











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El patrón de actividad horaria de un animal hace referencia a la forma en la que estos organismos regulan sus periodos de actividad en función a un ciclo de sueño-vigilia de 24 horas (Bennie et al. 2014). Este patrón está asociado a un conjunto de adaptaciones fisiológicas y morfológicas (Kenagy & Vleck 1982) e influenciado por factores extrínsecos propios de su entorno (Ikeda et al. 2016). Dentro de estos factores se encuentran, por ejemplo, la duración del día, la iluminación lunar, las interacciones depredador-presa o por competencia, la actividad antrópica, entre otras (Ikeda et al. 2016, Noor et al. 2017). En general, el patrón de actividad puede ser categorizado como: diurno, nocturno, crepuscular o catemeral (Bennie et al. 2014). Los patrones de actividad horaria de muchos mamíferos neotropicales son conocidos o han sido reportados durante los últimos años, debido al desarrollo de métodos no invasivos como las cámaras-trampa (Lee et al. 2019). Sin embargo, la variación geográfica o los detalles específicos asociados a su interacción dentro de ecosistemas amazónicos aún se desconocen o permanecen poco estudiados (Blake et al. 2012, Bennie et al. 2014).
De los países de Centro y Sudamérica, el Perú es el segundo con mayor cantidad de reservas de gas natural (PROMIGAS 2022). Las principales actividades de extracción de este recurso se centran en la región amazónica, especialmente en los lotes correspondientes al yacimiento de Camisea (García & Vasquéz 2004), ubicado al sureste del territorio, en el departamento de Cusco. El gas de Camisea es transportado desde la Amazonía hasta el litoral costero del Pacífico, mediante una red de tuberías que atraviesan diferentes localidades y áreas, entre las que destacan algunas áreas naturales protegidas como: el Parque Nacional Otishi y la Reserva Comunal Machiguenga (Sagua 2018). Las empresas a cargo de la extracción y transporte de este gas realizan periódicamente evaluaciones de impacto ambiental, como parte de los requisitos de operación impuestos por el gobierno peruano a través de sus organismos de control y fiscalización (Palomares & Novoa 2013). Este tipo de evaluaciones ha permitido generar información sobre la diversidad biológica que habita el área circundante a la tubería, lográndose conocer aspectos de la ecología de varias especies que habitan en zonas remotas o de difícil acceso (Rodríguez 2013).
Desde hace casi 20 años, con el inicio de la operación comercial, se han realizado labores de transporte del gas de Camisea dentro de la Reserva Comunal Machiguenga (Dammert & Molinelli 2006). A pesar de este considerable intervalo de tiempo, poca información relacionada a la fauna de la zona ha sido publicada sobre la reserva. Entre estas publicaciones tenemos la de Costa et al. (2018), quienes evaluaron el uso de aves y mamíferos. Otro aporte fue presentado por Repsol Exploraciones Perú (2016) en un libro donde se describen las características físicas y biológicas de la reserva Machiguenga. Por lo anterior, y considerando el compromiso socioambiental de la Compañía Operadora de Gas S.A.C. (COGA) y el interés del Servicio Nacional de Áreas Naturales Protegidas (SERNANP), en el presente trabajo se analiza la información de un año de evaluación de los patrones de actividad de los mamíferos que habitan en la zona circundante al tramo de la tubería de transporte de gas que atraviesa la Reserva Comunal Machiguenga.
Material y Métodos
Área de estudio. El estudio se desarrolló dentro del área de influencia de la Reserva Comunal Machiguenga (RCM), ubicada en el flanco oriental de la Cordillera de Vilcabamba, en el distrito de Megantoni, departamento de Cusco, Perú (Figura 1). Esta región se caracteriza por presentar un clima tropical, con una temperatura anual media de 25.8 °C y con valores precipitación que varían de 1600 a 2000 mm al año (Costa et al. 2018). De acuerdo con el mapa de clasificación de coberturas vegetales del MINAM (Ministerio del Ambiente 2015), la evaluación se desarrolló en zonas de Bosque de montaña basimontano y Bosque de montaña basimontano con Paca. La instalación de cámaras se realizó alrededor del ducto de la línea transportadora de gas natural de Camisea, entre el kilómetro 56 y 57 de esta, la misma que corresponde al lote 57 de exploración y explotación de hidrocarburos autorizada por el gobierno peruano. El relieve de la zona se caracteriza por poseer colinas, lomadas y terrazas, con pendientes que varían de moderadas a muy empinadas.
