











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Grandes extensiones de bosques tropicales poco perturbados pueden observarse aun en la región enmarcada por los ríos Amazonas y Ucayali en el norte y oeste, y por el río Yavarí en el este. Es una región con alta biodiversidad; por ejemplo, de árboles (ter Steege et al., 2000) y de mamíferos (Brum et al., 2017). Pero existen escasos lugares dentro de este interfluvio donde se ha aprovechado esta condición para estudiar esta biodiversidad en más detalle o para explorar y conocer más grupos de organismos, y realizar investigaciones en ecología y conservación. Uno de estos lugares es la Estación Biológica Quebrada Blanco (EBQB; Fig. 1) donde principalmente han sido estudiados los primates; pero que, por su diversidad biológica, y la existencia de una infraestructura permanente, se presta a investigaciones de muchos otros organismos y de sus interacciones ecológicas. Por eso, presentamos aquí datos sobre la EBQB con fines de informar a la comunidad científica y estimular investigaciones en biodiversidad, ecología, conservación y otras ramas de la biología en este lugar de la Amazonía peruana.

Ubicación de la EBQB, contexto geográfico-geológico y clima
La EBQB está ubicada en la cuenca del río Tahuayo, en el margen derecho de su afluente Quebrada Blanco, a 4°21’S, 73°09’W (Easting: 705305, Northing: 9518934, UTM Zone 18, WGS 84; Fig. 2a). [Quebrada Blanco anteriormente llamada río Blanco, pero por su pequeño tamaño y para evitar una confusión con el río Blanco, afluente del río Tapiche, fue renombrado Quebrada Blanco]. Está situada cerca al límite entre una terraza antigua del río Amazonas y de la Formación Pebas (Fig. 2b; Kalliola et al., 1998). Los edificios de la EBQB se encuentran al interior del bosque, a una distancia de aproximadamente 750 m en línea recta desde la orilla de la Quebrada Blanco, a aproximadamente 130 m de altitud. Está rodeado principalmente por bosque primario. Hacia el oeste, existe un gran pasto de búfalos, y hacia el sur un ex-pasto, creado en el año 1990, abandonado y regenerándose desde el 2000 (Heymann et al. 2019). En el margen izquierdo de la Quebrada Blanco, a una distancia de ca. 1 km en línea recta de la EBQB, se encuentra la comunidad de Diamante - 7 de Julio.
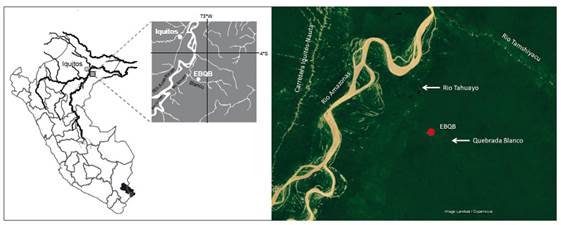
Figura 2 (a) Ubicación geográfica y (b) contexto geo-ecológico de la Estación Biológica Quebrada Blanco (EBQB). Fuente de imagen satelital: Google Earth.
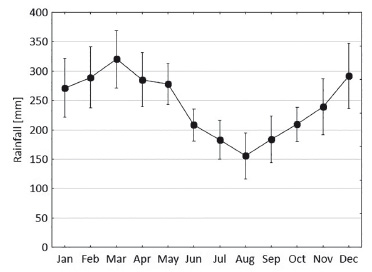
Como en toda la región, el clima es tropical-húmedo. La precipitación anual es aprox. 3000 mm, con una marcada estacionalidad de las lluvias (Fig. 3) Basado en datos entre 1997-2017 de la estación meteorológica de Tamshiyacu, 40 km al norte de la EBQB, los meses de diciembre hasta mayo reciben un promedio de >250 mm, y los de meses julio a setiembre un promedio de <200 mm.
