matucana, on stony soil near the river between 2200 and 2700 m, 12.1901, A. Weberbauer 95 (B†).
- Hypseocharis pimpinellifolia J. Rémy var. punicea Kuntze --Revis. Gen. Pl. 3[3]: 33. 1898 [28 September 1898]. Petala punicea, Bolivia, 3600 m, Tiraque, O. Kuntze? (B†). - Kuntze gives only the locality, not the collector. Since he describes both his own collections as various collections from colleagues in this treatise, it is uncertain whose collection the name was based on.
Diagnosis:
Acaulescent rosette herb. Root tuber 5-40 cm long and up to 7 cm thick, leaves narrowly oblong in outline, (5-)10-20 cm long, lamina 1-13 × 0.5-4.0 cm pinnate, bipinnate or bipinnate with ternate leaflets. Petiole 0.2-4.7 cm long, pubescent (rare glabrous). Leaflets up to 26, ovate or elliptical, leaflets subsessile, alterna-te or subopposite, margins dentocrenate (rarely entire), deeply pinnatisect to pinnate with 3-4 incisions on each side, usually deeply pinnatifid or pinnate into narrowly oblong-acuminate lobules or ovate-acuminate leaflets, terminal leaflet deeply dissected, not clearly set off from rachis. Lateral leaflets 0.3 × 0.3 to 2 × 1 cm, obovate or more commonly elliptic in outline, apex dentate, 3-dentate (rarely 2 or 5-dentate) or entire; terminal leaflet 0.4 × 0.2-2.4 × 1.2 cm, obovate, apex dentate or 3-dentate (rare 2-dentate), slightly larger than lateral leaflets, usually lobed. Pedicels (5-) 10-20 cm, pubescent, with 1, more rarely 2-5 (-8) flowers. Petals subcircular, obovate or obtriangular, 1-2 × 1-2 cm, red, orange, white or yellow. Stamens 15, dorsifixed, with filaments 0.3-1.0 cm long. Ovary pubescent with two type of hairs, simple hairs on the style, glandular and/or simple hairs on the ovary. Fruit a ventricidal capsule, locules with (1-) 2-3 seeds per carpel (Figure 3).
Specimens seen:
PERU:
Depto. Ancash, Prov. Carhuaz, Huascarán National Park, Quebrada Ishinca, 9°23´S, 77°29´W, 3880 m, 15.02.1985, D.N. Smith et al. 9607 (USM, HUT, MO).
Depto. Lima, Prov. Huarochiri, Matucana, 2650 m, 12. 04-03.05.1922, J. F. Macbride & Featherstone 206 (F, USM). Below Matucana, 2200-2300 m, 02.1909, A. Weberbauer 5200 (F!). Side valley south of Matucana, 2400 m, 22.03.1942, T.H. Goodspeed 33072 (MO). Km 56 route Lima-La Oroya, 1400-1500 m, 20.02.1954, R. Ferreyra 9650 (USM). Chacahuaro near Matucana, 2500-2650 m, 09.03.1953, R. Ferreyra 8969 (P, USM). Valley south of Matucana, 2400 m, 22.03.1942, T.H. Goodspeed 33072 (US).Prov. Canta, San Buenaventura, 2650 m, 30.04.1994, G. Vilcapoma S. et al. 3257 (USM). Near Canta, 27.01.1963, I. Meza 22 (USM). Road from Canta to Lima, on the first road bend from Canta, 11° 47’ 798’’ S, 76° 62’ 869’’ W, 2738 m, 01.03.2018, M. Weigend & K. A. Peña Ramos 9715 (BONN, USM).
Depto. Junín: Prov. Jauja, Ricrán, 3750 m, 09.04. 1967, B. Hilario 1 (P, USM). Tajana, 3900 m, 26.01.1980, C. Hastorf 319 (USM). Incahuasi, 03.1947, J. Soukup 3228 (US). Prov. Tarma, Río Mantaro between Ororya and Huancayo, 3570 m, 19.02.1964, P.C. Hutchison & O. Tovar 4207 (P). Laive, 3820 m, 19.04.1983, B. Maas 1008 (USM). Prov. La Oroya, Pachacayo, 3700 m, 22.05.1974, P. Gutte 2137 (USM). Pachacayo, 3800 m, 11.1981, K. Tiller 107 (USM).
