











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La pérdida de bosques en la selva tropical ocasionada por deforestación es un problema global que ocasiona pérdida de biodiversidad (MEA 2003, Foley et al. 2007), en particular, la cuenca amazónica como un gran foco de pérdida de selva tropical (Potapov et al. 2014).
La teledetección y los Sistemas de Información Geográfica (SIG) permiten comparar estudios de cambios en el uso de la tierra a través del tiempo (Cigliano & Torrusio 2003); mientras que imágenes de la superficie terrestre permiten detallar la cobertura de suelo y evaluar los cambios en la vegetación (Cabello & Paruelo 2008). En el marco de la conservación de los bosques del Perú, el Ministerio del Ambiente (MINAM) desarrolló la plataforma GEOBOSQUES para el "monitoreo de la cobertura de bosque, cambio de bosque a no bosque y la degradación forestal". Esta plataforma distribuye información a los distintos usuarios (MINAM 2014).
La deforestación registrada en el Mapa Forestal del Perú (Malleux 1975) señaló 4’500000 ha de selva deforestadas, causada por agricultura y ganadería (MINAM & MINAGRI 2014). Registros de los años 2001 al 2014 muestran que la deforestación antrópica bruta causó la pérdida de 1’653121 ha de bosque (MINAM 2015). Por otro lado, en selva alta, la deforestación también es elevada y debida tanto a eventos naturales como a causas antrópicas (Dourojeanni et al. 2009, Franke et al. 2005). En particular, el departamento de Huánuco se ha observado la más alta pérdida de cobertura boscosa, tanto en el 2005 (25556 ha) como en el 2009 (24190 ha), además, de la más alta pérdida de bosque acumulada entre el 2000 y 2011 (MINAM 2014a, 2015). En este departamento se encuentran los bosques de Carpish, los cuales a pesar de contener un alto número de endemismo de flora y fauna (Parker & O’ Neill 1976, Young & León 1999, Beltrán & Salinas 2010, Jiménez & Pacheco 2016), y establecerse medidas para su conservación (El Peruano 2020), son amenazados por la extracción de leña y la apertura de campos de cultivo (Beltrán & Salinas 2010), siendo necesarias más investigaciones sobre su biodiversidad.
Los roedores cricétidos demuestran ser organismos de fácil muestreo debido a su comportamiento y roles en el ecosistema (Sunyer et al. 2013, Clark & Bunck 1991, Shomita et al. 2004, Manson et al. 2001), teniendo las características de un organismo que respondería rápidamente a la fragmentación (Santos & Tellería 2006). La diversidad de roedores en Perú se estima en 188 especies (Pacheco et al. 2018) siendo escasos los reportes para Huánuco (Pacheco & Noblecilla 2019). Así mismo, el conocimiento de cómo varían estas poblaciones ante el cambio del paisaje es poco conocido. Por lo que, este estudio tiene por objetivo analizar los resultados de diversidad y abundancia de los roedores cricétidos frente a la pérdida de bosque en Carpish en el año 2002 y 2017, tomando en consideración la estructura y composición de los mismos. Adicionalmente, se discute la respuesta de algunas especies de roedores considerados comunes y raras.
Material y métodos
El área de estudio se ubica en el departamento y provincia de Huánuco, distrito Chinchao, localidad San Pedro de Carpish (Fig. 1). El área de estudio pertenece a la ecorregión Yungas (sensu Dinerstein et al. 1995), o Bosque Húmedo Montano Oriental (Young & León 1999).
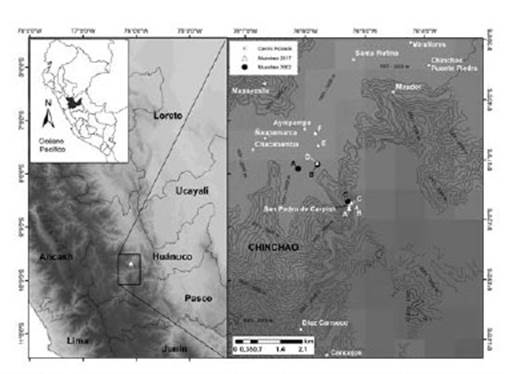
Figura 1 Ubicación del área de estudio en la localidad de San Pedro de Carpish. Evaluación del año 2002 (círculos negros): sitios de muestreo A, B y C. Evaluación del año 2017 (triángulos blancos): sitios de muestreo A, B, C, D, E y F.