Historia de la EBQB
Desde 1975, la zona del río Tahuayo, incluyendo su afluente Quebrada Blanco, ha sido explorada por investigadores del Proyecto Peruano de Primatología (PPP) e identificada como una zona de alta diversidad primatológica, que se presta a medidas de conservación y manejo sostenible de primates en medio natural (Tapia et al. 1990). En 1975, Castro y Soini (1977) realizaron estudios preliminares del pichico barba blanca, Saguinus mystax, en esta zona y fueron los primeros en mencionar la formación de asociaciones interespecíficas (manadas mixtas) entre este primate y el pichico común, Leontocebus nigrifrons (anteriormente conocido como Saguinus fuscicollis nigrifrons; véase Rylands et al. 2016 por el cambio de la taxonomía y la nomenclatura de los pichicos). En 1977, Soini y de Soini (1990) realizaron un estudio de la ecología poblacional de S. mystax en la zona.
En 1981, el Proyecto Peruano de Primatología (PPP) fundó una estación biológica en el margen izquierdo de la Quebrada Blanco, en el lugar donde ahora se encuentra la comunidad de Diamante - 7 de Julio. En dicha estación se realizaron estudios sobre la ecología de pichicos por la bióloga peruana Marleni Ramirez en 1981-82 (Ramirez 1989), y por los primatólogos estadounidenses Marilyn A. Norconk en 1982-83 (Norconk 1986) y Paul A. Garber en 1984 (Garber 1988a, 1988b). Por iniciativa del biólogo peruano Rogerio N. Castro Coronado, el PPP fundó una segunda estación biológica en 1984, con fines de ampliar las opciones para estudiar los pichicos. En esta segunda estación Castro Coronado realizó estudios en 1984-85 (Castro Coronado 1991) y también, durante el mismo período la bióloga peruana Tula G. Fang Monge (Fang 1990) estudió la ecología de pichicos, y este fue el origen de lo que ahora es la EBQB.
Seguidamente, en 1985-86, la EBQB fue la sede para estudios en ecología y comportamiento de pichicos de los investigadores del Deutsches Primatenzentrum (DPZ, Centro Alemán de Primates) (p.ej., Bartecki & Heymann 1990, Heymann 1990). Durante este período, también fueron colectados datos del huapo rojo, Cacajao calvus ucayalii (en esa época todavía llamado Cacajao calvus rubicundus) (Bartecki & Heymann 1987). Las observaciones del huapo rojo fueron un importante aporte para las primeras propuestas de crear una reserva en la zona (Bartecki et al. 1986); para la historia de lo que ahora es el Área de Conservación Regional Comunal Tamshiyacu-Tahuayo (ACRCTT) véase https://es.wikipedia.org/wiki/área_de_conservación_regional_comunal_Tamshiyacu-Tahuayo.
Entre 1990 y 1996, la EBQB fue usada en forma discontinua para estudios de los autores (p.ej., Heymann 1996, 2000, Tirado Herrera 1998) y de estudiantes de doctorado de Alemania e Inglaterra (Smith 1997, Knogge 1999). Sin embargo, en 1997 la EBQB comenzó a recibir fondos del DPZ; y la autora ER Tirado Herrera (ERTH) trabajó como directora de la EBQB, además de realizar sus estudios ecológicos y etológicos de las dos especies de pichicos y del tocón colorado, Plecturocebus cupreus (anteriormente Callicebus cupreus; véase Byrne et al. 2016, por el cambio taxonómico) (p.ej. Tirado Herrera et al. 2000, Tirado Herrera & Heymann, 2004). La presencia de ERTH y un asistente local garantizó un flujo continuo de información sobre grupos habituados de pichicos y tocones y el mantenimiento de la EBQB, esta continuidad además permitió tener mayor seguridad para plantear nuevos proyectos de investigación.