Depto. Ayacucho, Prov. Lucanas, between Puquio and Nazca, 3400-3500 m, 20.03. 1949, R. Ferreyra 5501 (P, USM). Prov. Huanca Sancos, road Putajasa to Huanca Sancos, 3700-3730 m, 13° 58´ 10,6´´ S, 074° 18´08,7´´ W, 25.02.2002, A. Cano et al. 12029 (USM). Lucanas, no precise locality, 3800 m, 13.03.1954, Rauh-Hirsch P453 (F).Road Putajasa to Huanca Sancos, 3700-3900 m, 14° 00´ 02,6´´S, 074° 17´14,1´´W, 25.02. 2002, A. Cano et al. 12019 (USM). Sacsamarca, 3500-3600 m, 13° 57´ 51,2´´ S, 074° 18´ 45,1´´ W, 25.02. 2002, A. Cano et al. 12042 (USM). Aya-cucho, no precise locality, 01.1995, M. Romero s.n. (USM).Prov. Huamanga, Ullpachaka, 3580 m, 29.01.1974, L. Arraguirre et al. 3015 (USM). Depto. Huancavelica, Prov. Tayacaja, Hacienda Huari, 4300-4600 m, 18.07.1955, E. Salaverry C. 135 (USM).
Depto. Cuzco, Prov. Cuzco, Choko, 3400 m, 02.1937, C. Várgas C. 233 (F). Prov. Quispicanchis, Vilcanota Valley near Urcos, 3800 m, 27.02. 1929, A. Weberbauer 7883 (F).Distr. Marcapata. near Ocongate, 3350 m, 11.12.1938, C. Vargas 9722 (F). Canchis, E of Sicuani, 14° 16’ S, 71° 13’ W, 3450 m, 28-29.03.1988, P. Núñez 8912 (P, MO). Prov. Urubamba, Chincheros, trail Antasaka rock to Parqo spring, 13° 24’ S, 72° 03’ W, 3810 m, 12.01.1982, W. Davis et al. 1361 (USM). Prov. Kaira, 3300 m, 18.03.1948, A. López 676 (HUT).
Depto. Arequipa, Prov. Cailloma, above Chivay, 3900 m, 04.1914, A. Weberbauer 6899 (F). Prov. Cabanaconde, 15° 36’ 32’’ S, 71° 52’ 29’’ W, 3725 m, 02.04. 2005, C. Aedo & A. Galán de Mera 11081 (USM).
Depto. Moquegua, Prov. General Sánchez, Cerro, Distr. Lloque, Ccellan to Lucco, 16° 16´ 20´´ S, 070° 44´38´´W, 3860 m, 12.04.2012, D. B. Montesinos 3768 (USM).
Depto. Tacna, Candarave, 2900-3000 m, A. Weberbauer 7381 (F). Prov. Tarata, Talabaya, 4500-4700 m, 30.03.1998, A. Cano 8287 (USM). Without precise locality: Peru, Cl. Gay 1099 (P).
BOLIVIA:
Depto. Potosí, Prov. Quijarro, 3700 m, 16.12. 1987, M. Schulte, 70 (LPB). Prov. Tomás Frías, 3800 m, 06.02. 1988, St. G. Beck, 137 (LPB). Prov. Sud Chichas, 21°29’ 02’’ S, 65°41’ 35’’ W, 3439 m, 11.03. 2012, R. F. Zenteno et al. 12040 (LPB). Potosí, open low grassland, 3800 m, 05.03.1981, St. G. Beck 6144 (LPB).
Depto. La Paz, 3750 m, 02.1913, O. Buchtien s.n. (F). LaPaz, 3750 m, 02.1913, O. Buchtien 643 (US). La Paz, 3700 m, 18.01.1909, O. Buchtien s.n. (B). Near La Paz, 3800 m, 02.1857, G. Mandon 787 (P). Prov. Murillo, near to "Muela del Diablo" mountain, 3811 m, 22.10.2002, ES 18,(LPB). Near to "Muela del Diablo" mountain, 18.02. 1979, M. Liberman 11 (LPB). Cerro Muela del Diablo, 16°35’ S, 68° 08’ W, 3700 m, subpuna, 01.02. 2011, Teresa Ortuño 1361 (LPB). Rocky soil, 3800 m, 18.03.1989, St. G. Beck 19849 (LPB). Near Muela del Diablo 16° 32’S 68° 06’ W, 3671 m, 07.12. 2012, W. Tejeda, 18 (LPB). Below Muela del Diablo, 3800 m, 18.03.1989, St. G. Beck 16849 (LPB). Rio Zongo Valley, cerca la escuela de Cambaya, 2400 m, 16° 08’ S, 68° 06’ W, 30.01.1988, J. C. Solomon 17774 (F, P, MO). Prov. Aroma, 17° 22’ S, 68° 15’ W, 17.02.2005, R. Yanapa, 274 (LPB). Cantón Belén, Comunidad Huaraco, 3800 m, 11.02.1979, St. G. Beck 338 (LPB). Huaraco en la loma, 150 km sud de La Paz, 3750 m, 01.10.1981. Liberman 304 (P).