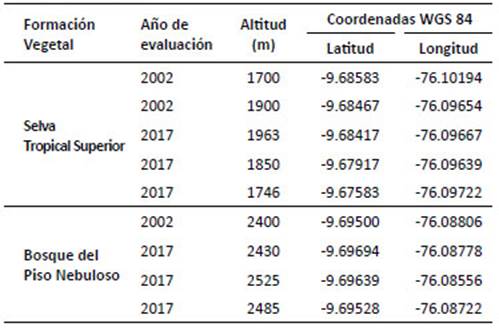
Localidad San Pedro de Carpish-SPC (1780 - 2400 m).- El paisaje se observa habitualmente cubierto de nubes. Las actividades humanas comunes son la ganadería, agricultura y extracción de madera. Cuenta con caminos que permiten acceder a la carretera Huánuco - Tingo María. La ganadería vacuna no está extendida por la topografía del lugar. En el área presenta gran variedad de vegetación arbórea, arbustiva y herbácea con predominancia de las rubiáceas y melastomatáceas, gran variedad de briofitas y abundantes epífitas. Beltrán y Salinas (2010) clasifican la localidad como formaciones vegetales de selva tropical superior y bosque del piso nebuloso:
Selva tropical superior-STS (1700 - 2300 m).- Esta formación evidencia el cambio de bosque montano alto a bajo, el dosel varía de 30 a 35 m de alto. Se observa lianas y epífitos, el sotobosque muestra arbustos de los géneros Piper, Palicourea, Miconia y Psychotria. El paisaje contiene amplias zonas deforestadas, donde se observa campos de cultivo de rocoto y papa, así como caminos hacia la carretera Huánuco-Tingo María.
Bosque del piso nebuloso (2300 - 2750 m) .- Esta formación contiene la mayor diversidad arbórea de todo el bosque, el dosel varía de 10 a 15 m. Se observa helechos arborescentes, epífitas (musgos, bromelias, orquídeas y aroideas). Se presenta un estrato conformado por la copa de árboles y otro compuesto por hierbas y arbustos del sotobosque. Es una zona medianamente afectada por acción del hombre debido a la extensión de campos de cultivo, ganadería y la construcción de carreteras, lo cual produce grandes claros de bosque.
Pérdida de bosque.- La información al nivel de distrito se obtuvo de la plataforma GEOBOSQUES (http://geobosques.minam.gob.pe/geobosque/view/perdida.php). Tablas y gráficas anuales fueron analizadas para observar los patrones en la pérdida de bosque. Además, se obtuvo la información de Concentración de pérdida de bosque entre los años 2001 al 2017 para el departamento de Huánuco, en coordinación con el encargado del Programa Nacional de Conservación de Bosques para la Mitigación del Cambio Climático - PNCBMCC.
Estimación de la diversidad y abundancia.- Se evaluaron tres sitios de muestreo en el 2002 y seis sitios de muestro en el 2017, todos ubicados en San Pedro de Carpish, considerando las partes más bajas y altas (Tabla 1, Fig. 1). Los ejemplares colectados en el año 2002 en San Pedro de Carpish, corresponden a la colección de Mastozoología del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (MUSM) los cuales ascienden a 41 ejemplares pertenecientes a cuatro especies. Los datos tomados incluyeron: número de especies, datos morfométricos (largo total, largo de cola, largo de oreja, largo de pata y peso), condición reproductiva, sexo, fecha de colecta, hora de captura de los ejemplares colectados, georreferenciación.