En 1999, la Universidad Nacional de la Amazonía Peruana (UNAP) y el DPZ firmaron una Carta de Entendimiento, la cual creó la base formal para la existencia y funcionamiento de la EBQB. Desde entonces se han realizado un gran número de estudios ecológicos y etológicos, no solo sobre primates sino también de otros organismos. Así, destacan los estudios en primates enfocados en la dispersión de semillas (p.ej., Knogge et al. 2003, Culot et al. 2011, Bialozyt et al. 2012, Heymann et al. 2019); ecología alimenticia y forrajeo (p.ej., Nickle & Heymann 1996, Heymann et al. 2000, Nadjafzadeh & Heymann 2008); ecología sensorial (p.ej., Smith et al. 2012, Nevo et al. 2016); sistemas sociales y de apareamiento (p.ej., Huck et al. 2004, Dolotovskaya et al. 2020); y comunicación olfativa (p.ej., Heymann 1998, Lledo Ferrer et al. 2011). Es así como, los estudios en la EBQB resultaron en más de 200 publicaciones y tesis (Heymann 2021).
Cabe destacar que, desde sus inicios la EBQB ha contado con el valioso apoyo de las comunidades de Chino, San Pedro y Diamante - 7 de Julio, cuyos integrantes han venido participando en los diversos estudios como asistentes de campo, ayudando tanto en la recolección de datos y de muestras biológicas (p.ej., heces de los monos) y también en la logística (p.ej., mantenimiento de la EBQB y transporte).
Hábitat, aguas y suelos
El hábitat de la EBQB está caracterizado por bosque de altura según la terminología de Encarnación (1985), con terreno que varía entre plano, con bosque de terraza y colindando con bosque de colina. A lo largo de pequeñas quebradas se encuentran zonas pantanosas, con presencia del aguaje, Mauritia flexuosa. La vegetación a lo largo de la Quebrada Blanco está caracterizada por bosque de tahuampa.
Mientras que, la Quebrada Blanco lleva agua blanca, las pequeñas quebradas dentro de la zona de estudio llevan agua de mayor transparencia o agua negra (Fig. 4). Después de lluvias fuertes, las pequeñas quebradas pueden ser dominadas por agua turbia. Los suelos consisten de ultisoles, con una gran proporción de arena blanca (hasta 75%; Pitman et al. 2008) (Fig. 5). Dentro del área de estudio existe gran variación en la fertilidad de los suelos (Fig. 6; Cárdenas Ramírez et al. 2021).

Figura 4 Una quebrada con agua clara dentro del área de estudios de la EBQB. Después de lluvias fuertes, esta quebrada lleva agua turbia. Foto © Eckhard W. Heymann.

Figura 5 En grandes partes del área de estudios de la EBQB, los suelos contienen una alta proporción de arena blanca. Foto © Eckhard W. Heymann.
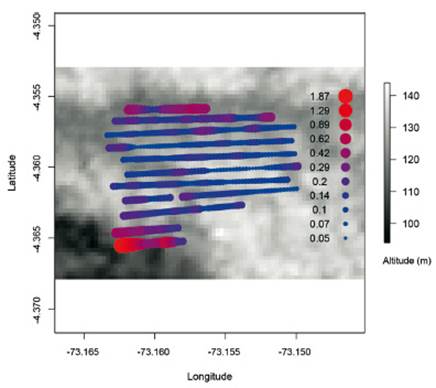
Figura 6 La fertilidad de los suelos en el área de estudios de la EBQB demuestra un gradiente desde del suroeste hacia el norte y noreste. Fuente: Cárdenas Ramírez et al. (2021). Reproducido baja Creative Commons license 4.0.
Flora
Hasta ahora han sido realizados solo dos inventarios botánicos, uno de árboles con DAP >10 cm (Dávila Cardozo & Ríos Paredes 2006), y otro de helechos y lycophytas (Cárdenas Ramírez 2019). El inventario de árboles se realizó en dos parcelas de 1 ha cada una, en bosque de terraza (Fig. 11a). Las familias dominantes fueron Fabaceae, Elaeocarpaceae, Burseraceae, Lecythidaceae y Chrysobalanaceae y los géneros Sloanea, Licania, Protium, Eschweilera y Oenocarpus, según diversidad y abundancia (Tabla 1). El inventario de helechos incluyó toda la zona de estudios de la EBQB y reveló 81 especies, entre generalistas y especialistas sea de suelos fértiles o de suelos magros (Cárdenas Ramírez 2019).
