











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
The genus Cheilanthes Sw. is currently accepted with over 65 species, of which, 26 are South American including its type species C. micropteris Sw. (Ponce & Scataglini 2022). This genus, like other members of the Cheilanthoideae, includes small to medium size terrestrial and saxicolous plants, with pinnate-pinnatifid leaves, and laminae margins mostly revolute and forming a false indusium, homoplastic characters that have hindered its circumscription (Gastony & Rollo 1995).
Cheilanthes is a complex genus, whose circumscription is still uncertain. Both, Ponce and Scataglini (2022), and Sosa et al. (2021) had underlined that the delimitation of the genus Cheilanthes is still in progress, while its affinities within the cheilanthoid ferns are supported as a member of the hemionitid clade (Windham et al. 2009). Progress in molecular studies have also shown that the delimitation in the treatment of Peru is untenable, in part given by the segregation of several species to Aleuritopteris, Astrolepis, Doryopteris, Gaga, and Myriopteris. Furthermore, species still in Cheilanthes include several lineages that could be treated as distinct genera (Kessler et al. 2017, Ponce & Scataglini 2018); however, as stated by Ponce (2016), there are still difficulties in establishing morphological synapomorphies for distinguishing those genera.
The genus Cheilanthes has benefited from several cytological, morphological and molecular phylogenetic studies, like those of Benham and Windham (1992), Gastony (1995), Li et al. (2012), Grusz et al. (2013), Ponce and Scataglini (2018, 2022) and Sosa et al. (2021) allowing for a better definition of its limits and therefore the emerging of a new genus concept. Several genera have been segregated from Cheilanthes, which have also implications for composition and richness of the Peruvian fern flora.
In Peru, Cheilanthes is mainly found in semiarid environments of the Andean slopes, and interandean valleys. Tryon and Stolze (1989) recognized 23 species and mentioned the contentious limits of the genus. Their taxonomic approach was based on morphological characters, many of which are convergent and are present in all lineages of the cheilanthoids (for example genera Myriopteris in Grusz & Windham 2013; and Baja in George et al. 2019).
This study aims to provide an update of the taxonomic treatment of Cheilanthes from Peru. Cheilanthes is presented under the new concept of Ponce and Scataglini (2022), and we complement species observations for several morphological characters not previously discussed. Here we also assessed the conservation status of all four endemic Cheilanthes, following the International Union for Conservation of Nature (IUCN) standards, and we provide a brief assessment for those commonly locally traded. This work also includes a taxonomic key both in Spanish and English for the 15 species currently in Cheilanthes for Peru.
Material and methods
A total of 459 specimens were examined from four Peruvian herbaria (HSP, HUSA, HUT, and USM, citations as in Index Herbariorum). Field work in Apurímac, Ayacucho, Huancavelica, Junín, and Lima allowed complete observations on habitat preferences and altitudinal distribution (2019-2021) (Fig. 1). Additional data was obtained from Tryon and Stolze (1989), photographs of type material from international and national herbaria institutions (B, HUT, P), and from digital collections of the Pteridophyte Collections Consortium (https://pteridophytes.berkeley.edu/ accessed until August 2022). The descriptions include all material examined for this study. For plant collecting, we followed the traditional methods, as described by Cerrate (1969). All collections used for evaluating species conservation status were georeferenced.
Evaluation of morphological characters. Morphological characters were examined in 131 herbarium specimens, from which one to three samples were taken. Examination of indument required a previous rehydration step.
A total of 46 morphological characters were recorded: six quantitative and forty qualitative. Some of the latter have more than one state, such as rhizome type, varying from erect or short to long creeping or multicipital. Characters were defined based on Lellinger (2002) and complemented with Font Quer et al. (1982). Microcharacters were observed using a Leica DM500 optic microscope and a Leica S9i or Nikon SMZ745T stereoscope.
Descriptions include characterization of scales, ratio between frond and petiole, and spore number.
For rhizome and frond scales, measurements’ range and average were taken, considering maximum length and width of one to ten scales, per organ, per specimen. Thirteen species were evaluated for these measurements in the rhizome; we excluded two species (C. andina and C. cantangensis) due to scarcity of material. In the case of frond scales, we were able to register measurements for all six scaly species.
The ratio between frond and petiole lengths were obtained from specimens representing fourteen species, except for C. andina, making one to three measurements, per frond, per specimen.
The number of spores per sporangium was obtained from the examination of 454 sporangia, belonging to 11 species of Cheilanthes. Additional information for C. pruinata, C. rufopunctata and C. poeppigiana was obtained from Ponce and Scataglini (2018). All samples were prepared following D’Ambrosio de Argüeso (1986), with samples being embedded in glycerin gel medium, followed by a resting period of 24 hours at room temperature. A single sporangium was mounted and pressed using the crushing technique. Number of spores was recorded under the microscope, by performing repetitive counting across the whole area of the 22×22 mm cover slip.
Classification and taxonomic identification. The taxonomic position of Cheilanthes in Pteridaceae follows PPG-I (2016), while the genus concept follows Ponce and Scataglini (2022). Protologues were obtained from original sources available in BHL. Also, taxonomic treatments, like those of Ponce (2016) for Argentina, and Kessler et al. (2017) for Bolivia, were used to assess species characteristics and distribution.
A taxonomic key is provided with novel characters for Peruvian species. For each species, nomenclatural history, description, distribution, reviewed specimens, are indicated for the first collector and the herbarium with the acronym in parentheses, additionally accompanied by an illustration. A composed figure for each species was mostly made using Adobe Photoshop CS26, version 13.0.1.
We also reevaluated the proposal of León et al. (2006) for the conservation status of endemic species of Cheilanthes. For conservation status, we considered the criterion of geographic range (B) and used the Geospatial Conservation Assessment Tool (GeoCAT; Royal Botanic Gardens, Kew: http://geocat.kew.org) to measure extent of occurrence (EOO) and area of occupancy (AOO) to apply IUCN categories (IUCN version 3.1, 2012). We include a detailed description of the assessment requirements (Appendix 1).
Results and discussion
Taxonomic treatment of genus Cheilanthes Sw. in Peru
Cheilanthes Sw., Syn. Fil. 5:126. 1806, nom. cons. Lectotype: Cheilanthes micropteris Sw. (designated by Maxon, Sci. Surv. Porto Rico and Virgin Islands 6(3): 428. 1926).
RHIZOMES: erect, decumbent or short to long-creeping, multicipital (many headed; a cluster of short, ascending rhizome branches), scaly. Scales basifixed, bicolorous (partially or wholly sclerotized central area), concolorous, linear-acicular, sinuate, lanceate-triangular, margins continuous, toothed, uncinate, ciliate, the latter inconspicuous. PETIOLES: one vascular bundle near the base, terete, striate, or laevigate, sometimes adaxially sulcate, glabrous, scaly or pilose. Scales sub-basifixed or basifixed, variable in shape, bicolorous or pseudoclathrate (defined cell wall, but not contrasting as clathrate). Hairs simple or branched, eglandular and glandular. LAMINAE: unipinnate to tripinnate, linear-lanceolate to triangular ovate, glabrous or with indument (scales, microscales, hairs, squamules). Scales have similar characteristics than ones on the petioles. Microscales on the adaxial surface of the pinnae in some species. Hairs simple or branched, eglandular, and glandular. Rachises terete, striate, sometimes sulcate and winged adaxially. Pinnae petiolate or adnate, mostly alternate, opposite or subopposite (distanced 1mm). Segment adnate, petiolulate, variable in shape, margins of segment revolute. Veinlets widened at apex, visible or hidden. Pseudoindusium differentiated (lunate, lunulate, or linear) or sometimes hyaline border (not distinct). Sori discrete, sporangia with 16, 32 and/or 64 spores.
Peruvian Cheilanthes are recognized in this updated taxonomic treatment considering the phylogeny of Ponce and Scataglini (2022) as a framework for species affinities. We reexamined the morphological characters historically used for Cheilanthes in Peru (Tryon 1964, and Tryon & Stolze 1989) that allows the acceptance of 15 species for Peru’s flora, one of which was previously considered in synonymy by Tryon and Stolze (1989). However, further sampling and phylogenetic and chromosomal evaluations are still necessary to define and understand limits in several species, especially those not included in previous molecular assessments. Therefore, for this update we provide as many contrasting frond characters as possible between morphologically similar taxa.
The Peruvian species occupy mainly arid and semi-arid areas of interandean valleys and Andean slopes, extending down to coastal hills. They are found in 20 of the 24 departments of Peru, with two distinct regional centres, North and Central-South (Rivera 2022). These species frequently inhabit rocky interstices in open sites, more rarely in Andean forests, between (150-) 500-4200 (-5000) m of altitude. They occupy several ecosystems: grasslands, relict forests, shrublands, seasonally dry forests and lomas (MINAM 2019).