La evaluación del 2017 empleó la metodología dePacheco et al. (2007). Los sitios de muestreo incluyeron una trampa de golpe (Víctor o Museum Special) yuna trampa de caja (Sherman), por lo que se obtuvo180 trampas noche (NT) activadas por día. Se utilizóun cebo estándar preparado a base de mantequilla demaní, esencia de vainilla, avena y agua. Las trampasfueron preparadas a las 16:00 h y revisadas a las 08:00h del día siguiente. La evaluación se realizó por 12 noches. Mediante ayuda de la literatura especializada selogró la determinación de los especímenes hasta el nivel de especie (Patton et al. 2015, Jiménez et al. 2013,Jiménez & Pacheco 2016). Los especímenes se encuentran depositados en la colección del Departamento deMastozoología del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (MUSM). Lasestimaciones de la diversidad fueron comparadas conlos resultados del año 2002, de tal forma que se realizóun análisis temporal y espacial.
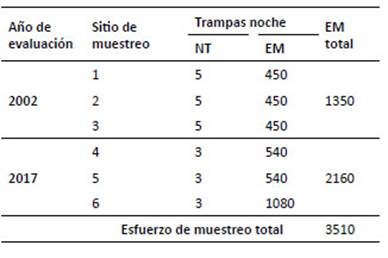
Las evaluaciones se desarrollaron durante 21 días en total y se emplearon 3510 TN, distribuidas en 1350 TNdurante el 2002 y 2160 TN durante el 2017 (Tabla 2). Enrelación a la representatividad del muestreo empleado, seelaboró la curva de acumulación de especies según la ecuación de Clench, para comparar los inventarios (Jiménez-Valverde & Hortal 2003) del 2002 y 2017. Mediante EstimateS (Colwell 2006) se calcularon los índices y se empleó Statistica (StatSoft Version 10) para graficar la curva.
Tabla 2 Esfuerzo de muestreo para roedores en las evaluaciones de los años 2002 y 2017 en la localidad San Pedro de Carpish. Donde: NT= número de transectos. EM= esfuerzo de muestreo.
La abundancia relativa (AR) se calculó por la razón entre el número de especímenes colectados entre el esfuerzo de muestreo en el sitio de evaluación, expresada en unidades de 100 trampas noche (Pacheco et al. 2007). Se empleó el coeficiente de correlación de Pearson para evaluar relación entre la AR con la altitud.
La diversidad alfa fue estimada con los índices de Shannon, Simpson, Equidad y el Índice de diversidad verdadera de orden 1 (1D), se empleó SPADE (Chao & Shen 2010). La diversidad beta se estimó mediante el índice de Whittaker y el índice de similaridad de Jaccard. Las estimaciones de la diversidad alfa y beta son útiles para comprender los cambios en la biodiversidad con relación en la estructura del paisaje (Whittaker 1972) y los efectos de las actividades humanas (Halffter 1998).
Análisis estadístico.- La existencia de una relación entre la perdida de bosque con la abundancia relativa yel número de especies en cada año de evaluación fue analizada mediante un ANOVA, que consideró: a) factor: lapérdida de bosque por año, b) variables: las especies y c)réplicas independientes del análisis: días de evaluación encada año. Además, para verificar la existencia de diferencias significativas entre las AR de las especies registradaspor año con la pérdida de bosque, se empleó el PERMANOVA (Anderson 2017), utilizando una matriz de disimilaridad (distancia euclidiana), para conocer la respuestade diversas variables (especies en este estudio) que sondependientes de diversos factores (pérdida de bosque),las validaciones se hacen a partir de permutaciones (Anderson 2017). El paquete Vegan versión 2.4-1 (Oksanen etal. 2016) se usó para el análisis en R (R DCT 2014).
Resultados
La pérdida de bosque en el distrito de Chinchao entre los años 2001 y 2017 ascendió a 2306 ha de bosque. Además, los años 2012, 2014 y 2017 registraron las mayores pérdidas de bosque (Fig. 2). Los datos cualitativos en cuanto a la concentración de la pérdida de bosque en Huánuco se observa en la Figura 3 y 4. La información cualitativa y cuantitativa en Carpish evidencia la pérdida de bosque en el área de estudio ocurrida en los años 2002 a 2017.