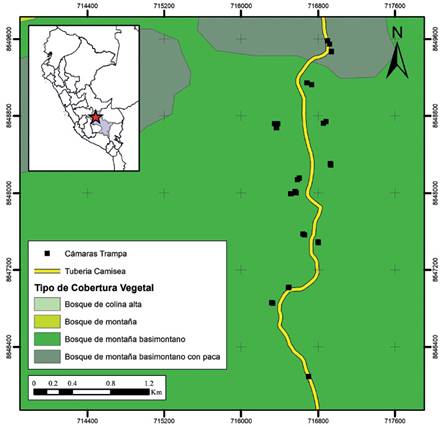
Figura 1 Mapa de ubicación de la zona evaluada y disposición de las cámaras trampa alrededor de la línea transportadora de gas que atraviesa la Reserva Comunal Machiguenga.
Evaluación fotográfica. Para el registro de los horarios de actividad de los mamíferos circundantes en el área, se utilizaron 28 cámaras trampa de la marca RECONYX Hyperfire2 repartidas en 14 puntos de evaluación que operaron desde el 07 de febrero del 2020 al 23 de enero del 2021. Las cámaras se instalaron a 60 cm del suelo y fueron configuradas para registrar eventos de detección cada cinco segundos. Luego del periodo de evaluación, las tarjetas de memoria se removieron y las fotografías documentadas se procesaron mediante el software Camera base 1.7 (Tobler 2007). La determinación de las especies se realizó a partir de guías especializadas para carnívoros (Emmons y Feer 1990), ungulados (Wilson y Mittermeier 2011) y roedores (Wilson et al. 2016). La taxonomía utilizada sigue la propuesta de Pacheco et al. (2021) para los mamíferos del Perú.
Patrones de actividad horaria. Para obtener registros independientes de cada especie, tanto temporal como espacialmente, se realizó una selección previa. A nivel temporal, los registros por especie y por estación que ocurrieron con una diferencia de al menos 60 minutos se consideraron como eventos independientes. A nivel espacial, cuando dos registros ocurrieron en un rango de tiempo menor a una hora y en estaciones cercanas, con una distancia menor al diámetro promedio calculado del home range de la especie analizada (Apéndice 1), estos se consideraron como un sólo evento independiente (Sollmann 2018). Para los análisis de actividad sólo se consideraron especies que tuvieron un mínimo de 100 eventos independientes (Lashley et al. 2018).
Los tiempos del amanecer y el atardecer fueron calculados en R (R Core Team 2022), con el paquete suncalc (Thieurmel y Elmarhraoui 2019). La duración del día, el tiempo de diferencia entre el atardecer y el amanecer, varió entre 11 y 13 horas dado que la evaluación comprende un periodo de varios meses. Por este motivo se estandarizó la hora de los registros a un día y noche de 12 horas teniendo un atardecer a las 18:00 h y un amanecer a las 6:00 h (hora estándar):
Hora estándar = 6 + 12*(hora del registro - hora del amanecer)/(duración del día)
Los patrones de actividad se generaron mediante las funciones de densidad de Kernel (Frey et al. 2017), a partir de los datos estandarizados. Los registros horarios se convirtieron a radianes, formando parte de una distribución circular continua. Siguiendo las recomendaciones de Ridout y Linkie (2009), se utilizó el parámetro Kmáx = 3 para obtener estimaciones robustas del ancho de banda óptimo tanto para distribuciones de actividad unimodales como para bimodales. Para los patrones de actividad mensual sólo se consideraron 16 cámaras, las cuales se mantuvieron activas durante al menos 297 días de tal modo que la mayoría de los meses del año tuvieran un esfuerzo de muestreo similar.