Taxonomic key
[For the use of this taxonomic key, we suggest reviewing illustrations and images provided in this work for each species]
1. Fronds with dense indument, scales on the lamina and axes, or if with hairs, these are scattered to dense on the adaxial surface of the lamina. 2
1’. Fronds glabrous, glabrescent or with indument of hairs on the lamina and axes, sometimes scattered scales on axes. 7
2. Rhizomes with lanceate scales; lamina unipinnate-pinnatisect; adaxial surface with eglandular, multicellular hairs. C. cantangensis
2’. Rhizomes with linear-acicular scales; lamina bipinnate-tripinnate; adaxial surface without visible hairs. 3
3. Scales in the abaxial lamina with reduced cell lumen, margins laciniate accompanied with long filaments extending to the adaxial surface, arachnoid-like, ca. 5 cells in the central area (called squamules). C. scariosa
3’. Scales in the abaxial lamina with translucent visible cell lumen, margins erose, serrate, toothed or laciniate, long filaments absent. 4
4. Laminae bippinate-pinnatisect or bipinnate-pinnatifid; ultimate segments oblong-triangular, ovate-triangular. 5
4’. Laminae bi-tripinnate in the basal pinnae; ultimate segments ovate-triangular, flabellate, and orbiculate. C. incarum
5. Petioles length mostly ¼, ⅕ of the laminae length; basal pinnae gradually reduced or less than ⅔ length of central pinnae. C. lonchophylla
5’. Petioles length as long as the lamina, or near twice the length, or ½, or ⅔ of the lamina length; basal pinnae not reduced. 6
6. Adaxial lamina surface glabrous or occasionally with inconspicuous scales; fertile segment margins not distinctive, hyaline pseudoindusium. C. arequipensis
6’. Adaxial lamina surface mostly with clathrate microscales; fertile segment margins distinctive, membranaceous pseudoindusium. C. peruviana
7. Laminae hairs branched or acaroid-like, dense to sparse. 8
7’. Laminae hairs simple, dense, or absent. 9
8. Rhizome scales lanceate; laminae bi-tripinnate; basal pinnae ⅔ in length of longest distal pinnae; hairs radial to pseudoactinomorphic. C. mollis
8’. Rhizome scales linear-acicular; laminae bipinnate, rarely bipinnate-pinnatifid at the base; basal pinnae as long as the longest distal pinnae; hairs acaroid-like. C. obducta
9. Laminae and axes with eglandular hairs. 10
9’. Laminae and axes glabrous or with glandular hairs. 11
10. Laminae triangular, ovate; rhizome short, erect; castaneous, stramineous scales; rachis sulcate and winged; pseudoindusium with hairs; apex of veinlets evident; fronds with short hairs (2-8 cells), sparce to dense. C. fractifera
10’. Laminae narrowly lanceolate, linear; rhizome creeping; bicolorous (base and central zone blackish brown and castaneous margins) scales; rachis not sulcate nor winged; pseudoindusium glabrous; apex of veinlets not visible; fronds with very long, fine, lanate hairs. C. fraseri
11. Petioles and rachises adaxially sulcate, and with green wings at the rachises; laminae and axes glabrous (except on abaxial side of laminae in C. rufopunctata); apex of veinlets conspicuous, widened. 12
11’. Petioles and rachises adaxially not sulcate; laminae and axes with glandular, multicellular hairs; apex of veinlets not conspicuous. 13
12. Rhizomes erect or decumbent; laminae unipinnate-pinnatisect, without glands on the abaxial surface of laminae. C. poeppigiana
12’. Rhizomes creeping; laminae bipinnate-tripinnate, with reddish glands on the abaxial surface of the laminae. C. rufopunctata
13. Laminae subcoriaceous; fronds glutinous, glandular hairs with dark yellow secretion. C. pruinata
13’. Laminae herbaceous to chartaceous; fronds glutinous without dark yellow secretion. 14
14. Rhizome scales bicolorous (base and central zone blackish brown and atrocastaneous margins), sclerotic; fronds with conspicuous hairs ca. 1mm. C. pilosa
14’. Rhizome scales, concolorous, castaneous, rarely sclerotic; fronds with inconspicuous hairs. C. andina
Clave taxonómica
[Nota: Para el uso de la clave taxonómica, se sugiere consultar las ilustraciones e imágenes de las especies mostradas en este trabajo]
1. Frondas escamosas, escamas densas en lámina y ejes, si con pelos, estos dispersos a densos en el lado adaxial de la lámina. 2
1’. Frondas glabras, glabrescentes o con indumento de pelos en láminas y ejes, a veces escamas esparcidas en los ejes. 7
2. Rizomas con escamas lanceadas; lámina unipinnado-pinnatisecta; superficie adaxial con pelos pluricelulares, eglandulares. C. cantangensis
2’. Rizomas con escamas linear-aciculares; lámina bipinnado-tripinnada; superficie adaxial sin pelos visibles. 3
3. Escamas en la lámina abaxial con lumen celular reducido, márgenes laciniados (acompañado de filamentos largos que se extienden hasta la superficie adaxial, de tipo aracnoides, ca. 5 células en la zona central, llamadas escuámulas). C. scariosa
3’. Escamas en la lámina abaxial con lumen celular visible translúcido, márgenes erosos, aserrados, dentados o laciniados, filamentos largos ausentes. 4
4. Láminas bipinnadas-pinnatisectas o bipinnadas-pinnatífidas; últimos segmentos oblongo-triangular, ovado-triangular. 5
4’. Láminas bi-tripinnadas en las pinnas basales; últimos segmentos ovado-triangulares, flabelados, orbiculares. C. incarum
5. Longitud de peciolos principalmente ¼, ⅕ del largo de las láminas; pinnas basales gradualmente reducidas o menos de ⅔ del largo de las pinnas centrales. C. lonchophylla
5’. Longitud de pecíolos tan larga como las láminas, o casi el doble de la longitud, o ½ o ⅔ de la longitud de las láminas; pinnas basales no reducidas. 6
6. Superficie adaxial de la lámina glabra u ocasionalmente con escamas inconspicuas; márgenes de los segmentos fértiles no distintivos, seudoindusio hialino C. arequipensis
6’. Superficie adaxial de la lámina con microescamas clatradas; márgenes de los segmentos fértiles distintivos, seudoindusio membranáceo C. peruviana
7. Láminas con pelos ramificados, densos a dispersos 8
7’. Láminas con pelos simples, densos o ausentes 9
8. Rizomas con escamas lanceadas; láminas bi-tripinnada; pinnas basales ⅔ en longitud de las pinnas distales más largas; pelos radiales a seudoactinomorfos C. mollis
8’. Rizomas con escamas linear-aciculares; láminas bipinnadas, raramente bipinnada-pinnatifidas en la base; pinnas basales tan largas como las pinnas distales más largas; pelos de apariencia acaroideos. C. obducta
9. Láminas y ejes con pelos eglandulares. 10
9’. Láminas y ejes glabros o con pelos glandulares. 11
10. Láminas triangulares a ovadas; rizomas cortos, erectos; escamas castañas, estramíneas; raquis sulcado y alado; seudoindusio con pelos; ápice de venas visibles y dilatados; frondas con pelos cortos (2-8 células), dispersas a densas. C. fractifera
10’. Láminas angostamente lanceoladas a lineares; rizomas rastreros, escamas bicoloras (base y zona central marrón negruzco y márgenes castaños); raquis no sulcado ni alado; seudoindusio glabro; ápice de venas no visible; frondas con pelos muy largos, finos, lanosos. C. fraseri
11. Peciolos y raquises adaxialmente sulcados, y con alas verdes en el raquis; lámina y ejes glabros (excepto el lado abaxial de la lámina de C. rufopunctata); ápice de venas visibles, ensanchadas. 12
11’. Peciolos y raquises adaxialmente no sulcados; lámina y ejes con pelos glandulares; ápice de venas no conspicuas. 13
12. Rizomas erecto o decumbente; lámina unipinnada-pinnatisecta, sin glándulas en la superficie abaxial de la lámina. C. poeppigiana
12’. Rizomas rastrero; lámina bipinnada-tripinnada, con glándulas rojizas en la superficie abaxial de lámina. C. rufopunctata
13. Láminas subcoriáceas; frondas glutinosas, pelos glandulares con secreción amarillo oscuro. C. pruinata
13’. Láminas herbáceas a cartáceas, frondas glutinosas sin secreción amarillo oscuro. 14
14. Escamas del rizoma bicoloras (base y zona central marrón y márgenes atrocastaneous), esclerosadas; frondas con pelos conspicuos ca. 1mm. C. pilosa
14’. Escamas del rizoma concoloras, castaño claras, raras veces esclerosadas; frondas con pelos inconspicuos. C. andina
Description of Peruvian species
Cheilanthes andina Hook.
Sp. Fil. 2: 115. 1852. Type: Peru, Andes, J. MacLean s.n. (holotype K - K000633278).
Cheilanthes valdiviana Phil., Linnaea 29: 106. 1858. Type: Chile. Valdivia, Huanehue, 1835, C. Gay 2450 (holotype: SGO-82317, photographs BM, SI) syn. nov. designated by M. Ponce and A. Scataglini 2022.
FRONDS: 3.8-10 cm long, caespitose. RHIZOMES: decumbent to short-creeping, scaly. Scales, mostly castaneous, rarely sclerotic. PETIOLES: of length as long as the lamina, sulcate on adaxial side, sometimes with lateral ribs, scales and hairs dispersed in the axes. LAMINAE: herbaceous to chartaceous, deltoid-oblong or narrow lanceate, bipinnate-pinnatifid or tripinnate at the base, inconspicuous hairs similar to the petiole. Rachis with short lateral ribs, with scattered scales and few glandular hairs. Pinnae deltate-ovate to oblong, basal pinnae as long as the central pinnae, with 4-5 sessile lobate pinnulae 0.5-1 mm apart, lobes roundish, with slightly crenulate or entire and revolute margins. Pseudoindusium distinct, scarious, fimbriate-ciliate margins.
Observations. This species was described by John MacLean from a specimen collected probably east of Lima, since other plant collections were made by him in Cajamarquilla and Langa (cfr. Trujillo and Paredes-Burneo 2021), and for Peru, it was the only collection available to us. Tryon (1964) and Tryon and Stolze (1989) considered it a synonym of Cheilanthes pilosa Goldm., observing narrower ultimate segment and thicker texture, explained as a probable response to drier environmental conditions.
Ponce and Scataglini (2022) and Sosa et al. (2021) accepted C. andina as a distinct species from C. pilosa, based on few morphological characters. Cheilanthes andina is characterized by its small frond size, adaxially sulcate petiole with dispersed scales, and with scarce indument in the adaxial surface of the lamina (Ponce, pers. comm. 2022) vs. petiole and raquis terete and rachis scales absent in C. pilosa. We found, however, that frond size and petiole characters varied similarly in more “typical” C. pilosa plants. But a combination of basal pinnae size and spread indument in the adaxial side provides easier separation from C. pilosa. However, further botanical exploration and studies will be required to confirm its distribution range in Peru.
Cheilanthes andina was reported to have 64 spores per sporangium (Sosa et al. 2021). It was proposed as belonging to the Australasian-South American clade based on morphological features (Ponce & Scataglini 2022), and therefore, its affinities are not close to C. pilosa (in the South American clade) as its morphology implies.
Distribution and habitat. Cheilanthes andina, originally described from Peru, is only known from the department of Lima, and it is also reported from Bolivia (Sosa et al. 2021), and from Chile (Ponce & Scataglini 2022) based on synonymizing C. valdiviana Phil., whose type was collected in Valdivia, southern Chile. Therefore, suggesting a wider distribution in Peru, as described here. Ponce and Scataglini (2022) suggested probable affinities with C. sarmientoi Ponce from Argentina, the latter is a small fern with subcarnose laminae and strongly round lobes.
Cheilanthes arequipensis (Maxon) R.M. Tryon & A.F. Tryon
Rhodora 83: 133. 1981.
Notholaena arequipensis Maxon, Smithsonian Misc. Coll. 65(8): 9. 1915. Type: Peru, Arequipa, near Tingo, 5 August 1914, Ms. Rose and J. N. Rose 18797 (holotype US-US761435; isotype NY-NY00144380).