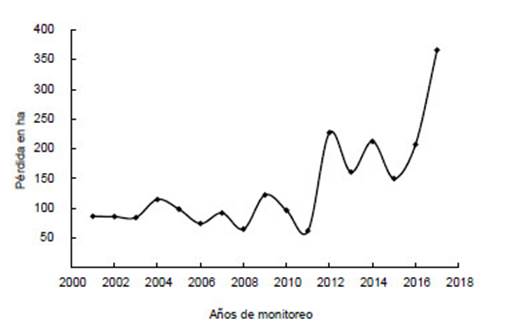
Figura 2 Pérdida de bosque (en hectáreas) en el distrito de Chinchao observada entre los años 2001 a 2017. Fuente: Plataforma Geobosques.
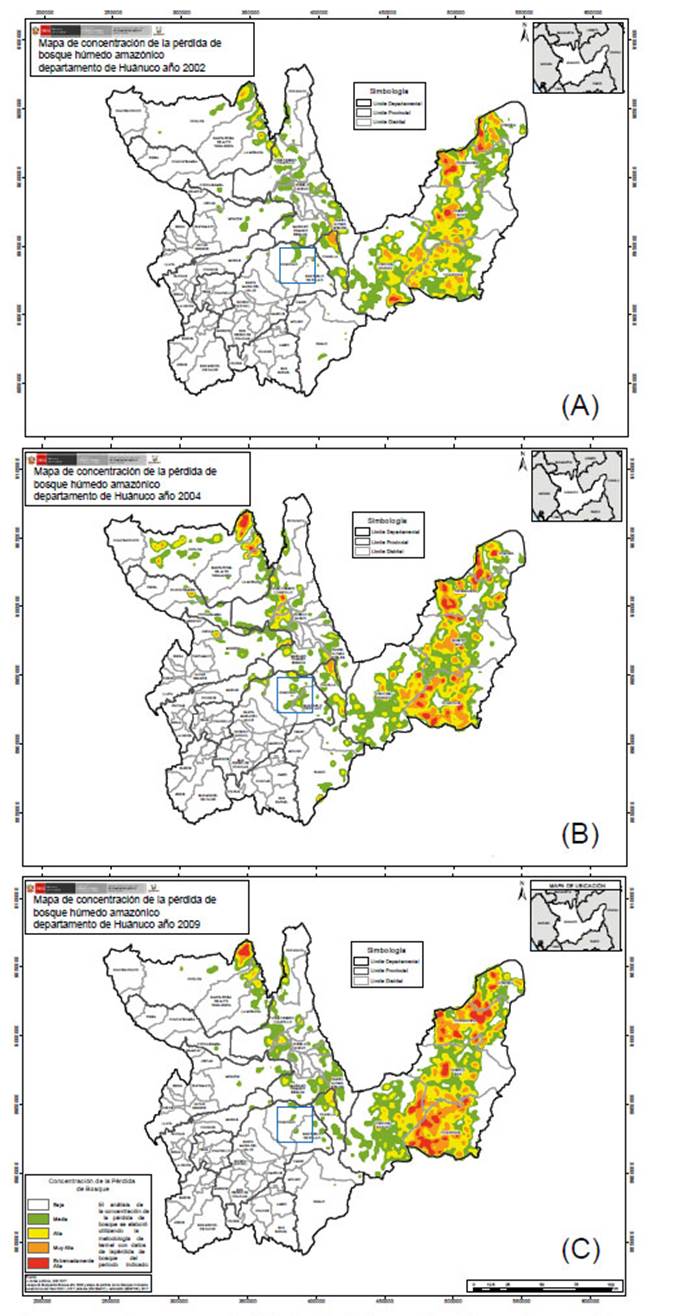
Figura 3 Mapa de concentración de la pérdida de bosque húmedo amazónico en el departamento de Huánuco. Recuadro en azul, distrito de Chinchao: A= año 2002. B= año 2004. C= año 2009. Pérdida media en verde, alta en amarillo, muy alta en anaranjado y extremadamente alta en rojo. Fuente: Programa Nacional de Conservación de Bosques para la Mitigación del Cambio Climático - PNCBMCC.