Resultados
Se obtuvieron un total de 14723 fotografías pertenecientes a 25 especies de mamíferos, de los cuales 9 se encuentran dentro alguna categoría de amenaza según la legislación peruana (DS N.º 004-2004-MINAGRI, Tabla 1). Debido a la dificultad en la distinción entre especies del mismo género con actividad nocturna, como el caso del armadillo de nueve bandas (Dasypus novemcinctus) y el armadillo de Pastaza (Dasypus pastasae), o como en el caso del venado colorado (Mazama americana) y el venado gris (Mazama nemorivaga), se identificaron estos individuos solo hasta el nivel de género. Del total de registros, se encontró que las siguientes especies de mamíferos mayores y medianos terrestres superaron los 100 eventos independientes (Tabla 1): el “añuje de Kalinowskii” (Dasyprocta kalinowskii), el “majaz” (Cuniculus paca), los “armadillos bandeados” (Dasypus spp.), los “venados” (Mazama sp.), el “tapir” (Tapirus terrestris), y la “tayra” (Eira barbara), con los cuales se procedió con los análisis de actividad.
Tabla 1 Lista de especies de mamíferos detectados alrededor de la línea transportadora de gas de Camisea que atraviesa la Reserva Comunal Machiguenga.
| Orden | Familia | Especie | Nombre común | N° de fotos con registros | N° de Eventos independientes | Categoría D.S. 004-2014 |
|---|---|---|---|---|---|---|
| Artiodactyla | Cervidae | Mazama americana | Venado colorado | 465 | 51 | DD |
| Mazama sp. | Venado | 134 | 156 | |||
| Carnivora | Felidae | Herpailurus yagouaroundi | Yaguarundi | 58 | 8 | |
| Leopardus pardalis | Ocelote | 293 | 58 | |||
| Leopardus wiedii | Margay | 89 | 21 | DD | ||
| Panthera onca | Jaguar | 15 | 1 | NT | ||
| Puma concolor | Puma | 384 | 46 | NT | ||
| Mustelidae | Eira barbara | Tayra, Manco | 546 | 154 | ||
| Procyonidae | Nasua nasua | Coati | 245 | 36 | ||
| Ursidae | Tremarctos ornatus | Oso de anteojos | 29 | 5 | VU | |
| Cingulata | Chlamyphoridae | Cabassous unicinctus | Armadillo de cola desnuda | 8 | 2 | |
| Dasypodidae | Dasypus sp. | Armadillo bandeado | 66 | 146 | ||
| Didelphimorphia | Didelphidae | Didelphis marsupialis | Zarigüeya | 8 | 1 | |
| Metachirus nudicaudatus | Raposa marrón de cuatro ojos | 76 | 19 | |||
| Perissodactyla | Tapiridae | Tapirus terrestris | Tapir, Sachavaca | 5285 | 177 | NT |
| Pilosa | Myrmecophagidae | Tamandua tetradactyla | Oso hormiguero amazónico | 39 | 9 | |
| Primate | Atelidae | Ateles chamek | Mono araña | 12 | 5 | EN |
| Cebidae | Cebus cuscinus | Machin blanco | 3 | 1 | ||
| Sapajus macrocephalus | Machin negro | 3 | 1 | |||
| Rodentia | Cuniculidae | Cuniculus paca | Picuro- majaz | 2023 | 284 | |
| Dasyproctidae | Dasyprocta kalinowskii | Aguti | 1532 | 273 | DD | |
| Myoprocta pratti | Acuchí | 3 | 1 | |||
| Echimyidae | Proechimys sp. | Rata espinoza | 1519 | 349 | ||
| Sciuridae | Hadrosciurus spadiceus | Ardilla roja | 838 | 179 | ||
| Microsciurus flaviventer | Ardilla enana amazónica | 1050 | 180 | DD |
Las especies D. kalinowskii y Eira barbara mostraron mayormente actividad diurna; mientras que, C. paca, Mazama spp., Dasypus spp. y T. terrestris presentaron mayormente actividad nocturna. Las especies diurnas mostraron un patrón unimodal con picos de actividad a las 16:00 h (E. barbara) y las 17:00 h (D. kalinowskii). Las especies nocturnas mostraron diferentes patrones; la actividad de Dasypus spp. y T. terrestris se ajusta más a una distribución bimodal con picos máximos cercanos a los periodos crepusculares, siendo especialmente más pronunciados en Dasypus spp. entre las 20:00 h y las 4:00 h, tendiendo a tener menor actividad durante la medianoche; la actividad de Mazama se ajusta más a una distribución unimodal, con un pico máximo alrededor de las 20:30 (no tan pronunciado). Por otra parte, C. paca se ajusta a una distribución trimodal teniendo el pico más pronunciado posterior al atardecer (~ 19:30 h) y un período de alta actividad con picos a la medianoche y previo al amanecer (~ 4:00 h) (Figura 2).