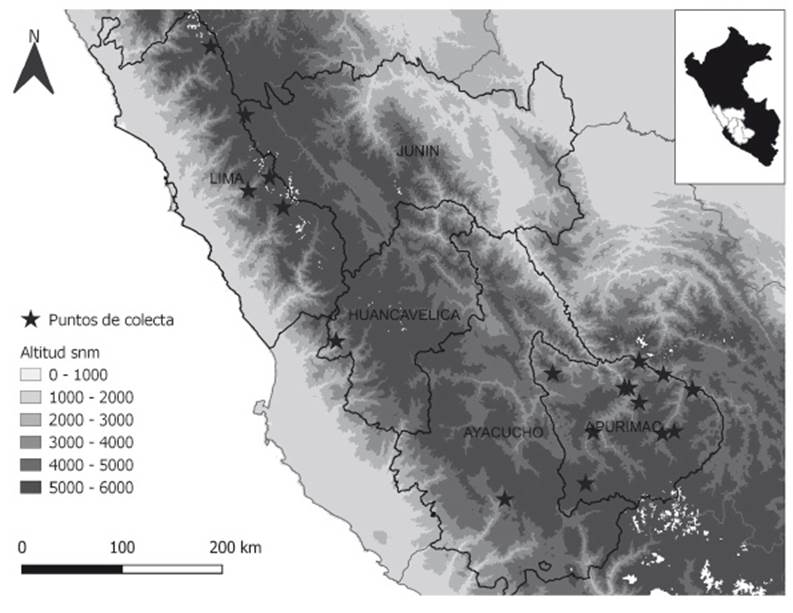
FRONDS: 5.6-17 cm (x 10.55 cm) long. RHIZOMES: erect, short, scaly. Scales 2.4-9 mm (×5.08 mm) × 0.15-1 mm (×0.43 mm), basifixed, brown to ferrugineous, linear-acicular, straight, or slightly sinuate, margins shortly toothed, inconspicuously uncinate. PETIOLES: 2.3-9 cm (×5.71 cm), of length as long as the lamina, or near twice the length of the lamina, terete, striate, dark brown. Scales sub-basifixed, scattered, pseudoclathrate around insertion, lanceate, oblong, round, ovate-triangular, margins toothed, erose to laciniate. LAMINAE: 2.5-8 cm (x 4.81 cm) long, lanceate, lanceolate, rhomboidal, lanceate-triangular, bipinnate to bipinnate-pinnatisect, scaly. Rachises striate. Pinnae opposite or subopposite; proximal pinnae petiolate, distal pinnae adnate, pinnae adaxially glabrous or with indument of scattered scales. Ultimate segment oblong to ovate-triangular. Scales 1.2-4.5 mm (x 2.69 mm) × 0.5-1.5 mm (x 0.98 mm), apices curving towards the adaxial side, imbricate, sub-basifixed, light castaneous, pseudoclathrate at the base, narrowly lanceate, narrowly ovate, elliptic, triangular, slightly subulate, margins laciniate to erose with acute marginal not prolonged tooth. Segment margins revolute, fertile margins not differentiated or sometimes hyaline. Sori discrete, sporangia with 16, 32 y/or 64 spores (Fig. 2).
Observations. Cheilanthes arequipensis is a scaly taxon with close affinities to C. incarum Maxon, C. lonchophylla (Weath. ex R.M. Tryon) R.M. Tryon & A.F. Tryon, C. peruviana (Desv.) T. Moore and C. scariosa (Sw.) C. Presl, all members or putative members of the South American-Australasian clade (Ponce & Scataglini 2022). From C. peruviana, it can be distinguished by its laminar scales with toothed acute margins and not distinctive pseudoindusium as shown in the key. It differs from C. incarum by having oblong segment and adaxially glabrous or with indument of scattered scales (vs. orbicular segment and linear or acicular, dispersed microscales).
Cheilanthes arequipensis and allied Peruvian species were proposed as closely related to the non-Peruvian species C. squamosa Gillies & Hook. ex Grev. (Ponce & Scataglini 2022). These two species can be morphologically distinguished by having rhizome scales linear-acicular, straight, or slightly sinuate, margins shortly toothed in C. arequipensis vs. triangular scales, conspicuous, retrorse toothed margin in C. squamosa. Cheilanthes arequipensis has a partial sympatric distribution with C. squamosa in Chile and Argentina.
Distribution and habitat. Cheilanthes arequipensis is mostly known from southern Peru, extending its range around and above 3000 m a.s.l. to northern Chile and northwestern Argentina A. In Peru, this species has been recorded in Arequipa, Lima, Moquegua and Tacna from 2400 to 3480 m a.s.l. It grows in rocky slopes, and humid ravines within shrublands.
Examined specimens. Arequipa: Arenas G.102 (USM); Cáceres F. 5333, 1605, 8518 (HUSA); Cueva M. 31 (HUSA); León B. 5737b (USM); Munz P. 15500 (GH); Pennell F. 13198 (B. F. GH, US); Quipuscoa V. 1744, 2512, 2526 (HUSA); Roque J. 906 (USM); Stafford D. 570 (BM); Vargas 8019 (UC); Williams R. 2638 (GH, NY, US). Lima: Beltrán H. 2952 (USM); Saunders 612 (F). Moquegua: Montesinos T. 2097 (WAG). Tacna: La Torre M. 1844 (USM).
Cheilanthes cantangensis (R.M. Tryon) R.M. Tryon
Fieldiana Bot., n.s. 22 (2): 32. 1989.
Notholaena cantangensis R.M. Tryon, Rhodora 63: 81. 1961. Type: Peru, Dep. Cajamarca, Prov. Celendin, Cantange, Ruta Celendin-Rio Marañon, 4 June 1960, A. López and A. Sagástegui 3366 (holotype GH; isotype 3366-HUT).
FRONDS: 13-44.2 cm (x 30.63 cm) long. RHIZOMES: short, erect. Scales bicolorous (sclerotic central portion and brown), dark castaneous, basifixed, lanceate, margins dentate to serrulate. PETIOLES: 4.8-20 cm (x 12.93 cm), near of length as long as the lamina, or ½, ⅔ of the lamina length, terete, striate, scabrous or muricate, brown. Scales sub-basifixed or basifixed, non-clathrate or pseudoclathrate, lanceate to triangular or ovate, margins serrate, dentate, ciliate to laciniate. LAMINAE: 8.2-24.2 cm (x 17.70 cm) long, lanceate or triangular to elliptic, pinnate-pinnatisect to bipinnate-pinnatifid. Scales 1.2-2.7 mm (x 2.17 mm) × 0.2-0.9 mm (x 0.52 mm), light castaneous and dark. The axes are densely scaly. Pinnae oblong, lanceate, petiolulate, proximal and central pinnae subopposite (distanced 1mm) and slightly asymmetrical. Segment adnate, adaxially pubescent, dense or spread, and abaxially densely scaly. Scales imbricate, non-clathrate or pseudoclathrate, lanceate or slightly subulate, margins laciniate, apices caudate. Hairs non-glandular, multicellular with 12-15 rectangular cells, distal cell smaller or shorter than the rest. Segment margins revolute, fertile margins not differentiated or sometimes hyaline. Sori discrete, 64 spores per sporangium (Fig. 3).

Observations. This species is easily recognized by the combination of non-glandular, long hairs on the adaxial surface and densely imbricate scales abaxially. Tryon (1956) proposed this taxon under Notholaena, commenting on its affinities to Paraguayan N. hassleri Weath. (later recognized as Cheilanthes hassleri (Weath.) Ponce), because of the presence of similarly sparse hairs in laminae. These species are morphologically very similar, differ because in C. hassleri the pinnae are more developed basiscopically, are elongated in the proximal pinnatifid (vs. proximal and central pinnae subopposite and slightly asymmetrical in C. cantangensis, although this character needs further verification with fresh material). Affinities of C. cantangensis are currently more clearly understood. Ponce and Scataglini (2018) included C. hassleri but not C. cantangensis in their two-chloroplast marker phylogeny, resolving it as a member of the “C. obducta group”, a group sister to Hemionitis. This lineage was later not supported apart from Cheilanthes by Ponce and Scataglini (2022), who also suggested C. cantangensis and C. hassleri to belong to the South American-Australasian clade, due to their morphological similarities to species in this clade, like C. obducta and the Brazilian C. pantanalensis E.L.M. Assis, Ponce & Labiak.
Distribution and habitat. Cheilanthes cantangensis is a Peruvian endemism, only known from northern Peru (Amazonas and Cajamarca). The original locality remains the single record for its presence in Cajamarca, while other collections have confirmed its range extension to the north along the Marañon and the Utcubamba basin. It has been recorded from 500-1900 m a.s.l., inhabiting seasonally dry forest ecosystems in areas dominated by shrubs.
Examined specimens. Amazonas: Gonzáles P. 6067 (USM); van der Werff H.14607, 14670 (USM). Cajamarca: López A. 3366 (HUT).
Cheilanthes fractifera R.M. Tryon
Rhodora 62: 7, t. 1252. 1960. Type: Peru, Dep. Ayacucho, 45 km from Nasca to Puquio, 14 February 1958, D.S. Correll and E.E. Smith P169 (holotype GH-GH00020739; isotype TEX-LL0037001).
Cheilanthes saundersii Alston, Lilloa 30: 110, t. 6. 1960. Type: Peru, Dep. Lima, Huarochiri, km 64 Central Highway, 28 April 1957, S.G.E. Saunders 353 cited as “350” (holotype BM-BM00936608; isotype US-US236049).
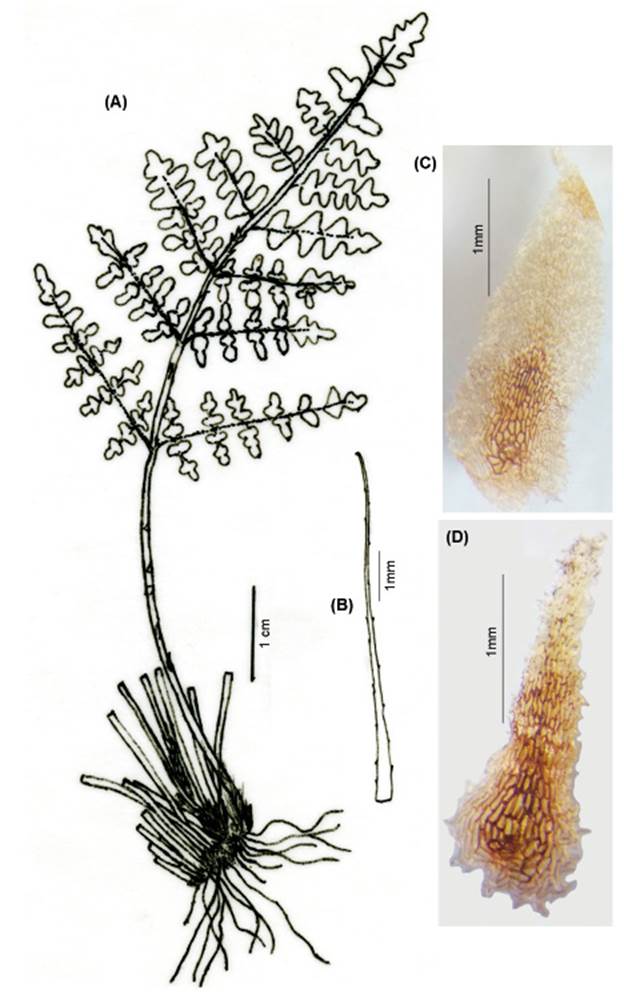
FRONDS: 7.5-15 cm (x 11 cm) long. RHIZOMES: short, erect. Scales 1.5-8.0 mm (x 4.13 mm) × 0.2-1.2 mm (x 0.59 mm), basifixed, sub-basifixed, concolorous, castaneous, stramineous, linear, or acicular, lanceate or triangular, margins entire or toothed, uncinate, ciliate. PETIOLES: 2.0-5.0 cm (x 4.0 cm), of length as long as the lamina, or ½, ¼, ⅔ of the lamina length, terete, laevigate, or striate, brown to dark, ebeneous, pubescent, hairs like those of the laminae, petiole base fractiferous with conspicuous persistent scales, the latter lanceate to triangular, caudate apices. LAMINAE: 3.5-10 cm (x 7.0 cm) long, triangular, ovate, unipinnate-pinnatisect to bipinnate-pinnatifid, pubescent. Rachises sulcate and wings adaxially, ebeneous color limited to the first pinnae pair, distal portions green. Pinnae, ovate to triangular, sessile to shortly petiolulate 1-2 mm long, proximal pair opposite to subopposite, and distal ones alternate, both sides with eglandular hairs, the latter simple, 2-8 cells, spreading. Segment margins revolute. Veinlets with broad apices and visible. Pseudoindusium membranaceous, lunulate, margin crenate, lobed, erose, and pubescent. Sori discrete, sporangia with 32 spores (Fig. 4).