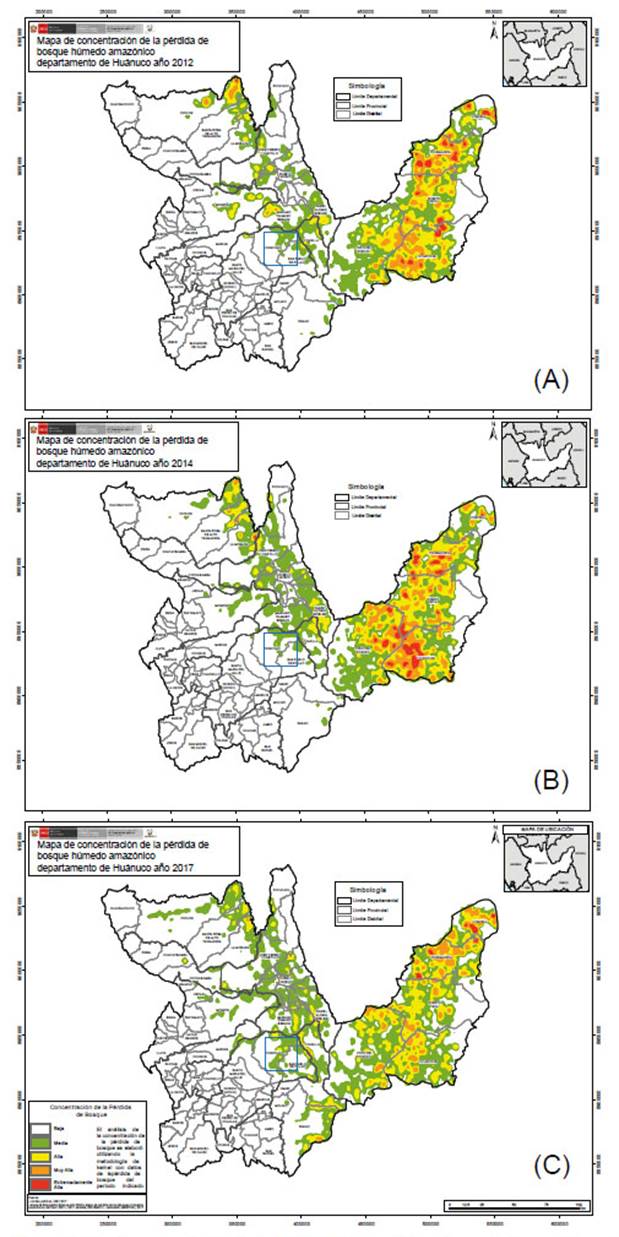
Figura 4 Mapa de concentración de la pérdida de bosque húmedo amazónico en el departamento de Huánuco. Recuadro azul, distrito de Chinchao: A= año 2012. B= año 2014. C= año 2017. Pérdida media en verde, alta en amarillo, muy alta en anaranjado y extremadamente alta en rojo. Fuente: Programa Nacional de Conservación de Bosques para la Mitigación del Cambio Climático - PNCBMCC.
En el 2002 se registraron 41 ejemplares de roedores cricétidos, representando cuatro especies; mientras que en el 2017 se registraron 82 ejemplares representando también cuatro especies. El número de especiesregistradas (Sobs=4) en el 2002 fue el 83.3 % del total esperado (Sesp= 4.8) y las especies registradas en el 2017 (Sobs=4) corresponde al 85.11% del total esperado (Sesp= 4.7); estos resultados muestran una evaluación bastante completa con los métodos de captura empleados. La curva de Clench muestra que a partir del día 9, la curva se estabiliza (Fig. 5 y Fig. 6).
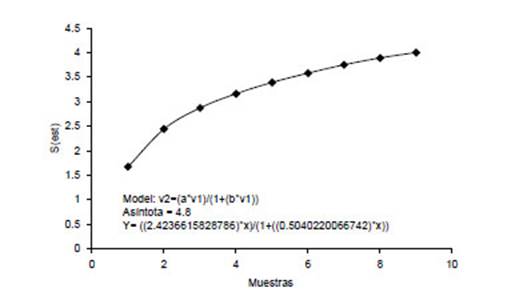
Figura 5 Curva de Clench para roedores cricétidos capturados en San Pedro de Carpish en el año 2002.
