











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
The Tropical Andes region is an important centre of South-American biodiversity (Hoorn et al. 2010), containing hotspots for plant and animal biodiversity (Myers et al. 2000, Young et al. 2002, Swenson et al. 2012). In this region, the transition zone between the upper montane forest and wet grasslands (the treeline ecotone) is of particular interest due to its outstanding species and functional diversity (Bader et al. 2007, Ramírez et al. 2009, Hofstede et al. 2014, Peters et al. 2014, Llambí 2015) with high endemism rates for many taxa of Lepidoptera (Willmott et al. 2001, Lamas 2003, Pyrcz 2004, Hall 2005, Ignatov et al. 2011, Pyrcz et al. 2014, Sublett et al. 2019), including Lycaenidae (e.g. Prieto & Bálint 2007, Prieto & Rodríguez 2012).
In 2016, during an entomological expedition carried out to the high mountains of the Tropical Andes of northern Peru, one female adult of an undetermined Eumaeine species was collected, and provisionally labelled as “Lycaenidae sp.” in the scientific collection of the Museo de Historia Natural, Universidad Nacional de San Agustín de Arequipa (MUSA). Recently, we examined this specimen and consider that it belongs to the genus Podanotum Torres & Johnson, 1996 (Lycaenidae: Eumaeini) due to its wing pattern and female genitalia characters. Podanotum currently contains eleven described species (Robbins 2004, Bálint & Wojtusiak 2006, Bálint & Attal 2007, Prieto & Rodríguez 2012) distributed from Venezuela to southern Peru, along the upper montane forests of the Tropical Andes (Bálint & Wojtusiak 2006, Lamas et al. 2021).
Torres et al. (1996) enumerated the morphological diagnostic characters of Podanotum, including a bipartite habitus with sclerotized terminal element in the female genitalia. Moreover, it was recorded that sexual dimorphism in the genus is limited, male and female phenotypes being very similar to each other (Bálint & Wojtusiak 2006, Bálint & Attal 2007, Prieto & Rodríguez 2012). The single specimen recently collected in Peru represents a hitherto unknown phenotype. Therefore, the aim of this study is to describe formally a new species of Podanotum, based on one female specimen collected in northern Peru at 3200 m elevation and to place it amongst congeners with the help of a key to identification.
Material and methods
Material examined. One female specimen of Podanotum was examined from the Museo de Historia Natural, Universidad Nacional de San Agustín de Arequipa, Arequipa, Peru (MUSA). Photographs of all type specimens of Podanotum taxa were examined from images available in Bálint & Wojtusiak (2002), Prieto & Rodríguez (2012), and Warren et al. (2017).
Morphological analysis. The abdomen was removed from the specimen and soaked in a 10% KOH solution for ten minutes. Subsequently, the abdomen was preliminarily cleaned out of soft tissue in water in order to expose the genitalia, afterwards stained with chlorazol black in order to identify the soft parts. Water was extracted from the dissected genitalia by using 90% and 95% ethanol solutions. Wing venation and genitalic terminology follows Nicolay (1977) and Klots (1970) respectively. Measurements are in millimetres and were made from photographs of specimen taken next to a scale and magnified on a computer screen. Photographs of adult were taken with a Nikon D610 digital camera through a Nikkor 105 mm f/2.8G AF-S VR Micro lens; photographs of genitalia were taken with a camera Nikon DS-Ri2 through a Nikon SMZ25 stereomicroscope. Images were assembled and edited in Nikon NIS-Elements and Photoshop CS5.1.