Observations. Cheilanthes fractifera is easily recognizable from other Peruvian Cheilanthes by the combination of its characters: ebeneous, fractiferous petioles with dispersed whitish hairs and spread scales, and pubescent laminae. The presence of 32 small spores supports its definition as having a sexual reproduction system.
Its morphology resembles that of the western North American C. brandegeei D.C. Eaton, currently recognized as Baja brandegeei (D.C. Eaton) Windham & L.O. George, that made Tryon and Tryon (1982) to propose their close affinity. However, laminae outline shape, fractiferous petiole, and pseudoindusium are homoplasious characters; molecular data supported the recognition of B. brandegeei apart from Cheilanthes and any of the hemionitid ferns (George et al. 2019).
Morphologic differences exist in each life stage of B. brandegeei and C. fractifera. The sporophyte of B. brandegeei has unicellular, acicular hairs from a dilate, multicellular base; while the sporophyte of C. fractifera has hairs made of single row cells. The gametophyte growth in B. brandegeei is typically bommeriid, while in C. fractifera is typically hemionitid (George et al. 2019).
Distribution and habitat. Cheilanthes fractifera is a Peruvian endemism. It is known from Arequipa, Ayacucho, La Libertad, and Lima. It grows in rocky slopes and in dry ravines among interstices, and with an altitudinal range of 1450-2900 m a.s.l.
Examined specimens. Arequipa: Cáceres F. 1629 (HUSA); Cano A. 6419 (USM); *Muller G. 12592 (GH); Montesinos D. 3351 (HUSA, USM), 3347 (USM); Obando Y. 21 (USM); *Rauh W. P570 (B); Weigend M. 9864/ MW 19-54 (HUSA, USM). La Libertad: Sagástegui A. 4482 (HUT). Lima: León B. 5712 (USM); Rauh W. P143 (B); Saunders S. 219 (BM), 1050 (GH). *Records not used for conservation status assessment.
Cheilanthes fraseri Mett. ex Kuhn
Linnaea 36: 83. 1869. Syntypes: Peru, H. Ruiz and J.A. Pavón s.n. (B-B200101079a). Ecuador, J. Fraser s.n. (B-B200101078); Ecuador, M. Wagner s.n. (Type fragment: B-B200101079b).
Notholaena fraseri (Mett. ex Kuhn) Baker, Syn. Fil. (Hooker and Baker) ed. 2, 514. 1874.
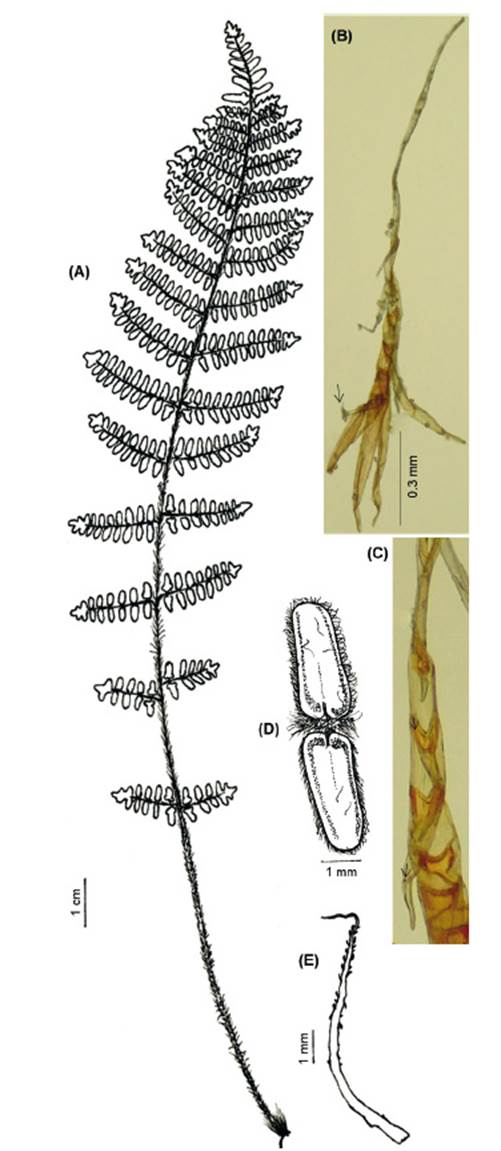
FRONDS: 8.1-33.5(-38.5) cm (x 20.85 cm) long. RHIZOMES: short or long-creeping, entire or branched, 1-10 mm distance between fronds. Scales 1.0-2.6 mm (x 1.83 mm) × 0.1-0.5 mm (x 0.25 mm), basifixed or adpressed, bicolorous (base and central zone blackish brown and castaneous margins), lanceate with caudate translucent apices, margins toothed or slightly uncinate. PETIOLES: 3.6-15.5(-25) cm (x 8.52 cm), of length as long as the lamina, or longer than the length of the lamina, or ½ to ⅓ of the lamina length, terete, brown, villose, or tomentose, hairs similar to those of the laminae, or glabrescent. LAMINAE: 3.9-21.3(-24.5) cm (x 12.30 cm) long, linear or narrowly lanceolate, unipinnate-pinnatifid to bipinnate-pinnatifid at the base, simple glandular hairs, sericeous, villous, tomentose, lanate, or glabrescent. Rachises terete, striate and indument similar to the petiole. Pinnae ovate or triangular, asymmetric having larger basiscopic side, proximal and central pinnae petiolulate, proximal pinnae slightly reduced with 3-5 segments per side, while central and distal pinnae with up to 5 segments per side. Segment margins revolute. Veinlets with visible and broad apices. Pseudoindusium membranaceous, linear or lunate, margins erose. Sporangia with 32 and 64 spores (Fig. 5).

Observations. Cheilanthes fraseri is one of seven species resolved as members of the South American clade (Ponce & Scataglini 2022), all sharing multicellular hairs.
Cheilanthes fraseri was considered morphologically similar to C. bonariensis (Willd.) Proctor (Tryon and Stolze 1989), however, Grusz and Windham (2013) transferred it to Myriopteris, as M. aurea (Willd.) Grusz & Windham, a segregated genus encompassing a monophyletic group earlier recognized as the “myriopterid clade” (Windham et al. 2009), and distantly related to the type species of Cheilanthes. Both, C. fraseri and M. aurea share general lamina morphology and indument, however, they differ by creeping rhizomes and scales with translucent apices in the former vs. nodose rhizomes and scales with sclerosed apices in the latter.
Two other species of Cheilanthes, not found in Peru, are also similar to C. fraseri, these species are C. buchtienii (Rosenst.) R.M. Tryon from Bolivia and Argentina, and C. hypoleuca (Kunze) Mett. from Chile and Argentina (Ponce 2016). Tryon and Stolze (1989) cited C. buchtienii and C. hypoleuca as probably present in the Peruvian flora. Cheilanthes buchtienii can be distinguished of C. fraseri by having narrowly triangular laminae, and few deciduous scales among hairs on the rachises (vs. linear or narrowly lanceolate and no scales); while C. hypoleuca has canaliculate petiole and rachis, and subulate rhizome scales (vs. terete axes and lanceate rhizome scales).
Distribution and habitat. From Ecuador to Bolivia. In Peru is found in Amazonas, Ancash, Apurimac, Ayacucho, Cajamarca, Cusco, Huancavelica, Huanuco, Junin, La Libertad, Lima, and Piura. It grows in rocky slopes of seasonally dry tropical forests and shrublands, between 800-2600 m a.s.l.
Examined specimens. without locality, Raimondi A. s.n. (USM). Amazonas: Werff H. 16878 (USM). Ancash: Cerrate E. 4049, (USM); Weberbauer A. 3020 (USM). Apurimac: Rivera S. 65, 73, 81, 87, 100, 101 (USM); Vargas 8910 (MO, UC); Zúñiga V. 640 (USM). Ayacucho: Velarde 4951 (GH). Cajamarca: Correll D. P909 (GH, US); Gonzáles P. 5388 (USM); Weberbauer A. 4139 (USM). Cusco: Mexia Y. 8055 (USM); Stork 10449 (F, GH, UC); Vargas C. 22365 (USM). Huancavelica: Huamán E. s.n. (USM); Hutchison P. 1684 (USM); Rivera S. 103 (USM); Tovar O. 3848 (USM). Huanuco: Macbride J. 2328 (F, GH, US); Calderón S. s.n. (USM); Ferreyra R. 6912, 19401 (USM); Mantel M. 1873, s.n. (USM). Junin: Weberbauer A. 1991 (USM). La Libertad: Castillo H. 152a; Gonzáles P. 5894, 683, 6847, (USM). Lima: Rauh W. 145, 146 (USM). Piura: Scolnik R.1435 (UC).
Cheilanthes incarum Maxon
Smithsonian Misc. Coll. 65(8): 5. 1915. Type: Peru, Cuzco, 1 September 1914, J.N. Rose and Rose 19061 (holotype US-US761644; isotype NY- NY0014436, fragment P-P00584337).