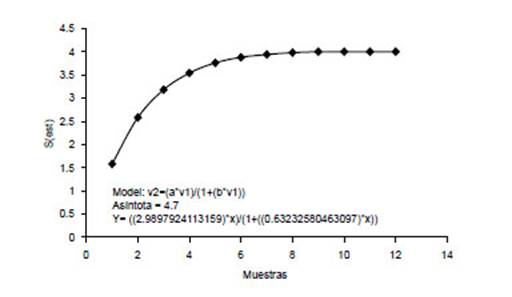
Figura 6 Curva de Clench para roedores cricétidos capturados en San Pedro de Carpish en el año 2017.
Akodon kotosh fue la especie con mayor registro (41 ejemplares, 2017), AR= 7.59 ind/100-TN (Tablas 3 y 4). Con relación a las AR por altitud (Tabla 4), fue alta en el año 2002 (6.44) a 1900 m y en el 2017 registró el más alto valor (10.93) a 2450 m. La AR por año y por especie (Tabla 5), mostró que O. destructor fue la especie más abundante en el 2002 (1.93 ind/100-TN); mientras que A. kotosh resultó más abundante en el 2017 (2.18 ind/100-TN). Es importante resaltar que Nephelomys keaysi fue registrada sólo en el 2002 y Thomasomys kalinowskii sólo en el 2017 (Tabla 3). La evaluación de la relación entre la AR y la altitud, resultó en una relación no significativa.
Tabla 3 Roedores cricétidos registrados por altitud en San Pedro de Carpish (SPC) durante los años 2002 y 2017.
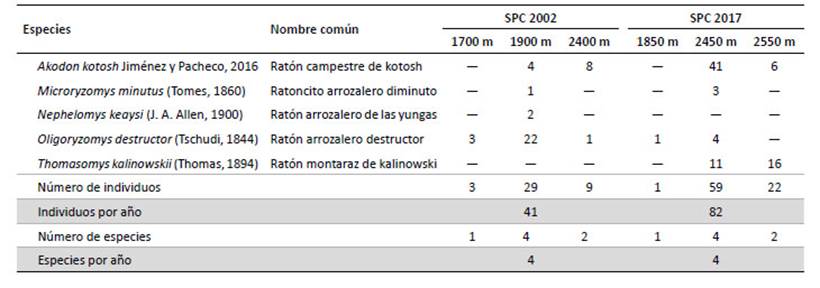
Tabla 4 Abundancia relativa (en base a 100 TN) por altitud para roedores cricétidos registrados en San Pedro de Carpish durante los años 2002 y 2017.
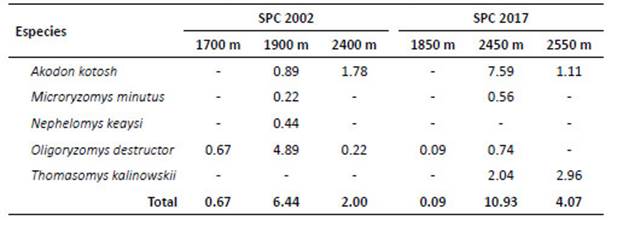
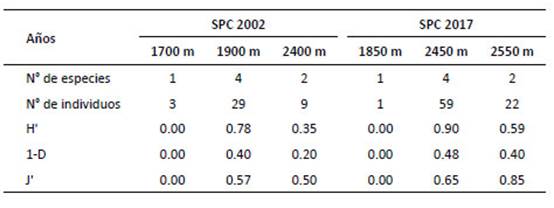
La evaluación reporta cinco especies en total; el análisis ANOVA evidenció la diferencia significativa del número de especies entre la pérdida de bosque o años de evaluación o (F Model= 5.87, Pr [>F]= 0.0255). Los resultados indican que O. destructor presentó la más alta densidad poblacional en el año 2002 con 1.93 ind/100-TN, mientras que A. kotosh lo presento en el año 2017 con 2.18 ind/100-TN. El PERMANOVA mostró que hay diferencia significativa en las AR de las especies en relación a la pérdida de bosque o años de evaluación (F Model= 835.59, Pr [>F]= 0.00099). La diversidad (Tabla 6) resultó alta en el 2017 (H’= 0.98; 1‒D= 0.56); sin embargo, la diversidad verdadera (1D) resultó baja para ambos años. Cuando se muestra la diversidad por altitud (Tabla 7), fue alta (H’= 0.90) a 2450 m; sin embargo, con el estimador Simpson, se muestra baja (1‒D= 0.48). La diversidad beta con el índice de Jaccard= 0.6, señala que existen especies compartidas entre los años 2002 y 2017; por lo que, "la composición de especies" es similar en los años de evaluación. El índice de Whittaker= 1.67, mostró el número de veces que cambia "la composición de especies" en los años de evaluación. Las especies compartidas son Akodon kotosh, Thomasomys kalinowskii y Microryzomys minutus. No se realizaron comparaciones con otros estudios ya que contienen diferencias en la amplitud del análisis, el tamaño de la unidad geográfica y región geopolítica, tales diferencias implican variaciones en las especies como en el ambiente, en esas condiciones la comparación directa no es posible (Koleff 2005).