Tabla 1 Familias y géneros de árboles con DAP >10 cm de mayor diversidad y abundancia y especies más abundantes. Diversidad: # de especies; Densidad relativa: # de individuos de especie i *100 / # de individuos de todas especies en las dos parcelas. Basado en datos de Dávila Cardozo y Ríos Paredes (2006).
| Familias | Géneros | Especies | |||||
|---|---|---|---|---|---|---|---|
| Rango según: | diversidad (# de especies) | densidad relativa | diversidad (# de especies) | densidad relativa | densidad relativa | ||
| 1 | Fabaceae | Lecythidaceae | Sloanea | Eschweilera | Eschweilera coriacea | ||
| 2 | Elaeocarpaceae | Fabaceae | Licania | Licania | Oenocarpus bataua | ||
| 3 | Burseraceae | Chrysobalanaceae | Protium | Oenocarpus | Eschweilera tessmannii | ||
| 4 | Chrysobalanaceae | Euphorbiaceae | Pouteria | Protium | Iryanthera polyneura | ||
| 5 | Sapotaceae | Myristicaceae | Inga | Iryanthera | Micrandra spruceana | ||
| 6 | Lauraceae | Burseraceae | Pourouma | Micrandra | Zygia racemosa | ||
| 7 | Myristicaceae | Lauraceae | Tachigali | Sloanea | Licania reticulata | ||
| 8 | Euphorbiaceae | Arecaceae | Guarea | Tachigali | Guarea macrophylla | ||
| 9 | Rubiaceae | Sapotaceae | Eschweilera | Inga | Micrandra cf. elata / Eschweilera itayensis / Ophiocaryon manausense | ||
| 10 | Annonaceae / Moraceae | Elaeocarpaceae | Virola | Guarea | |||
Observaciones cualitativas sugieren que también existe una alta diversidad de árboles con DAP <10 cm, lianas, epífitas y hongos. Tambien, se encuentran dentro del área de estudio varias “supay chacras”, que son pequeños parches creados por la interacción entre Duroia hirsuta (Rubiaceae) y la hormiga Myrmelachista schumanni (Fig. 7). Además, la flora de la EBQB incluye numerosas especies de plantas de hábito inusual, como las myco-heterotróficas Sciaphila purpurea (Triuridaceae; Fig. 8) que solo crecen en termitarios (Franke 1999) o la liana Marcgravia longifolia (Marcgraviaceae) que presenta inflorescencias e infrutescencias a largo de su tronco desde el suelo hasta el dosel, interactuando con un gran espectro de animales nectarívoros y frugívoros en los diferentes estratos del bosque (Tirado Herrera et al. 2003, Thiel et al. en revisión).

Figura 7 Una “supay chacra” en la EBQB. El área alrededor del árbol de la especie Duroia hirsuta está mantenido libre de otras plantas por la actividad de las hormigas Myrmelachista schumanni. Foto © Eckhard W. Heymann.

Figura 8 Sciaphila purpurea es una planta myco-heterotrófica que solo crece en termitarios (Franke 1999). Foto © Eckhard W. Heymann.
Fauna
Mamíferos.- En un inventario realizado en la cercanía del caserío San Pedro, aprox. 5 km de la EBQB, Valqui (2001) registró 81 especies de mamíferos (sin murciélagos ni especies estrictamente acuáticas; Tabla 2). Dentro de los mamíferos, destacan 13 especies de primates en la zona (Tabla 3). El huapo rojo, Cacajao calvus ucayalii (Fig. 9), es una especie emblemática de la zona. En un estudio sobre las interacciones planta-animal en la liana Marcgravia longifolia, se registraron 25 especies de murciélagos (Gottstein 2018); sin embargo, en este estudio, el uso de las redes de neblina solo hasta 8 m de altura podría indicar una subestimación de la diversidad real.