FRONDS: 12.5-37.5 cm (x 23.82 cm) long. RHIZOMES: multicipital, compact, erect or short- creeping. Scales 5.2-11 mm (x 8.20 mm) × 0.15-0.8 mm (x 0.33 mm), basifixed, castaneous to atrocastaneous, linear to acicular, sinuate, margins toothed, occasionally inconspicuously uncinate. PETIOLES: 3.8-14.2(-14.5) cm (x 7.72 cm) long, of length as long as the lamina, or ⅔, ½, ⅓ to ¼ of the lamina length, brown, terete, striate, scaly. Scales sub-basifixed, pseudoclathrate, lanceate-ovate, lanceate-triangular, or pyriform, apices acuminate, acute, cuneate or caudate, margins erose, serrate, toothed or laciniate LAMINAE: 8.5-23.3 cm (x 16.16 cm) long, narrowly lanceate, lanceolate, bipinnate-pinnatifid basally tripinnate (basal segment basiscopically divided), scaly. Scales 1.3-4.5 mm (x 2.95 mm) × 0.5-1.8 mm (x 1.08 mm). Rachises terete, striate, scales similar to those of the petioles. Pinnae, ovate-triangular, petiolulate, basal pinnae subopposite (distanced 1mm) and near same length as central pinnae. Apices of abaxial scales extending to the adaxial side. Scales similar to petioles, sub-basixifed, imbricate, pseudoclathrate, light castaneous and grayish. Microscales, sinuate, linear or acicular, dispersed on the adaxial side, ca. 1-4 mm long, ca. 4 cells wide, margins uncinate, toothed. Segment ovate a triangular, flabellate, orbicular. Segment margins revolute. Pseudoindusium distinct, membranaceous, scarious, lunulate, margins erose. Sori discrete, sporangia with 32 spores (Fig. 6).

Observations. Cheilanthes incarum is morphologically similar to other four Peruvian species, C. arequipensis, C. peruviana, C. lonchophylla, and C. scariosa, by being densely scaly on the axes and on the abaxial side of the laminae. It differs from C. scariosa by microscales on the adaxial surface of segments (vs. absent). Ponce and Scataglini (2022) suggested affinities of C. incarum to C. squamosa as part of the South American-Australasian clade that also includes those Peruvian species discussed above. Cheilanthes incarum shares similar indument of microscales on the adaxial side of laminae with C. squamosa; and differs by oblong ultimate segment in the latter vs. orbicular segment in the former.
Distribution and habitat. Cheilanthes incarum occurs in Peru and Bolivia, probably in northwest Argentina, although it was not included by Ponce (2016) for its pteridoflora, and it is not listed for Zuloaga (2018). In Peru, it has been recorded in Ancash, Apurimac, Arequipa, Cusco, Huancavelica, Junin, Moquegua and Puno. It grows in open, exposed xerophytic sites, in rocky slopes and dry ravines, between 2200-4200 m a.s.l.
Examined specimens. without locality, Tovar O. 1391 (USM). Ancash: Griffin D. s.n. (USM); Weberbauer A. 3021a (USM). Apurimac: Hutchison 1690 (GH); Lehnert M. 340 (USM); Rivera S. 94 (USM); West 3903 (UC). Arequipa: Montesinos D. 2978 (USM, HUSA). CU: Davis W. 1455 (USM); Ferreyra R. 2668 (USM); Núñez P. 7939, 7332 (USM); Tryon R. 5343 (USM). Huancavelica: Tovar O. 1014 (USM). Junin: Cerrate E. s.n. (USM). Moquegua: Montesinos D. 2244 (USM). Puno: Tutin 1032 (BM).
Cheilanthes lonchophylla (Weath. ex R.M. Tryon) R.M. Tryon & A.F. Tryon
Rhodora 83: 133. 1981.
Notholaena lonchophylla Weath. ex R.M. Tryon, Contr. Gray Herb. 179: 19. 1956. Type: Peru, Chachapoyas, 1939, B.F. *Mathews 610 (holotype K-K000633281; isotype P-P00586992).
FRONDS: 19.4-27 cm (x 22.47 cm) long. RHIZOMES: short, scaly. Scales 6-9.5 mm (x 8.56 mm) × 0.1mm, basifixed, castaneous to atrocastaneous, linear or acicular, sinuate, margins toothed, uncinate. PETIOLES: 3.4-5 cm (x 4.47 cm), petiole length mostly < ¼ length of the lamina or ⅕, striate, terete, brown. Scales dark castaneous in the center with lighter margins, sub-basifixed, pseudoclathrate, lanceate, or elliptic, margins erose to serrate or laciniate, apices caudate. LAMINAE: 16-22 cm (x 19 cm) long, narrowly lanceolate or lanceolate, bipinnate-pinnatifid, basal pinnae gradually reduced or less than ⅔ length of central pinnae, scaly. Scales 2-3.2 mm (x 2.64 mm) × 0.7-1.2 mm (x 0.96 mm). Rachises striate, scales similar to those of the pinnae. Pinnae, ovate to triangular, petiolulate. Segment oblong to triangular. Abaxial scales apices curved to the adaxial side. Abaxial side densely scaly, scales, sub-basifixed, imbricate, dark castaneous in the region central and margins light, lanceate to triangular, margins toothed, laciniate, apices caudate. Adaxial side with spreading microscales or glabrescent, microscales linear or acicular. Segment margins revolute. Pseudoindusium distinct, membranaceous, scarious. Sori discrete, sporangia with 32 spores (Fig. 7).

Observations. Cheilanthes lonchophylla is related to other scaly Peruvian species, C. arequipensis, C. incarum, C. peruviana and C. scariosa, all members of the South American-Australasian clade (Ponce & Scataglini 2022). It is recognized as distinct from the other members mentioned in the taxonomic key. Cheilanthes scariosa differs too from C. lonchophylla by its orbiculate ultimate segment. Ponce and Scataglini (2022) also proposed affinities to C. squamosa as part of the South American-Australasian clade. Cheilanthes lonchophylla shares similar indument of microscales on the adaxial side of laminae with C. squamosa, but they differ by narrowly lanceate laminae in the former vs. ovate lanceolate in the latter.
Distribution and habitat. Cheilanthes lonchophylla is a Peruvian endemic, known from Amazonas, Cajamarca and La Libertad. It is known from 2200-2950 m a.s.l. It inhabits rocky sites associated to seasonally dry tropical forest.
Examined specimens. Cajamarca: *Aguado H. 6791 (HUT); Hutchinson P. 5297 (UC); López A. 7653 (HUT); Mostacero J. 881 (HUT). La Libertad: Gonzáles P. 7193 (USM). *Records not used for conservation status assessment.
Cheilanthes mollis (Kunze) C. Presl
Tent. Pterid. 160. 1836.
Notholaena mollis Kunze, Linnaea 9: 54. 1834. Type: Chile (Valparaíso), near Valparaiso, Playa Ancha, June 1827, E.F. Poeppig s.n. (LZ, destroyed) (lectotype designated by M. Ponce and A. Scataglini 2022 as LE-00008529; and isolectotype: BM-001067980).
FRONDS: 10.5-27 (x 18.17 cm) long. RHIZOMES: short or long-creeping, nodose, scaly. Scales 2.2-3 mm (x 2.58 mm) × < 1mm, lanceate, basifixed, adpressed, bicolorous (base and central zone blackish brown and light castaneous margins), margins inconspicuously toothed. PETIOLES: 3.5-9 (x 6.25 cm), ½ to ⅓ of the lamina length, terete, laevigate, or striate, brown, pubescent to glabrescent, eglandular hairs, branched, stellate or pseudactinomorphs, sparse scales spreading along the axes base similar to those of the rhizome. LAMINAE: 7-18 (x 12 cm) long, lanceolate, bi-or-tripinnate, pubescent. Pinnae petiolulate. Segment orbiculate, or oblong. Abaxial surface densely hairy, tomentose, pluricellular eglandular hairs, stellate or pseudactinomorphs, with (8-) 9-12 long branches around an axis of few articulate cells, whitish or ferrugineous. Adaxial surface glabrescent or with spreading hairs similar to those abaxially. Segment margins revolute, fertile margins not differentiated or sometimes hyaline. Sori discrete on vein apices, sporangia with 32 spores (Fig. 8).

Observations. Cheilanthes mollis is easily distinguished from other Peruvian species by its fronds: laminae tomentose on the abaxial surface with stellate hairs. It was included in the phylogeny of Ponce and Scataglini (2022) and resolved within the South American clade that included another five Peruvian species (C. fraseri, C. pilosa, C. poeppigiana Mett. ex Kuhn, C. pruinata Kaulf., and C. rufopunctata Rosenst.).
Distribution and habitat. It is known from Peru and Chile. In Peru, it has been found in Arequipa, growing in rocky sides and ravines of lomas ecosystem, between 50-1050 m a.s.l. In Chile, it also occupies low elevation sites below 1300 m (Rodríguez et al. 2009).
Examined specimens. Arequipa: Cano Y. 33 (HUSA); Ellenberg H. 4153 (GH); Hutchison P.C. 7130 (USM); Weberbauer A. 1545 (B).
Cheilanthes obducta Mett. ex Kuhn
Linnaea 36: 83. 1869. Type: Bolivia, La Laguna, June 1907, C. D. D'Orbigny 386 (holotype B-B200102214; isotype P-P00586581, P-P00586582).
Notholaena obducta (Mett. ex Kuhn) Baker, Syn. Fil., ed. 2, 515. 1874.
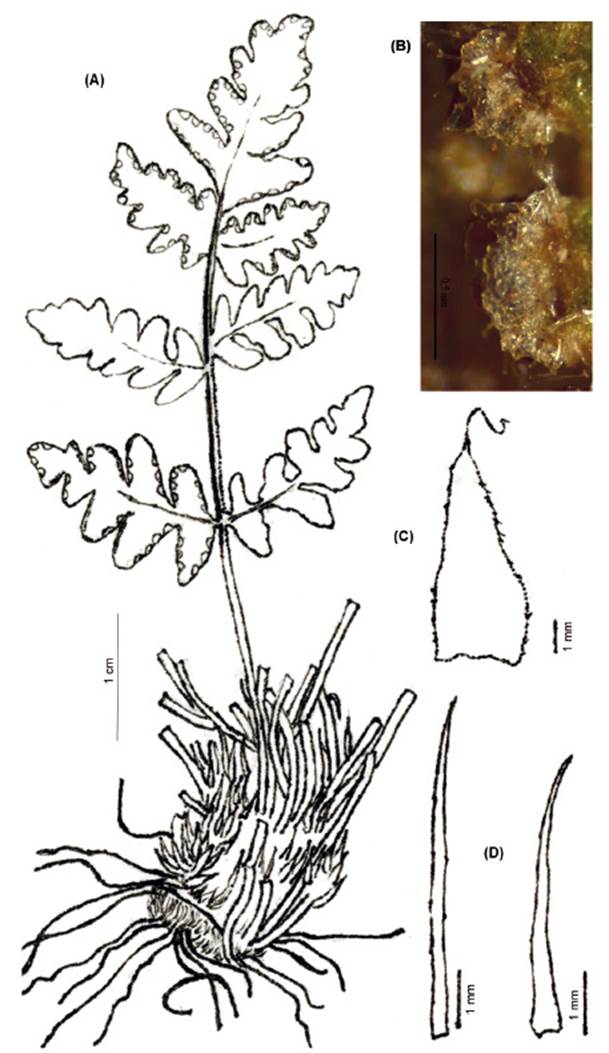
FRONDS: 8.7-33 cm (x 21.39 cm) long. RHIZOMES: short-creeping, compact, scaly. Scales 4-12 mm (x 6.54 mm) × 0.1-0.3 mm (x 0.18 mm), basifixed, castaneous or atrocastaneous, linear or acicular, slightly sinuate, margins apically uncinate or toothed. PETIOLES: 1.5- (2.7-5.5) -7.5 cm (x 4.18 cm) long, near ⅓, ¼, ⅕, ⅙ of the lamina length, brown, terete, laevigate or striate, pubescent, lanate, eglandular, acaroid-like hairs. LAMINAE: 7.2-22.5 (x 16.44 cm) long, lanceolate, lanceate or elliptic, bipinnate, rarely bipinnate-pinnatifid at the base, pubescent with hairs similar to those on petioles, lanate. Segment adnate or shortly petiolulate, oblong-deltoid, margins revolute. Abaxial surface with multicellular, eglandular, sometimes they have a capitate terminal cell, acaroid-like hairs, the latter short multicellular base (1-2 cells), main axes in a single row of caudate or pectinate cells, with tangential walls embedded into the neighboring distal cell. Adaxial surface glabrescent or villous, hairs similar to those on opposite side. Segment margins fertile not differentiated or sometimes hyaline. Sori discrete, sporangia with 32 spores (Fig. 9).