Tabla 5 Abundancia relativa (en base a 100 TN) por año de los roedores cricétidos registrados en San Pedro de Carpish.
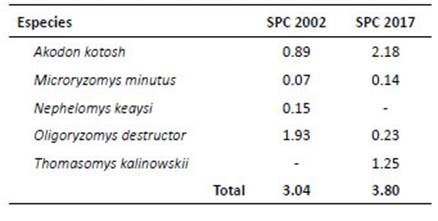
Tabla 6 Índices de diversidad de roedores cricétidos, número de especies e individuos capturados en la localidad de San Pedro de Carpish (SPC), años 2002 y 2017. Índices de diversidad: 1D= Índice de diversidad verdadera de orden 1 (q=1); H’= Shannon; 1-D= Simpson y J'= equidad.
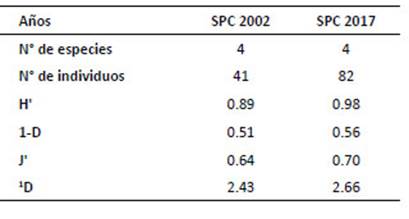
Tabla 7 Índices de diversidad de roedores cricétidos, número de especies e individuos capturados en la localidad de San Pedro de Carpish (SPC), años 2002 y 2017, detallado por altitud. Índices de diversidad: H’= Shannon, 1-D= Simpson y J'= equidad.
Los resultados mostraron disminución en la riqueza de especies puesto que en el año 2017 no se registró Nephelomys keaysi; mientras que, la abundancia descendió en dos de las cinco especies registradas en los años evaluados. Es importante señalar que aunque la pérdida de bosque creció al 2017 para el distrito de Chinchao, la abundancia relativa se mantuvo entre los valores más altos. Akodon kotosh fue la especie más abundante en San Pedro de Carpish; sin embargo, en el 2002 lo fue Oligoryzomys destructor. La pérdida de bosque no disminuyó la diversidad de roedores cricétidos, pero afectó su composición. Ante el aumento de perturbación humana y pérdida de bosque A. kotosh mantiene una alta abundancia.
Discusión
El número de especies de cricétidos en el presente estudio (San Pedro de Carpish 1700 ‒ 2550 m) es semejante al reportado para bosques de Huánuco entre los 2564 m y 3850 m (Noblecilla & Pacheco, 2012). Sin embargo, un estudio realizado entre los 2700 y 3000 m, en el bosque de Mayobamba en Carpish (Pacheco & Noblecilla 2019), se registraron ocho especies de roedores cricétidos. Lo cual podría indicar la importancia de las diferencias altitudinales. Por otro lado, las investigaciones de cricétidos en los bosques montanos centrales de Perú aún son escasos, en comparación con los realizados en el sur de Perú (Pacheco et al. 2007, 2011), donde se reportan 12 especies en la cuenca del río Apurímac, siete especies en Ayacucho, que incluso presentaba localidades impactadas (Pacheco et al. 2013).