Genomic DNA was extracted from abdominal tissue, following the procedures described by Huanca-Mamani et al. (2015). A fragment of the COI gene was amplified by polymerase chain reaction (PCR) with the primers LEP-F1 and LEP R1 (Hebert et al. 2004). PCR reactions were performed in a final volume of 20 μL. Each reaction contained 1 μL of DNA extract, 10 ρmoles of each primer, 2.5 mM of each dNTP, 2 mM MgCl2, 1X PCR buffer (KCl), 1 unit of Taq DNA polymerase (Thermo Scientific) and sterile distilled water. The amplification program was 5 min at 94 °C, 35 cycles of 30 s at 94 °C, 30 s at 47 °C, 1 min at 72 °C, and a final elongation step of 10 min at 72 °C. Three μL of each PCR product was visualized on 1.5% agarose gel stained with gel-red (Biotium). Reactions containing fragments of the expected size were purified and directly sequenced by a commercial facility (Macrogen, South Korea).
The following abbreviations are used:
MUSM-Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Peru.
MUSA-Museo de Historia Natural, Universidad Nacional de San Agustín de Arequipa, Peru.
DFW-Dorsal forewing
DHW-Dorsal hindwing
VFW-Ventral forewing
VHW-Ventral hindwing
Results
Podanotum pajaten Farfán, Cerdeña & Bálint sp. nov.
Type material. - Holotype, female: PERU, LL [La Libertad], Pataz, Parque Nacional Abiseo, Puerta del Monte, 3200 m, 0741/7727 [7°41´S, 77°27´W], 19.v.2016, Leg. J. Farfán (MUSA) [will be deposited in MUSM]. Genbank accession number: OK484465.
Type locality. - Peru, La Libertad, Pataz, Puerta del Monte, 7°41’S, 77°27’W, 3200 m.
Diagnosis. - Podanotum pajaten sp. nov. can be distinguished from the other known congeneric species by the following combination of characters: (1) dorsal wing surface pale blue colored (vs. brown color in P. andrewneildi Bálint, 2001 and P. melanissimum Bálint & Wojtusiak, 2006), (2) conspicuous discoidal line on the VFW (vs. absent in the other species except P. glorissimum, with metallic green dorsal wing surface and P. andrewneildi, with brown dorsal wing surface), (3) hindwing tailed (vs. absent in P. glorissimum), and (4) ostium bursae distally toothed (vs. untoothed in P. glorissimum).
Description. -
Female (Fig. 1). Forewing length 12.6 mm (n=1).
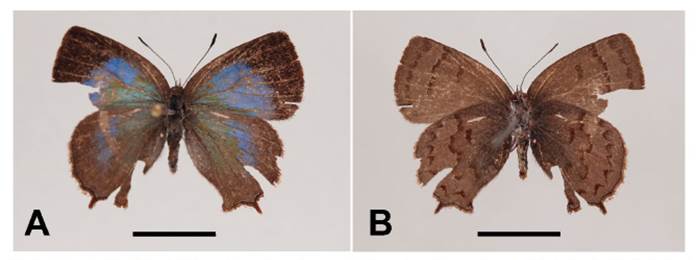
Figure 1 Adult female of Podanotum pajaten sp. nov., Holotype. A. dorsal view; B. ventral view. Scale bar: 10 mm.
Head: Primarily black with brown hairs dorsally, brown eyes with white hair-like scales around, hairy palpus with white hair-like scales, antenna striped white and black.
Thorax: Wings dorsally pale blue. DFW with broad blackish border (approx. 1.5 mm in width) to apical and outer margins. Hindwing rounded, DHW dark grayish brown border to outer margins with thin marginal black line, presence of slight tornal lobe and short brown tail. Fringe on both wings formed of long light brown scales. Wings ventrally dark grayish brown. VFW with discal line, postmedian line terminates posteriorly at vein Cu2, and submarginal line terminates posteriorly at vein A2, all lines brown. VHW with discal line, postmedian line irregular terminates posteriorly at vein Cu2, and submarginal line with lunular spots (basally reddish brown, distally black scales) all lines reddish brown.
Abdomen: Blackish dorsally and light grey ventrally, with long white hairs.