Depto. Chuquisaca, Prov. Zudañez, Cantón Presto, Tomoroco, 19°55’ S, 64°56’ W, 2500 m, 01.1978, W. E. Carter s.n. (F). Cantón Presto, Tomoroco, 19° 55’ S, 64° 56’, 2500 m, 01.1978, W. E. Carter s.n. (F). Open stony slopes with rough grassland, scattered low shrubs and tough perennials with occasional Polylepis graves; apparently frequent on flat open areas between rocks, 2900 m, 2.04.2000, J. Wood & A. Carretero 16183 (LPB). Prov. Nor Cinti, San Lucas, Pucaloma, 20° 15’ 51.8’’ S, 65° 06’ 4.4’’ W, 3350 m, 05.02.2003, L. Rico et al. 1250 (K, F). 01.1846, Weddell 3986 (P). Prov. Tomina, anno 1846, A. Weddell 3933 (P). Anno 1846, A. Weddell 3986 (P). Prov. Belisario Boeto, steep surface with grass plants, 2650 m, 03.07.1993, J. Terán 535 (LPB).
Depto. Cochabamba, Prov. Tarata, Anzaldo, 3600 m, 09.01.1943, M. Cardenas et al. 7615 (F). Prov. Mizque, Lagunilla on Mule trail between Chuguarani and Quioma, 3200 m, 01.04.1939, W. J. Eyerdam 25086 (F).2800 m, 2.12.1986, M. Sigle, 189 (LPB). Prov. Capinota,17° 53’ 7.63’’ S, 66° 14’ 9.14’’ W, 3530 m, 25.01.2003, T. Evert, 131 (LPB). Prov. Ayopaya, in a crop border, 3400 m, 01.2001, R. López, 26 (LPB). Cona cona, 18° S, 66° 45’ W, 4300 m, 03.02.1949, W. M. A. Brooke 5192 (P), Prov. Quillacollo, Monte Tunari, 3650 m, 17.01.1971, A. Lourteig 2593 (P). Near snowline, Tunari, anno 1891, M. Bang 1043 (GH, MO). Quillacollo steep slope with remnants of Polylepis and young planted Eucalyptus, area heavily grazed with rocky soil, 17° 16’ 9’’ S, 66° 19’ 40’’ W, 3650 m, 27.01.1996, N. Ritter and J. Wood 2820 (LPB, MO, NHA). Prov. Tiraque, Toralapa, 3430 m, 12.01.1995, E. Emshwiller et al. 233 (LPB, F).
Depto. Tarija, Prov. Cercado, 2500 m, 11.11.1980, C. Rojas, 1564 (LPB). Aliso forest, 2950 m, 23.01.1986, E. Bastian, 524, Cuesta de Sama, 3250 m, 10.02.1982, G. Gerold 199 (F, P). Prov. B O’Connor, open Puna and high Andean vegetation, 2700 m, 10.03.2006, F. Zenteno, 4551 (LPB). Prov. Tapacari, Pongo, 3950 m, 17° 37’ S, 066° 39’ W, J.C. Solomon & M. Nee 18091 (F, P, MO). Prov. Aniceto Arce Ruiz, open grassland with Northeast exposition, rocky soil with a few bushes, 21° 52’ S, 64° 52’ W, 2700 m, 15.03.1998, St. G. Beck et al. 26091 (LPB). 3400 m, 19.01.1986, R. Ehrich 63 (LPB).
Depto. Santa Cruz, Prov. Vallegrande, humid grassland with nebline in the morning and strong winds in the afternoon, 18° 42’ 38’’ S, 64° 08’ 02’’ W, 2715 m, 22.01.2011, M. Huanca et al. 2916 (MO).