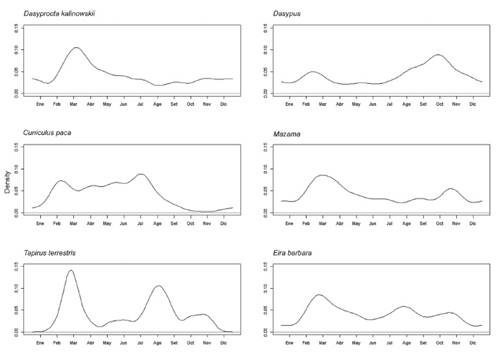
Respecto a los patrones mensuales, las especies D. kalinowskii, Mazama spp. y E. barbara mostraron mayor actividad durante la época húmeda (febrero - marzo), mientras que Dasypus spp. presentaron una mayor actividad durante la transición de la época seca a húmeda (setiembre - octubre). Por otra parte, T. terrestris mostró un pico de actividad en época húmeda y otro en época seca, y C. paca sólo no estuvo activo durante la transición entre la época seca y época húmeda, así como los primeros meses húmedos (diciembre y enero) (Figura 3).
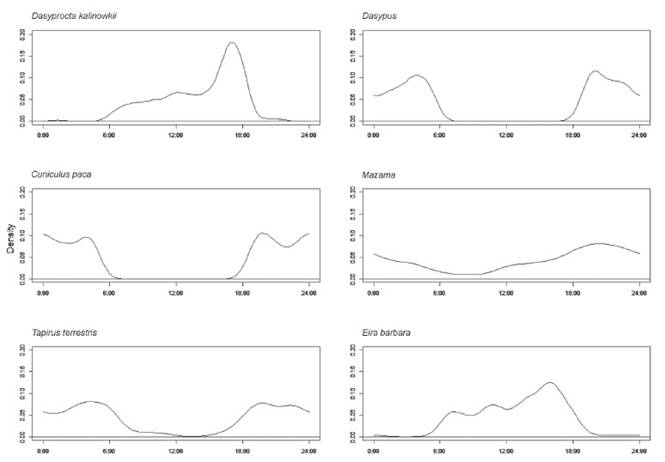
Figura 2 Patrón de actividad horario de los mamíferos alrededor de la tubería transportadora de gas de Camisea en la Reserva Comunal Machiguenga.
Discusión
A pesar de que Perú es el segundo país con mayor diversidad de mamíferos en el Neotrópico (Pacheco et al. 2021), se tienen escasos estudios sobre patrones de actividad temporal en escala horaria (e.g. Tobler 2009, Jiménez et al. 2010, Kolowski 2010, Ayala et al. 2021). En esta investigación se presenta el patrón de actividad de seis taxones de mamíferos terrestres: T. terrestris, Mazama spp., D. kalinowskii, C. paca, E. barbara y Dasypus spp., las últimas cuatro reportadas por primera vez dentro de un área natural protegida con actividad antrópica supervisada (transporte de gas) en Perú. En particular, con esta investigación se contribuye al entendimiento de los patrones de actividad de D. kalinowskii, una especie endémica del Perú categorizada en “Datos Insuficientes”, de la cual se desconoce gran parte de su historia natural.