La curva de Clench indicó que es poco probable que el número de especies en San Pedro de Carpish aumente con más muestreos, pero no se desestima el incremento a ocho especies según antecedentes en altitudes que varían de 1500 y 2500 m (Patton et al. 2015) y testigos (vouchers) depositados en Louisiana State University, Museum of Zoology (LSUMZ 12600, 14406, 14368). Así mismo, es probable que el empleo de diferentes métodos de captura (trampas Tomahawk, pitfall, etc.; Patterson et al. 2006) aumente el número del registro. Es preciso indicar que Thomasomys kalinowskii fue reportada, previo a este estudio, sobre los 2700 m (Pacheco & Noblecilla 2019) por lo que, es posible que otras especies sobre los 2500 m sean registradas en San Pedro de Carpish.
Oligoryzomys destructor fue la especie con mayor densidad poblacional en el 2002 pero mostró valores bajos en el 2017, año en el que se reporta la más alta pérdida de bosque; por lo que, la disminución en la densidad poblacional de esta especie se relacionaría con el incremento de la perturbación antrópica, sin embargo, se requerirá de más información para sustentar dicha relación. Opuesto es el caso de Akodon kotosh, que aumentó su densidad poblacional en el 2017, y a pesar de que es poca la información de su historia natural, se trata de un género común para las yungas del Perú (Pacheco et al. 2009). Por otro lado, la deforestación en las zonas altas de Carpish podría explicar el registro de T. kalinowskii en las zonas bajas, y aunque es una especie generalista (Noblecilla & Pacheco 2012), aun es necesario realizar observaciones sobre los 2500 m para confirmar su presencia. No obstante, es importante considerar que áreas abiertas producen microhábitats favorables para especies generalistas según Adler et al. (1997); lo cual explicaría que T. kalinowskii y A. kotosh actúen como especies generalistas en el sitio de estudio y sean considerados como "indicadores de perturbación" del bosque.
La diversidad resultó relativamente similar entre los años 2002 y 2017; pero se evidencia un cambio en la composición de especies, puesto que Nephelomys keaysi se registró sólo en el 2002, pero por tratarse de una especie considerada "rara" de registrar, se recomienda no compararla con otras especies.
Los cambios en la composición de especies registradas entre los años 2002 y 2017 podrían relacionarse a cambios en la estructura del bosque, debido que cada roedor cricétido desempeña un rol específico en su hábitat, como el rol en la "sucesión de los bosques" (Sunyer et al. 2013, Manson et al. 2001). Thomasomys kalinowskii es una especie herbívora, conocida por su alto consumo de semillas de Piperacea (Noblecilla & Pacheco 2012), ésta es una vegetación relacionada al bosque secundario, borde de bosque y claros (Sánchez-Coronado et al. 1990), así mismo es considerada como una especie dominante en sotobosque (Dyer & Palmer 2004); por lo que, la presencia de T. kalinowskii en las partes bajas de San Pedro de Carpish podría ser un indicador de la pérdida de bosque en esa zona. Diversos estudios de flora y fauna ya han reportado la disminución de bosque en el sitio de estudio (Young & León 1999, Pacheco & Noblecilla 2019, Beltrán & Salinas 2010) y que corrobora el Programa Nacional de Conservación de Bosques para la Mitigación del Cambio Climático: "Chinchao registra uno de los picos más altos de pérdida de bosque en el año 2017".
Por otro lado, el incremento en la densidad poblacional de A. kotosh, en relación con el aumento de perturbación del bosque, podría también considerar a esta especie como indicadora de perturbación antrópica en los bosques de Carpish. Además, si las poblaciones de esta especie llegaran a reducir drásticamente sería una señal preocupante del estado de conservación de este bosque.
Es importante resaltar la persistencia de estos roedores ante la perturbación antrópica (construcción de carreteras, agricultura y ganadería) en el sitio de estudio, lo cual podría indicar que mantienen la estructura y funciones básicas del ecosistema natural (Mena 2010); sin embargo, los cambios en la composición de especies afectan la estructura y composición del bosque. Se recomienda realizar investigaciones en la gradiente altitudinal de Carpish, además de acciones de conservación, considerando el aumento de la concentración de pérdida de bosque observado; así como también, planes de manejo adecuado para la agricultura y ganadería presentes en el área de estudio.