Tabla 2 Número de especies de mamíferos en la zona de la EBQB, según orden. Fuente: Valqui 2001.
| Orden | # de especies |
|---|---|
| Artiodactyla | 4 |
| Carnivora | 13 |
| Didelphiomorpha | 11 |
| Perissodactyla | 1 |
| Primates | 13 |
| Rodentia | 30 |
| Xenarthra | 9 |
Tabla 3 Especies de primates de la EBQB. Nombres en paréntesis son aquellos anteriormente usados para la especie.
| Familia | Especie | Nombre común | Categoría IUCN* | Presencia en la EBQB |
|---|---|---|---|---|
| Callitrichidae | Cebuella pygmaea | leoncito | LC | permanente |
| Callitrichidae | Leontocebus nigrifrons (Saguinus fuscicollis nigrifrons, Saguinus nigrifrons) | pichico común | LC | permanente |
| Callitrichidae | Saguinus mystax | pichico barba blanca | LC | permanente |
| Cebidae | Cebus unicolor (Cebus albifrons) | machín blanco | VU | transitorio infrecuente |
| Cebidae | Sapajus macrocephalus (Cebus apella) | machín negro | LC | permanente |
| Cebidae | Saimiri macrodon (Saimiri sciureus) | fraile | LC | permanente |
| Aotidae | Aotus nancymaae | musmuqui | LC | permanente |
| Atelidae | Alouatta seniculus | coto mono | LC | (permanente)# |
| Atelidae | Lagothrix lagothricha poeppigii | choro | VU | transitorio infrecuente |
| Atelidae | Ateles chamek | maquisapa | EN | transitorio muy infrecuente |
| Pitheciidae | Plecturocebus cupreus (Callicebus cupreus) | tocón colorado | LC | permanente |
| Pitheciidae | Pithecia monachus | huapo negro | LC | permanente |
| Pitheciidae | Cacajao calvus ucayalii | huapo rojo | VU | transitorio frecuente |
* LC (Least Concern): preocupación menor; VU (Vulnerable): vulnerable; EN (endangered): en peligro
# grupos de esta especie son residentes de un aguajal a aprox. 1 km de la EBQB; muy infrecuentemente esta especie transita la zona de estudios

Figura 9 El huapo rojo, Cacajao calvus ucayalii, es una especie emblemática de la EBQB. Tiene áreas domiciliares muy grandes (12 km² o más) y por regla general anda en manadas con más de 50 individuos. Foto © Thomas Ziegler
Aves.- La EBQB es uno de los lugares de estudios de ecología comunitaria de aves en el nor-oriente peruano (p.ej., Pomara et al. 2014; Socolar & Wilcove 2019). Durante los inventarios realizados para estos estudios, se registraron alrededor de 320 especies de aves en la zona de Quebrada Blanco (Lars Pomara y Jacob Socolar, datos no publicados; Tabla 4). Destacan las familias Tyrannidae, Thamnophilidae y Thraupidae por su gran diversidad de especies.