ARGENTINA:
Prov. Jujuy: Sierra de Zenta, 4000 m, 02.1931, E. Budin 7452 (2 × F). Depto. Yavi, La Quiaca, 3450 m, 16.02.1940, T. Meyer 33914 (F). 13 km of Abra Pampa on road to La Quiaca, 3500 m, 24.01.1982, W. R. Anderson 12352 (P). La Quiaca, 3450 m, 11.02.1940, T. Meyer 33918 (F). Abra Pampa, 3700 m, 30.01.1971, Ellenberg 392 (P). La Quiaca, 3442 m, 24.01.1940, Schreiter 11043 (F). Depto. Tumbaya, ascent to Abra Morada, 2000-3200 m, 05.03.1965, A. L. Cabrera et al. 16964 (HUT, P). Depto. Tilcara, Tilcara, 12.02.1926, S. Venturi 7627 (US). Depto. Humahuaca, Tres Cruces, 3700 m, 02.02.1968, A. L. Cabrera et al. 18929 (P).
Prov. Tucumán: Depto. Tafi, El Molle, Quebrada Amaicha, 2600-2700 m, 12.1931, Schreiter 6776 (F). Tafi del Valle to Ameicha, 2080 m, 20.12.1965, H. & E. Walter 622 (B). Sierra Famatina, between La Encrucijada and Las Cuevas, 03.02.1879, Hieronymus & Niederlein 345 (B,CORD).
Prov. La Rioja: Depto. Gral Roca, Mina San Juan, Sierra Famatina, 3050-3200 m, 02.03.1906, F. Kurtz 13673 (P). Barrancas Amarillas, between La Mariposa and Rodeito, 2100 m, 27.02.1906, R. Gaitan 13450 (P).
Hypseocharis tridentata Griseb., Nachr. Königl. Ges. Wiss. Georg-August-Univ. 20: 495. 1877. Holotype:-- ARGENTINA [Salta], Los Potreros at the foothills of Nevado del Castillo, Hieronymus & Lorentz 131 (GOET009182!,isotype: CORD00002965!).
Diagnosis:
Acaulescent rosette herb. Root tuber 4-6 cm long and 2-3.5 cm thick, leaves 1.2-1.9 cm long, 0.5-0.9 cm width, pinnate. Petiole 0.5-1.5 cm long, glabrate or pubescent with dilated base, margin almost membranous. Leaflets 11-17, entire, alternate or subopposite, sessile, lateral one 0.5-1.4 cm long, oblong or elliptic, terminal one larger than lateral ones, terminal leaflets 0.7-1.8 cm long, ovate, entire or with obtuse three dentate apex or more common with a middle teeth or mucron. Pedicels 6 cm long, 1-flowered. Petals 0.5-0.8 cm in diameter, caducifolious, white. Stamens 5 in a cycle, opposite to the petals. Ovary densely glandular. Fruit a schizocarp, with 5 dehiscent mericarps, 1-pauciseminades (Figure 4).
Specimens seen:
BOLIVIA. Depto. Cochabamba, Prov. Ayopaya, 3270 m, 11.04.1989, R. Baar 256 (LPB). Morochata, rocky slope with low dense grassland with signs of fire, 3809 m, 17° 13’ 45.9’’ S, 66° 26’ 33’’ W, 15.03.2020, Miranda G. 61. (4 × BOLV). ARGENTINA. Prov. Jujuy. 13 km N of Abra Pampa on road to la Quiaca, 3500 m, 24.01.1982. Willian R. Anderson 12352 (P). Depto. Valle Grande: Santa Ana, grasslands, 29.02.1940, Burkart & Troncoso 11690 p.p. (SI). Salta. Depto. Caldera: Potrero del Castillo, 2500 m, 14.03.1952, Sleumer & Vervoorst 2818 (US). Depto. Orán: Volcán, next to Gasoduct Nor Andino, 2800 m, 27.12. 1999, Grau et al. s.n. (LIL). Depto. Santa Victoria: on the way from Santa Victoria to Los Toldos, El Chorro, 3100 m, 01.1997, Grau & Seaman s.n. (LIL); Depto. Iruya: on the way up to Volcán a Zenta, 2600 m, 12.2000, Grau s.n. (LIL). Tucumán. Depto. Tafí del Valle: El Rincón, 2600 m, 20.02.2001, Slanis & Grau 1755 (LIL). No observations were registered in Peru.