Observations. Cheilanthes obducta is distinguished from other hairy Peruvian species by its lanceate bipinnate laminae with acaroid-like hairs, these hairs have a 1-2 cells base, main axes in a single row of caudate or pectinate cells, with tangential walls embedded into the neighboring distal cell (Fig 9D). Ponce and Scataglini (2018), in their two-chloroplast marker phylogeny, recognized the “C. obducta group”, that also included C. hassleri and the Brazilian C. pantanalensis, as a group sister to Hemionitis. Nevertheless, this lineage was later not supported apart from Cheilanthes by Ponce and Scataglini (2022), but instead was well-resolved in Cheilanthes with the inclusion of Argentinian C. sarmientoi Ponce, and suggesting C. cantangensis and C. hassleri as members of this lineage, all together members of the South American-Australasian clade.
Distribution and habitat. This species occurs from Colombia and Venezuela to Bolivia, Amazonian Brazil, Paraguay, and Argentina. In Peru is known from Amazonas, Cajamarca, and La Libertad, from 600-1700 m a.s.l. It grows between rocks in open sites or forested areas in the seasonally deciduous dry tropical forest ecosystem.
Examined specimens. Amazonas: Gonzáles P. 5618 (USM); Mendoza W. 6614 (USM). Cajamarca: Gonzáles P. 5373, 6214 (USM); Hutchison P.C. 1425 (USM). La Libertad: Arroyo S. 236 (HUT); Gonzáles P. 6559 (USM); Pillaca L. MA18 (USM).
Cheilanthes peruviana (Desv.) T. Moore
Index Fil. 250. 1861.
Notholaena peruviana Desv., Mém. Soc. Linn. Paris 6: 220. 1827. Type: Peru, locality unknown, *J. Dombey s.n. (holotype P-P00586588; isotype B-B200127985; fragment P-P00586540).
Notholaena sinuata var. bipinnata Hook., Sp. Fil. 5: 108. 1864. Type: Peru, (Huanuco), “Andes of Peru”, Baños, Brackenridge (Wilkes, U.S. Exploring Expedition) (holotype K).
Notholaena brackenridgei Baker, Syn. Fil. 371. 1868. Type: the same as for N. sinuata var. bipinnata Hook.
FRONDS: 7.8-29.9(-31.3) cm (x 16.25 cm) long. RHIZOMES: multicipital, compact, short, scaly. Scales 3.5-10.5 mm (x 5.57 mm) × 0.1-1mm (x 0.25 mm), basifixed, concolorous, brown, pale castaneous in group, linear to acicular, sinuate, margins uncinate and inconspicuously toothed. PETIOLES: 3-17.9 cm (x 8.11 cm), of length as long as the lamina, or near twice the length, or ½, ⅔ of the lamina length, terete, brown, castaneous, striate with scars, indument scaly or glabrescent. Scales sub-basifixed, pseudoclathrate, lanceate or triangular, or acicular, elliptic or ovate, apices attenuate or caudate, margins toothed or laciniate, inconspicuously uncinate. LAMINAE: 3-16.8 cm (x 8.15 cm) long, narrowly lanceate to narrowly elliptic, bipinnate-pinnatifid, scaly. Scales 1-2.5 mm (x 1.73 mm) × 0.1-0.9 mm (x 0.62 mm), castaneous light and dark. Rachises striate with indument similar to petioles. Pinnae, ovate, petiolulate, mostly alternate. Abaxial surface densely scaly, similar to those on petioles, pale castaneous or dark, scale apices curved towards the adaxial surface, pseudoclathrate, sub-basifixed, imbricate, marginal apiculate tooth. Adaxial side with microscales clathrate, scattered, light castaneous and dark (ca. 1 mm long, ca. 3 × 5 cells). Segment ovada-triangular, margins revolute. Pseudoindusium membranaceous, lunulate, margins erose. Sori discrete, sporangia with 16, 32 and/or 64 spores (Fig. 10).
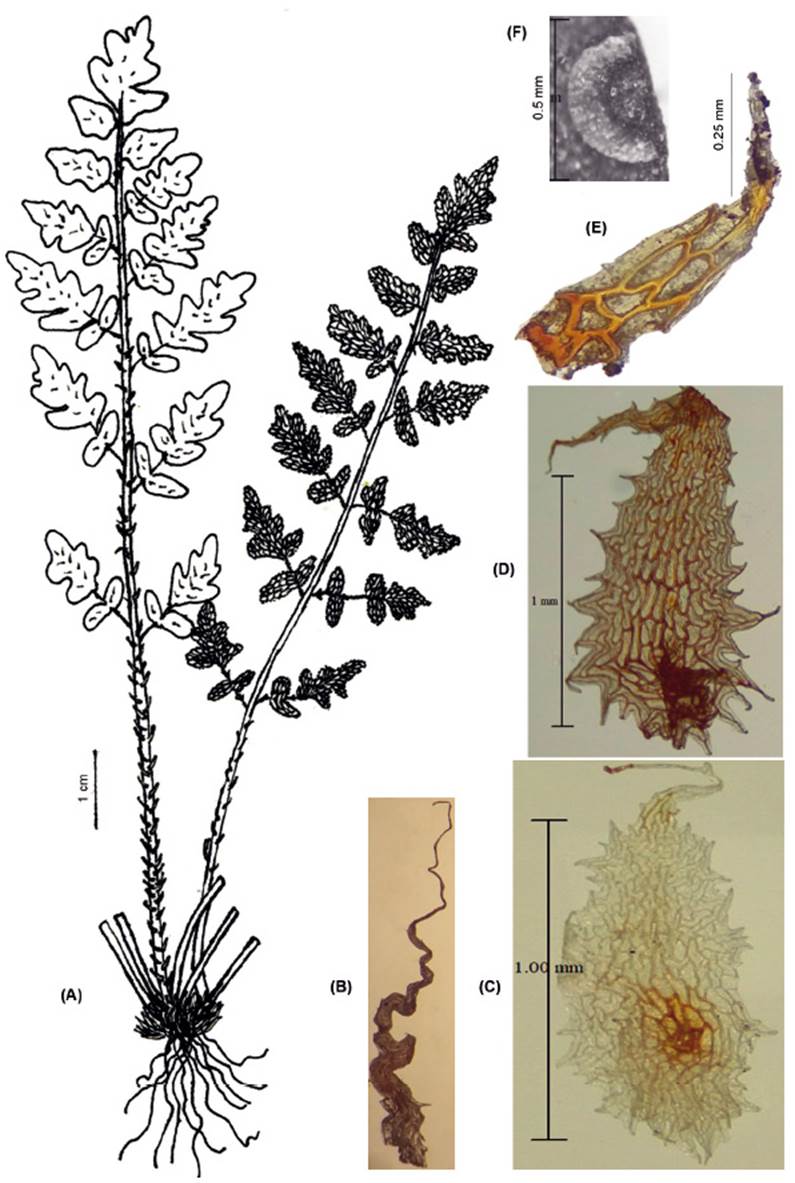
Observations. Cheilanthes peruviana is a scaly species with close affinities to C. arequipensis, C. incarum, C. lonchophylla, and C. scariosa, all members or putative members of the South American-Australasian clade (Ponce & Scataglini 2022). It is confused with C. arequipensis, but it can be distinguished: laminar scales with marginal apiculate tooth, clathrate microscales and pseudoindusium differentiated (vs. laminar scales with marginal acute tooth, without microscales and with pseudoindusium not differentiated in C. peruviana). Tryon and Stolze (1989) distinguished C. peruviana from C. arequipensis based on the color of rhizome scales, and laminae to petiole ratio length; however, these characters are plastic. Cheilanthes peruviana also differs from C. incarum by its adaxial side with clathrate microscales (vs. pseudo-clathrate microscales); and ovate-triangular ultimate segment (vs. mostly orbicular, flabellate or ovate a triangular).
Distribution and habitat. Cheilanthes peruviana is a Peruvian endemic. It is known from Ancash, Arequipa, Cajamarca, La Libertad, Lambayeque, Lima and Moquegua, from 400-3900 m a.s.l. It grows in rocky slopes, in ravines, and shrublands associated to lomas and relict Andean forests ecosystems.
Examined specimens. Ancash: Cerrate E. 6638 (USM); Smith D. 10896 (USM). Arequipa: Cano Y. 48 (HUSA); Coronado P. 39 (US); Del Carpio C. 140 (USM); Ferreyra R. 1426 (USM), 11968 (GH); Montesinos D. 3465 (USM); Quispe E. 124 (HUSA); Whaley O. 1840 (USM). Cajamarca: López A. 3688 (HUT), Sagástegui A. 3892, 5073, 6392, 7723, 8379, 9146, *9271, *10599, 12530, 10631a (HUT); Vargas M. 2220 (HUT). La Libertad: Sagástegui A. 3913 [before as Cajamarca] (HUT); Arroyo S. 242 (HUT); López A. 2665 (HUT); Weigend M. 97/210 (USM). Lambayeque: Correll D.S. P822 (US). Lima: Acleto C. 40 (USM); Cerrate E. 5753, 5775, 6344, 6358, (USM); Ferreyra R. 3458, 9742, 15464 (USM); Gonzáles P. 20 (USM, HUSA), 586b (USM); León B. 904 (USM); *Meza I. 12 (USM). Moquegua: Stafford D. 993 (BM). *Records not used for conservation status assessment.
Cheilanthes pilosa Goldm.
Nov. Act. Acad. Caes. Leopold. Carol. Nat. Cur. 19 (suppl. 1): 455. 1843. Type: Peru, F.J.F. Meyen s.n. (lectotype designated by Ponce 2016: B-20-0038096; isolectotype: B-20-0038097).