Tabla 4 Número de especies de aves en la zona de la EBQB, según familia. Fuente: Lars Pomara y Jacob Socolar, datos no publicados
| Familia | # de especies | Familia | # de especies | Familia | # de especies |
|---|---|---|---|---|---|
| Tyrannidae | 43 | Vireonidae | 7 | Caprimulgidae | 2 |
| Thamnophilidae | 40 | Cuculidae | 6 | Hirundinidae | 2 |
| Thraupidae | 26 | Trogonidae | 6 | Momotidae | 2 |
| Psittacidae | 15 | Galbulidae | 5 | Nyctibiidae | 2 |
| Dendrocolaptidae | 14 | Ramphastidae | 5 | Ardeidae | 1 |
| Picidae | 12 | Emberizidae | 4 | Conopophagidae | 1 |
| Bucconidae | 11 | Strigidae | 4 | Corvidae | 1 |
| Accipitridae | 10 | Troglodytidae | 4 | Heliornithidae | 1 |
| Furnaridae | 10 | Turdidae | 4 | Odontophoridae | 1 |
| Icteridae | 10 | Alcedinidae | 3 | Parulidae | 1 |
| Pipridae | 10 | Apodidae | 3 | Polioptilidae | 1 |
| Trochilidae | 10 | Capitonidae | 3 | Psophiidae | 1 |
| Tinamidae | 8 | Cardinalidae | 3 | Rhinocryptidae | 1 |
| Columbidae | 7 | Cathartidae | 3 | Steatornithidae | 1 |
| Cotingidae | 7 | Cracidae | 3 | ||
| Falconidae | 7 | Formicariidae | 3 |
Anfibios y reptiles.- Un inventario rápido de 10 días reveló 51 especies de anfibios (Medina Torres et al. 2012), una cifra que aumentaría con inventarios más intensos. Lo mismo aplica para los reptiles, de los cuales se han registrado 35 especies, incluyendo 15 especies de serpientes, la mayoría de la familia Colubridae. Una especie emblemática de anfibios muy poco conocida es el Cruziohyla craspedopus (Fig. 10). Además, está presente Atelopus spumarius, especie vulnerable y miembro de uno de los géneros de anfibios más amenazados a nivel mundial (La Marca et al. 2005, IUCN 2021).

Figura 10 El sapo Cruziohyla craspedopus presenta líneas y formas irregulares en su dorso que asemejan a hojas infectadas por larvas mineros. Foto © Eckhard W. Heymann.
Invertebrados.- No se ha realizado inventarios de ningún grupo de invertebrados, aunque es altamente probable que ellos también presenten una alta diversidad de especies en la zona, incluyendo especies nuevas. Por ejemplo, Kaderka (2016) describió una nueva especie de tarántula (Theraphosidae), Cyriocosmus giganteus. También, si consideramos que entre los ortópteros (“saltamontes”) que forman parte del espectro de presas de los pichicos L. nigrifrons y S. mystax, muchas especies que se registraron solo fueron identificadas hasta género o subfamilia (Nickle y Heyman 1996, Smith 2000), esto sugeriría la presencia de especies no conocidas. Por otro lado, por primera vez una asociación inusual entre un coleóptero y el tocón colorado fue reportada (Tirado Herrera et al. 2002), lo cual podría estar relacionado con nuevas especies de coleópteros.
Infraestructura de la EBQB y acceso
La EBQB tiene tres casas: una casa grande para dormir; otra casa grande para reunirse, comer y cocinar; y una casa pequeña que sirve como “laboratorio”, con escritorios, biblioteca y almacén (Fig. 11). Las pilastras que sostienen a las casas son de cemento, las demás construcciones son de madera, y los techos de calamina. El agua de las lluvias se colecta en cuatro tanques y sirve para un baño noble y la limpieza en la cocina. Paneles solares proveen electricidad para tener luz en la noche, manejar laptops y otros aparatos pequeños, y recargar pilas. Para casos de mayor demanda de electricidad hay un generador.

Figura 11 Vista aérea de las casas de la EBQB. En primer plano se ve la casa para comer, cocinar y reuniones; a la derecha la casa dormitorio; a la izquierda el “laboratorio”. Foto © Katrin Heer.
La EBQB dispone de un sistema de trochas, cubriendo un área de aprox. 1.2 km². Las trochas están en dirección norte-sur y oeste-este, con intersecciones aproximadamente cada 100 m (Fig. 12). El sistema de trochas facilita el desplazamiento dentro del área y la ejecución de estudios que requieren diseños sistemáticos con puntos de muestreo, transectos o parcelas (p.ej., Dávila Cardozo y Ríos Paredes 2006; Heymann et al. 2014). Las trochas son angostas y cubiertas por el dosel de los árboles.