De acuerdo con la gran cantidad de registros independientes (N = 253), y contrario a lo sugerido por Mares y Ojeda (1981), se encontró que D. kalinowskii presenta un claro patrón de actividad diurna. Aunque en general muchos aspectos de la ecología de esta especie son desconocidos (Patton & Emmons 2015), está mantendría un patrón de actividad similar al de otras especies dentro del género, como D. punctata (Suselbeek 2009) o D. azarae (Cid et al. 2015). Lambert et al. (2009) sugirieron que las especies de Dasyprocta organizan sus patrones de actividad con la intención de reducir el riesgo de exposición a depredadores, así como evadir las horas de mayor costo energético durante el día. Considerando que dentro de la RCM encontramos al ocelote, Leopardus pardalis, un felino de hábitos nocturnos mencionado como principal depredador de este tipo de roedores (Aliaga-Rossel et al. 2006, Moreno et al. 2006) y tomando en cuenta la hipótesis de alocación de riesgos (Lima 1998), es posible que D. kalinowskii mantenga hábitos diurnos pues representa el periodo de menor riesgo de depredación. Por otra parte, si bien reportamos un patrón unimodal para D. kalinowskii con un pico de actividad entre las 14:00 h y las 18:30 h, es posible que factores como la temperatura pueden alterar este patrón de conducta, similar a lo documentado en D. azarae que muestra un patrón unimodal en temperaturas bajas y un patrón con tendencia bimodal para temperaturas altas (Cid et al. 2015). Suselbeek (2009) señaló que la disponibilidad de alimento es un factor que influye en los períodos de actividad de Dasyprocta spp., por lo que es posible que la variación en la abundancia de frutos, estrechamente relacionada con la época más lluviosa en los bosques tropicales (Dunham et al. 2018) como los de la RNC, sea una de las principales causas de estas fluctuaciones mensuales.
En el caso del C. paca, se encontró un patrón de actividad nocturno, coincidiendo con la mayoría de otros registros para bosques de llanura amazónica (Michalski & Norris 2011, Leuchtenberger et al. 2018, Blake et al. 2012). No obstante, los picos de actividad horaria de esta especie parecen variar dependiendo de la zona evaluada. El patrón en la RCM muestra mayor actividad durante las primeras horas de la noche, entre las 18:00 h y 20:00 h, durante la medianoche entre 0:00 h y 1:00 h, y durante las horas previas al amanecer, entre las 4:00 h y las 6:00 h, similar a lo reportado en bosques de llanura amazónica en el este de Ecuador (Blake et al. 2012); mientras que, en la región suroeste de la amazonia brasileña se observa un solo pico de actividad entre las 2:00 h y las 4:00 h (Leuchtenberger et al. 2018). Debido a que se reconoce la plasticidad ecológica de C. paca y la poca influencia de variables abióticas sobre su actividad nocturna (Michalski & Norris 2011), es posible que sus patrones horarios estén relacionados con factores bióticos, como la presencia de potenciales depredadores en la zona (e.g. Panthera onca, Puma concolor) o la exclusión competitiva horaria con otros roedores medianos (e.g. Dasyprocta, Blake et al. 2011).