Cheilanthes macleanii Hook., Sp. Fil. 2: 93, t. 110 B. 1852. Type: Andes of Peru, J. MacLean s.n. (holotype K- K000633286; isotype GH-GH00571977).
FRONDS: 7-25.2 (-45) cm (x 22.98 cm) long. RHIZOMES: short or long-creeping, scaly. Scales 2.8-4.5 mm (x 3.52 mm) × 0.25-0.5 mm (x 0.36 mm), lanceate, basifixed, adpressed, bicolorous (base and central zone blackish brown and atrocastaneous margins), shining in mass, rigid, apices caudate, margins lacerate with glands. PETIOLES: 2.5-16 cm (x 8.72 cm), of length as long as the lamina, or near twice the length, or ½ of the lamina length, terete, light and dark castaneous, sometimes brown dark, striate, glabrescent or glandular-pilose. Hairs, ca. 5 cells long, glandular, simple, few inconspicuous spreading scales to base of axes. LAMINAE: 4.5-23.5(-26.5) cm (x 13.21 cm) long, herbaceous to chartaceous, narrowly ovate, linear a lanceate, lanceolate, glutinous (without dark yellow secretion), bi-tripinnate. Rachises terete and sulcate adaxial towards apex, hairs similar to the petiole, ca 1 mm long. Pinnae, petiolulate, lanceate, triangular. Segment petiolulate or adnate, margins lobulate, revolute, with visible apices veinlets. Abaxial surface glandular-pilose with hair apices cup-shaped and adaxially scattered glandular hairs. Pseudoindusium distinct, membranaceous, scarious, lunulate or lunate, margins erose with scattered hairs. Sori discrete, sporangia with 64 spores (Fig. 11).

Observations. Cheilanthes pilosa is one of two glandular hairy species in the Peruvian pteridoflora. Here, C. pilosa is provisionally recognized as a single species. It is a morphological variable species along its range, mainly in frond size and indument density (Ponce 2016, Kessler et al. 2017). We also found that characters of frond length, persistence of scales on axes and petiole morphology require a more detailed evaluation.
Two specimens from southeastern Peru (León 2233 and 2340) are included in the broad concept of C. pilosa, and show the need of further studies to clarify their identities. These specimens have the following characteristics: large size specimens, long-creeping rhizomes, rhizome scales, lanceate-lineal, caudate, dark castaneous, sclerotic, margin entire sometimes lacerate with glands on each laciniae; petiole terete, dark brown to dark reddish, 1/2 of lamina length, apparently almost glabrous but has spreading hairs of 1-2 cells (0.1 mm, 1 mm, 1.5 mm long) with apical gland, and with inconspicuous scattered scales that spread towards the rachis; rachis terete, only sulcate adaxially towards the most distal parts, hairs of 1-3 cells (0.1 mm, 0.5 mm, 1 mm, 1 mm long) with apical gland, and denser with respect to the petiole; pinnae with hairs of 0.1 mm, 0.5 mm, 1mm, denser, and adaxially their segments may be glabrescent; visible and broad apices veinlets; pseudoindusium distinct, membranaceous.
These specimens have some morphological similarities with the Bolivian endemic C. glutinosa Kessler & A.R. Smith by presence of septate short hairs on the axes, and dark castaneous, sclerotic rhizome scales; they are also morphological similar to the Ecuadorian endemic C. ecuadorensis Windham & K. Sosa by having short and long hairs on the lamina and scales on the rhizome.
Cheilanthes ecuadorensis belongs to a different clade than C. pilosa, and thus they are not phylogenetically close (Sosa et al. 2021). Cheilanthes glutinosa has also affinities with C. pilosa, and it is suspected to occur in Peru (Kessler et al. 2017).
Another morphologically similar species, C. andina, belongs in a different clade than C. pilosa, and thus they are also not phylogenetically close (Ponce & Scataglini 2022).
In addition, phylogenetic studies have shown strong support for the close affinities of C. pilosa to C. pruinata Kaulf. (Ponce & Scataglini 2018, 2022, and Sosa et al. 2021), both species are members of the South American clade.
Distribution and habitat. Cheilanthes pilosa ranges from Peru and Bolivia to Argentina. In Peru, it is known in 11 departments of the central and southern Peruvian Andes (Ancash, Apurimac, Arequipa, Ayacucho, Cusco, Huancavelica, Junin, Lima, Moquegua, Puno, and Tacna), between 2600-5010 m a.s.l. It grows in rock interstices and rocky sites associated to relict Polylepis forests, high Andean grasslands, and cryoturbated soil ecosystems.
Examined specimens. without locality, Rauh W. 1237 (USM). Ancash: Cerrate E. 1548, 2337, 3341, 3907, 6505, 7422, 8308, (USM); Sagástegui A. 12343 (HUT); Smith D. 11554, 11713, 11459a (USM), and 12141 (HUT); Weberbauer A. 2883 (USM). Apurimac: Rivera S. 96 (USM). Arequipa: Werff H. 20758 (HUSA). Ayacucho: Cano A. 11924, 12004, 19809 (USM); Lehnert M. 335 (USM); Morales M. 4085 (USM); Rivera S. 97(USM); Tovar O. 6701 (USM). Cusco: Cano A. 20583 (USM); León B 2233, 2340 (USM); Pennell 13994 (F, GH, US); Vargas 6541 (CUZ); Bües 1825 (US). Huancavelica: Rauh W. 359 (USM); Tovar O. 3117, 3126 (USM); Weigend M. 97/606 (USM). Junin: Beltrán H. 7135 (USM); Cerrate E. 5408 (USM); Killip E. P.23365 (US); Rivera S. 001 (USM); Saunders 647 (GH); Tryon 5467 (F). Lima: Beltrán H. 8248 (USM); Cerrate E. 4412 (USM); Gonzáles P. 2661 (SUM); León B. 905 (USM); Rivera S. 31(USM). Moquegua: Blanchard M. 86 (HUSA, USM); Montesinos D. 2025, 2176 (USM). Puno: Acleto C. 11185 (USM); Aguilar P. s.n. and s.n. (USM); Iltis H. 1317, 1320 (USM); Helena M. s.n. (USM); Mexia Y. 7783 (F, GH, MO, UC, US); Montesinos D. 2672 (HUSA, USM); Pennell F. 13454 (F, GH, US); Vilca S. 48 (HUSA); Weberbauer A. 717 (B); 909 (USM); Tacna: Cano A. 8235 (USM).
Cheilanthes poeppigiana Mett. ex Kuhn
Linnaea 36: 84. 1869. Syntypes: Peru, E.F. Poeppig s.n., and Bolivia, H. Cuming s.n. (B-B200038108a is determined by Mettenius and is doubtless part of the original materials).
FRONDS: 10.7-33 cm (x 21.04 cm) long. RHIZOMES: short, erect or decumbent, scaly. Scales 3-6.5 mm (x 3.88 mm) × 0.3-0.7 mm (x 0.43 mm), basifixed, bicolorous (base and central zone blackish brown and light castaneous margins), lanceate, apices caudate and sinuate with extended apical gland, margins inconspicuously uncinate, toothed or entire. PETIOLES: 5-17 cm (x 11.64 cm), of length as long as the lamina, or near twice the length, laevigate or striate, terete, sulcate adaxially without wings, atrocastaneous to atropurpureous (extending to the rachises), glabrous, inconspicuous spreading scales. LAMINAE: 5.7-17 cm (x 9.4 cm) long, ovate to triangular, unipinnate-pinnatisect to bipinnate-pinnatifid, glabrous. Rachises adaxially sulcate and wings, spreading scales. Pinnae triangular or lanceate-triangular. Ultimate segment adnate, lanceate, revolute margins, visible and broad apices veinlets. Pseudoindusium, membranaceous, short ending at the base of the segment, linear, margins erose or laciniate. Sori discrete, sporangia with 64 spores (Fig. 12).

Observations. Cheilanthes poeppigiana is an easily distinguishable species from others in the genus in Peru, by its glabrous laminae with modified margins only along segments. Because of its elongated pseudoindusium, Tryon and Tryon (1982) suggested affinities in the “C. marginata group”, however, this latter species was later recognized in the segregated genus Gaga, as G. marginata (Kunth) Fay W. Li & Windham (Li et al. 2012), having a modified margin that runs from segment to rachis.
Distribution and habitat. Cheilanthes poeppigiana is known from Ecuador to Argentina. In Peru occurs in Cajamarca, Cusco, Huanuco, Junin, La Libertad, Lambayeque, Piura, Puno and Tumbes, between 150-3500 m a.s.l. It grows in protected and humid rocky slopes, can be associated to Selaginella subg. Rupestrae, and shrublands.
Examined specimens. Cajamarca: Diaz A. 5070 (USM); López A. 3681, 3725, 5217 (HUT); Sagástegui A. 10596 (HUT). Cusco: Ferreyra R. 2684 (USM); León B. 455 (USM); Rauh W. 778, 876 (USM); Tryon R. 5396 (USM). Huanuco: Macbride J. 3672 (F). Junin: Cerrate E. 2808 (USM), Tryon R. 5436 (USM). La Libertad: Sagástegui A. 194 (HUT). Lambayeque: López A. 4042 (HUT). Piura: López A. 7703 (HUT). Puno: Weberbauer A. 566 (B). Tumbes: Coronado P. 226 (GH, UC); Schunke J. 3031 (F, GH, US).
Cheilanthes pruinata Kaulf.
Enum. Filic. 210. 1824. Type: Peru, non-designated locality, A. d. Chamisso s. n. (not located). “Note: Until now we have not been successful in the search of the specimen A. d. Chamisso s. n. cited by Kaulfuss in the original description, in the oldest herbaria of Europe, such as B, BM, BR, C, HAL, HBG, K, KIEL, L, LE (main collection), LZ (destroyed), M, P, S, W (Ponce & Scataglini 2022)”.
Cheilanthes mathewsii Kunze, Farrnkräuter 1: 50, t. 25. 1840. Type: Peru, A. Mathews 605 (holotype B-B200C38153).
Cheilanthes fasciculata Goldm., Nov. Act. Acad. Caes. Leopold.-Carol. Nat. Cur. 19 (suppl. 1): 456. 1843. Type: Peru, F.J.F. Meyen (holotype B-B200038142).