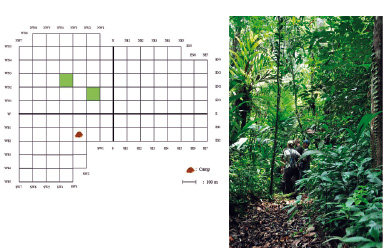
Figura 12 (a) Sistema de trochas de la EBQB. Los dos cuadrantes verdes indican las parcelas usadas en el inventario de árboles por Dávila Cardozo y Ríos Paredes (2006). (b) La mayoría de las trochas son angostas y cubiertas por el dosel.
El acceso a la EBQB es vía transporte fluvial, no hay pistas. Con deslizador, un viaje desde Iquitos demora aprox. 4 horas, pero durante la época de vaciante, el acceso con deslizador puede ser muy difícil, hasta imposible. Alternativamente, se puede viajar en lancha desde el puerto de Belén hasta el caserío Chino en el río Tahuayo. Desde Chino son 2-3 horas de viaje en embarcación menor (chalupa o peque peque), aunque puede demorar más en la época de vaciante. Desde el puerto de la Quebrada Blanco se camina 12-15 min hasta la estación.
Potenciales de investigación científica
Como se ha mostrado en los estudios de árboles por Dávila Cardozo y Ríos Paredes (2006), de helechos y lycophytas por Cárdenas Ramírez (2019), de aves por Pomara y Socolar (véase arriba), y en nuestros estudios, la EBQB tiene un gran potencial para investigaciones más allá de los estudios primatológicos.
El terreno complejo con terrazas, pendientes y zonas pantanosas, y la variación a pequeña escala de la fertilidad del suelo se presta a investigaciones a escala local de sus efectos sobre la diversidad de plantas y la composición y estructura de la vegetación. Los inventarios de Dávila Cardozo y Ríos Paredes (2006) estaban limitados a dos parcelas en bosque de terraza y solo incluyeron árboles con un DAP >10 cm. Sería de interés conocer tanto la diversidad de árboles en otros tipos de hábitats (p.ej., bosque de colina) como la diversidad de otros tipos de plantas como lianas, epífitas y de hongos. Además, el efecto de fertilidad del suelo sobre la composición de la vegetación y sobre la distribución local de las especies de plantas es otro tema interesante para desarrollar; por ejemplo, la liana M. longifolia solo se encuentra en el norte y este de la zona de estudios con baja o media fertilidad del suelo, pero no en el sureste, donde la fertilidad del suelo es elevada (datos no publicados; véase Fig. 5 para la fertilidad del suelo). La tamara, Leonia cymosa (Violaceae), presenta densidades más altas en las partes más fértiles del sureste que en el este, pero falta casi completamente en el norte (Gelmi-Candusso et al. 2019), sugiriendo la existencia de factores adicionales que variarían a escala local y determinarían la presencia y densidad de especies.
La mayoría de los animales dependen directa o indirectamente de plantas, sea como alimento (hojas, frutos, semillas, néctar, exudados, etc.) o como hábitat y refugio. En un proceso bottom-up, las diferencias en la fertilidad del suelo podrían relacionarse a la composición de las comunidades de animales, sus densidades poblacionales y al uso del espacio a través de la composición y estructura de la vegetación (véase Cárdenas Ramírez et al. 2021 para el efecto de la fertilidad del suelo sobre el tamaño del área domiciliar de pichicos).
Sobre estas preguntas abiertas, que proponemos en base a nuestros estudios y experiencias, expertos y especialistas en ecología y biodiversidad o en la biología de los diferentes taxones de animales y plantas, se podrían concebir numerosos y diversos temas de investigación en la EBQB.