Con respecto a T. terrestris, esta presentó un patrón de actividad nocturno, como se ha reportado previamente para la región sureste de la amazonia (Wallace et al. 2012, Cruz et al. 2014). Aunque se sabe que la especie también puede presentar hábitos catemerales (Oliveria-Santos et al. 2009), nuestros registros diurnos fueron eventos ocasionales en comparación con la gran cantidad de registros nocturnos. Los picos de actividad horarios observados en la RCM muestran una tendencia bimodal, similar a lo reportado en la zona del gran Madidi-Tambopata (Wallace et al. 2012) y los bosques Atlánticos en la zona argentina de Iguazú (Cruz et al. 2014). No obstante, se ha observado que los picos de actividad pueden variar dependiendo de la zona, como el patrón trimodal observado en los bosques lluviosos Atlánticos de la Reserva Vale Natural en Brasil (Ferreguetti et al. 2017). Por otra parte, el patrón de actividad mensual de esta especie en la RCM muestra mayor incidencia en los meses de febrero-marzo y agosto-setiembre, de manera similar a lo reportado en bosques de llanura amazónica al este de Ecuador (Blake et al. 2012); razón por la cual, es posible que estos picos de incidencia estén influenciados por variaciones relacionadas con los cambios estacionales en la precipitación y temperatura de la región.
En el caso de los armadillos bandeados (Dasypus spp.), se evidenció que en la RCM presentaron un patrón de actividad nocturno, similar a lo reportado tanto para D. novemcinctus como D. pastasae en otros bosques amazónicos de la región neotropical (Norris et al. 2010, Blake et al. 2012, Aya-Cuero et al. 2021). Aunque se sabe que algunos armadillos presentan una plasticidad considerable en su actividad alrededor del día (DeGregorio et al. 2021), en regiones neotropicales, el patrón de actividad diaria parece estar relacionado inversamente con los picos de temperatura (Bernardes et al. 2015). Dado que los armadillos son considerados homeotermos imperfectos (McNab 1985), se ha sugerido a la variación en la temperatura como factor clave en la regulación de su actividad diaria, incluso más que la luz disponible (Bernardes et al. 2015). No obstante, también es importante resaltar la edad como posible otro factor de variación, pues se ha reportado diferencias en el patrón de actividad entre juveniles y adultos, siendo los primeros más activos durante el día (McDonough y Loughry 1997). Con respecto al patrón de actividad mensual, notamos que en la RCM existe mayor incidencia de registros durante los meses de septiembre-octubre, transición hacia la temporada más lluviosa en la región amazónica del sur del Perú, por lo que es posible que la precipitación influya también en sus patrones de actividad.
Respecto a los venados (Mazama spp.), la mayoría de los registros diurnos corresponden al venado colorado M. americana, mientras que durante el periodo nocturno se tuvo una cantidad considerable de registros indeterminados (N=156). Considerando que la similaridad morfológica entre especies del género Mazama ha causado errores de determinación (Duarte et al. 2008), se optó por analizar los venados a nivel de género en lugar de descartar los registros dudosos (Srbek-Araujo et al. 2019). Sin embargo, la ausencia de registros diurnos del venado gris M. nemorivaga (un congénere amazónico) y el patrón de actividad principalmente nocturno sugieren que únicamente M. americana ocurre en el área de estudio. El patrón de actividad horaria de Mazama difiere considerablemente con un estudio realizado en el bosque atlántico brasileño donde la actividad es principalmente diurna, teniendo un pico entre las 7:00 h y 8:00 h, rango donde si se tuvieron registros diurnos de la especie Mazama gouazoubira (Srbek-Araujo et al. 2019). Un estudio previo sugiere que la sincronización del ciclo circadiano en el género Mazama se encuentra principalmente determinada por una restricción filogenética, en la cual las especies del “clado gris” son diurnas y las especies del “clado rojo” son nocturnas (de Oliveira et al. 2016). No obstante, es importante mencionar que la filogenia del género Mazama es compleja y ha tenido modificaciones recientes (Bernegossi et al. 2022, Cifuentes-Rincon et al. 2020, Peres et al. 2021). Asimismo, el patrón de actividad unimodal y principalmente nocturno es más similar a los reportes de estudios realizados en la amazonia para M. americana, en comparación con el patrón diurno para M. nemorivaga (Tobler et al. 2009, de Pinto et al. 2022). No obstante, el pico de actividad posterior al atardecer (~ 20:30 h) si difiere de los picos de actividad reportados previo al amanecer (~4:00 h y 5:00 h) (Tobler et al. 2009, de Pinto et al. 2022).