Female genitalia (Fig. 2): Fluted lamella postvaginalis, ductus bursae distally sclerotized, prominent, toothed, anterior section of ductus bursae large and thin tube weakly sclerotized. Corpus bursae with two very large and shield-like signa, inwardly toothed in lateral view; anal papillae enlarged and elliptic with apophyses about half the size of the ductus bursae length, with the proximal tip bent.

Figure 2 Podanotum pajaten sp. nov., female genitalia in lateral view; details of signa on upper left corner. Scale bar: 1 mm.
Male: Unknown.
Distribution: Podanotum pajaten is known only from the type locality in Río Abiseo National Park, at the treeline ecotone, approximately 100 km south of the type locality of the closest distributed species, P. glorissimum.
Etymology: The specific epithet is a masculine noun in apposition, derived from the “Gran Pajatén” archaeological site, which occurs in the Río Abiseo National Park, near where this new species was found.
Host plant: Unknown.
Remarks. - This new species is placed in the genus Podanotum (type species: Podanotum clarissimus Hall, Willmott & Johnson, 1996, by original designation) by the following combination of diagnostic characters: dorsal wing blue green, VHW with submarginal band of blackish-brown lunules or chevrons, female genitalia with sclerotized terminal element (Torres et al. 1996), and VFW with postmedian line ending posteriorly at vein Cu2 (Robbins & Busby 2015).
Discussion
According to Prieto & Rodriguez (2012), the wing pattern of Podanotum is not geographically variable within species, so we discarded that this new species could be a variation or subspecies of another population of Podanotum previously described, and we are convinced that the species status of P. pajaten is fully justified; despite the description being based on a single somewhat damaged female specimen, it contains enough characters to differentiate it from other species of the genus, and considering that within the genus there is very little sexual dimorphism, it is probable that the males of P. pajaten are very similar to the female described herein.
In the Neotropical butterfly checklist (Robbins 2004) Podanotum was placed next to Penaincisalia Johnson, 1992 (type species: Thecla ? culminicola Staudinger, 1894, by original designation) in the Micandra Section of Eumaeini, based on genitalia and wing pattern. In a preliminary phylogenetic analysis with molecular data, Podanotum joined with Rhamma Johnson, 1992 (type species: Thecla oxida Hewitson, 1870) (Quental 2008). Subsequently, Robbins & Busby (2015) in a phylogenetic analysis with morphological evidence indicate that Podanotum is closely related to Lathecla Robbins, 2004 (type species: Thecla latagus Godman & Savin, 1887). Therefore, further phylogenetic analysis, integrating morphological and molecular characters, is still required to determine the phylogenetic position of Podanotum.
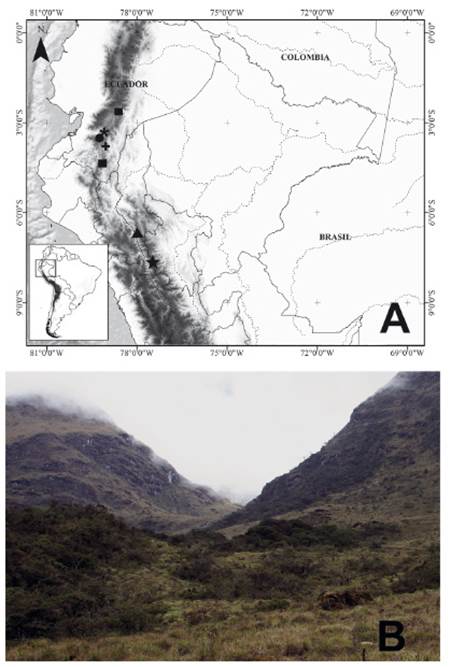
Figure 3 Distribution and habitat of Podanotum pajaten sp. nov. A. Distribution map of Podanotum species in Peru and Ecuador. P. pajaten sp. nov. = star, P. glorissimum = triangle, P. metallicus = circle, P. melanissimum = asterisk, P. aquaphilum = square, P. magnificum = cross; B. Type locality of P. pajaten sp. nov.
Key to Podanotum species with indications of country occurrence