Respecto a E. barbara, el patrón de actividad principalmente diurno es consistente con la revisión geográfica más reciente para esta especie (Villafañe-Trujillo et al. 2021) así como otros estudios previos realizados en el bosque atlántico de Brasil (Massara et al. 2016), Colombia y Costa Rica (Gonzales-Maya et al. 2015). Estudios sobre la interacción entre especies depredadoras indican que el comportamiento diurno de esta especie se puede explicar parcialmente por la segregación temporal con especies competidoras nocturnas como los tigrillos (género Leopardus) y con especies “depredadores top” que son principalmente catemerales (e. g. P. concolor y P. onca) (Botts et al. 2020). Eira barbara también presenta variación latitudinal en su patrón de actividad diurna, tendiendo a ser unimodal en latitudes cercanas al Ecuador con un pico pronunciado cercanos al mediodía (Villafañe-Trujillo et al. 2021). En la RCM, el principal pico de actividad es previo al atardecer por lo que posiblemente responda a ciertas condiciones estacionales teniendo en cuenta que posee mayor actividad durante febrero y abril (meses de época lluviosa) (Figura 3). Se ha reportado que E. barbara puede presentar picos más pronunciados en la mañana durante la época seca y al ocaso durante la época lluviosa (Gonzales-Maya et al. 2015).
En el Perú se han desarrollado pocos estudios relacionados con el periodo de actividad de mamíferos terrestres. Dos investigaciones se llevaron a cabo en la zona norte del país, una en los bosques nublados de Cajamarca sobre Cuniculus taczanowskii (Jiménez et al. 2010) y otra en los bosques de llanura amazónica de Loreto sobre Leopardus pardalis (Kolowski, 2010). Mientras que, en la zona sur, Tobler (2009) desarrolló un estudio en la llanura amazónica de Madre de Dios, donde reporta información sobre M. americana, M. gouazoubira (actualmente M. nemorivaga), Tayassu pecari, Pecari tajacu y T. terrestris. Nuestra investigación corresponde al cuarto estudio realizado en el país, en la zona sureste de la Amazonía peruana, presentando información sobre géneros de roedores (Dasyprocta y Cuniculus) y ungulados (Mazama y Tapirus) de importancia cinegética (Costa et al. 2018), así como el de un carnívoro mediano poco estudiado (E. barbara). Dado el escaso número de estudios sobre patrones de actividad de mamíferos en la selva peruana, se resalta la necesidad de documentar más información de este tipo de comportamientos en distintas regiones del país, especialmente en departamentos como Ucayali y Puno, donde existe una gran diversidad de especies y presión a sus poblaciones por actividades antrópicas (Pacheco et al. 2011, Ruelas et al. 2016).
En conclusión, se encontró que, alrededor de la línea transportadora de gas de Camisea dentro de la RCM, las especies D. kalinowskii y E. barbara presentan un patrón de actividad diurno; mientras que, C. paca, T. terrestris, Dasypus sp. y Mazama sp., presentan un patrón de actividad principalmente nocturno. Se sugiere que estos patrones pueden estar influenciados por distintos factores (e.g. la temperatura, disponibilidad de alimento, presencia de depredadores-presas, etc.). Los reportes de esta región de la Amazonía contribuirán al entendimiento de la ecología de estas especies en este tipo de ecosistema. Dada la variación mensual observada en la incidencia de las especies en nuestro estudio, se consideró necesario destacar que, en acuerdo con lo sugerido por Blake et al. (2012), las investigaciones relacionadas con actividad horaria en mamíferos neotropicales deben desarrollarse en un periodo largo de evaluación, de al menos un año de estudio con registros mensuales que permitan registrar la variación estacional que tienen algunas especies. Por último, se destaca la importancia del trabajo conjunto entre las empresas relacionadas a recursos energéticos, las comunidades nativas y el gobierno peruano, para poder desarrollar investigación sobre mamíferos en ambientes remotos donde, de otra manera, sería difícil poder acceder a este tipo de información.