FRONDS: 15.8-37(-41.2) cm (x 26.91 cm) long. RHIZOMES: multicipital, short or long-creeping, nodose. Scaly; scales 3-3.6 mm (x 3.27 mm) × 0.3-0.6 mm (x 0.42 mm), lanceate, basifixed, adpressed, bicolorous (base and central zone blackish brown and light castaneous margins), shining in mass, rigid, margins entire, apices caudate with extended apical gland. PETIOLES: 5.8-13.6(-15.5) cm (x 9.29 cm) long, ½, ⅔ of the lamina length, castaneous and dark brown, terete, striate, glabrescent, glandular-pilose. Hairs simple with an apical modified glandular cell, scales spreading toward the rachis. LAMINAE: 10-28.5 cm (x 17.62 cm) long, subcoriaceous, linear or narrowly lanceolate, bipinnate-pinnatifid, glandular-pilose, glutinous (with dark yellow secretion), hairs as those on petiole. Rachises with hairs of different sizes, terete. Pinnae ovate to triangular, petiolulate. Segment adnate or shortly petiolulate, margins lobulate, revolute, apices veinlets mostly not visible. Abaxial surface glandular-pilose, glutinous, hairs simple with 1-2 cells up to 8 and adaxially with spreading hairs or glabrescent. Pseudoindusium distinct, membranaceous, scarious, lunate or lunulate, margins erose. Sori discrete, sporangia with 64 spores (Fig. 13).

Observations. Cheilanthes pruinata is one of the most common species in Peru, and is easily distinguished by its glandular hairs and glutinous fronds. It was considered within the “C. micropteris group”, based on the presence of glandular hairs; however, molecular phylogenetic studies (Ponce & Scataglini 2018, 2022, and Sosa et al. 2021) strongly support its close affinity to C. pilosa. Cheilanthes pruinata is easily recognized from C. pilosa by its usually lineate and glutinous laminae with dark yellow secretion vs. ovate-triangular and glutinous laminae without dark yellow secretion. Both species belong to the South American clade (Ponce & Scataglini 2022).
Distribution and habitat. Cheilanthes pruinata is found from Peru to Argentina. In Peru grows in all departments with Andean territory, except Pasco and Piura. It grows in crevices among rocks in slopes and ravines associated to relict Polylepis forests, seasonally dry tropical forests, Andean grasslands, Andean shrublands and Puya-stands ecosystems, between 1400-4580 m a.s.l.
Examined specimens. without locality, Rauh W. 578 (USM). Ancash: Cano A. 12382, 13364, 13432 (USM); Cerrate E. 162, 306, 500, 637, 654, 691, 791, 2174, 2523, 3103, 3287, 3382, 3874, 6498, 8013, 8292, 572a, s.n. (USM); Ferreyra R. 3287, 7490, 14539, 16876 (USM); Gonzáles P. 3257 (USM); La Torre M. 160, 161 (USM); Pérez E. 92 (USM); Smith D. 9058, 9344, 9398, 9862, 10460, 10593, 10772, 11285, 11550, 11941, 12408 (USM); Tovar O. 9601 (USM); Weberbauer A. 2670, 2864 (USM). Apurimac: Espinoza B. 499 (USM); Hocking P. 13a (USM). Arequipa: Aedo C. 10992, 11087 (USM); Arenas G. 104 (USM); Arenas L. 53 (HUSA); Cáceres F. 271, 1594, 2837, 5511, 8533 (HUSA); 5335 (USM, HUSA); Cano A. 22318 (USM); Castillo G. 8 (HUSA); Clostre G. 72 (USM); Cueva M. 35 (HUSA); La Torre M. 3338 (USM); León B. 5739b (USM); Mariño L. 4 (HUSA); Montesinos D. 3407 (HUSA), 3464 (USM, HSP); Museo de Historia Natural 2 (USM); Nuñez E.M. 65 (USM); Pauca A. 41 (HSP); Quipuscoa V. 2521 (HUSA, HSP), 2547(HUSA), 5920 (HSP, USM), 5945 (HSP, USM), 6885 (HSP, USM), 7374 (HSP); Ramos D. 1731, 1950 (HSP); Solomon J. 2858 (USM); Treviño I. 741 (HSP); Weibel M. 6 (USM); Weigend M 2000/518 (USM, HUSA), 9911/ MW 19-99 (USM, HUSA). Ayacucho: Cano A. 11863, 12006, 12030, 12069 (USM); Ferreyra R. 7214 (USM); La Torre M. 3277 (USM); Lehnert M. 331(USM); Roque J. 3136, 3148, 3152, 5442 (USM); Tejada C. 294 (HSP); Tovar O. 5667 (USM); Treviño I. 1561 (HSP). Cajamarca: Douglas W. 22037 (USM); Ferreyra R. 3224, 3322 (USM); Gutte P. 8748 (USM); Henning T. 20 (USM); Hutchison P. 5131 (USM); Pennell F. 15128, 15134 (USM). Cusco: Bonino E. 1005 (USM); Cano A. 20584 (USM); Davis E.1514, 1654, 1661(USM); Gutte P. 9498 (USM); Iltis H. 935a (USM); León B. 444 (USM); Nuñez P. 6694, 7417, 7977, 5362 (USM); Tryon R. 5362 (USM); Weberling F. 6529 (USM). Huancavelica: Cano A. 21872 (USM); Tovar O. 920, 926, 2029 (USM). Huanuco: Ferreyra R. 1943 (USM); Moya T. 2135 (USM). Junin: Cerrate E. 2901, 5409 (USM); Iltis H 130 (USM); Soukup J. 3147 (USM); Sullivan G. 1033 (USM); Tovar O. 5074 (USM); Yarupaitán G. 1133 (USM). La Libertad: Morales M. 3790 (USM); Pulido V. s.n. (USM); Roque J. 5002 (USM); Smith D. 2219, 3323 (USM); Weigend M. 97330, 97/248 (USM); Young K. 2917 (USM). Lima: Acleto C. 47 (USM); Baldeón S. 8120, 8122 (USM); Beltrán H. 8254 (USM); Cerrate E. 1005, 1740, 4224, 4260, 4485, 4713 (USM); Exp. Científica Univ. Varsovia Polonia 59 (USM); Ferreyra R. 5311 (USM); Gonzáles P. 489, 671, 3701, 3754 (USM), 672 (USM, HUSA); Rivera S. 28, 30 (USM). Moquegua: Blanchard M. s.n. (USM, HUSA); Cáceres F. 708 (USM); Montesinos D. 514 (USM). Puno: Aedo C. 11111 (USM); Aguilar P. 111 s.n. (USM); Arteta M. 324 (HUSA); Del Carpio C. 725 (USM); Iltis H. 1318 (USM); Mexia Y. 7780 (USM); Montesinos D. 2669 (HUSA, USM), 4162 (HSP); Quipuscoa V. 2498 (HUSA); Rauh W. 656 (USM); Solomon J. 2894 (USM); Tessene M. 1999 (USM); Tovar O. 5341 (USM); Vilca S. 41(HUSA); Zúñiga E. s.n. (USM). Tacna: Cano A. 8172, 8238, 8308 (USM); La Torre M. 1920, 2006. 2268, 2367, 2371 (USM).
Cheilanthes rufopunctata Rosenst.
Meded. Rijks-Herb. Leiden 19: 9. 1913. Type: Bolivia, (La Paz), Araca, October 1911, T. Herzog 2366 (holotype L-0063079; isotype UC-UC478369, US-US1187797).
FRONDS: 17.5-37.8 cm (x 22.13 cm) long. RHIZOMES: short or long-creeping, nodose, scaly. Scales 1.4-3 mm (x 2.03 mm) × 0.3-0.5 mm (x 0.39 mm), basifixed, adpressed, rigid, dark castaneous, partially sclerotic, lanceate, apices slightly caudate, margins entire. PETIOLES: 7.3-16.5 cm (x 11.2 cm) long, of length as long as the lamina, or near twice the length, brown or atrocastaneous, terete, laevigate, striate, adaxially sulcate, glabrescent, inconspicuous spreading scales. LAMINAE: 7.5-11.3 cm (x 9.42 cm) long, lanceate, ovate to triangular, bipinnate-pinnatifid to tripinnate-pinnatifid at the base, glabrous (except by reddish glands on abaxial side). Rachises adaxially sulcate and wings. Pinnae petiolulate (and wings). Adnate segment, lanceate, elliptic, lobed, segment margins revolute. Veinlets with broad apices, adaxially visible. Pseudoindusium broad, membranaceous, restricted to the segment, margins erose, laciniate. Sori discrete, sporangia with 64 spores (Fig. 14).

Observations. Cheilanthes rufopunctata is easily recognized by its mostly tripinnate-pinnatifid laminae with wings rachises, and abaxial surface of lamina with reddish glands. Its morphology resembles C. marginata Kunth, a species later included in the segregated genus Gaga, as G. marginata (Li et al. 2012); however, the latter has short creeping to erect rhizome and a pseudoindusium that extends to axes of the segment.
Affinities of C. rufopunctata have been proposed within the South American clade (Ponce & Scataglini 2022, Ponce et al. 2023). Ponce et al. (2023) proposed C. volcanensis de la Sota sister to C. rufopunctata, based on molecular studies. Morphologically, both species are similar, C. volcanensis can be differentiated by the few glandular hairs on the abaxial surface (vs. conspicuous reddish hairs in C. rufopunctata).
Distribution and habitat. Cheilanthes rufopunctata is known from Peru and Bolivia. In Peru, it occurs in Apurimac, Cusco, Huancavelica, Junin, Lima, and Puno, between 2400-3450 m a.s.l. It occupies rocky slopes.
Examined specimens. Apurimac: Hocking P. 7 (USM). Cusco: Soukup 29 (F, US); Vargas C. 4004 (CUZ, UC, US). Huancavelica: Tovar O. 316 (USM). Junin: Kunkel G. 4821/2 (GH). Lima: Acleto C. 45 (USM); Ferreyra R. s.n. (USM); Macbride J. 285 (F, GH, US); Muller G.12474 (GH). Puno: Soukup J. 29 (UC); Vargas 29 (MO).
Cheilanthes scariosa (Sw.) C. Presl
Reliq. Haenk. 1(1): 65. 1825.
Acrostichum lanuginosum Willd., nom. illeg., Schr. Acad. Erf.: 31, t.3, f.4, 1802, non Desf. 1800. Type: Peru, Malaspina Expedition (holotype B-W 19554010).
Acrostichum scariosum Sw., Syn. Fil. (Swartz) 16. 1806, nom. nov. for A. lanuginosum Willd.
Cheilanthes ornatissima Maxon, Smithsonian Misc. Coll. 65(8): 3. 1915. Type: Peru, Lima, mountains back of Lima, March 1892, W.E. Safford 996 (holotype US; isotypes F, GH-GH0002748, MO, UC, US).
Cheilanthes tripinnata Copel., Univ. Calif. Publ. Bot. 19: 301, t.58. 1941. Type: Peru, Dep. Apurimac, Chincheros to Andahuaylas, J. West 3724 (holotype UC).
FRONDS: 12-32.2 cm (x 21.24 cm) long. RHIZOMES: multicipital, short-creeping or erect, compact, scaly. Scales 5.8-19 mm (x 9.40 mm) × 0.05-0.8 mm (x 0.37 mm), basifixed, castaneous, atrocastaneous, ferrugineous, linear-acicular, with slightly sinuate apices, margins toothed, inconspicuously uncinate. PETIOLES: 3.4-9 cm (x 5.98 cm) long, ½, ⅓, ¼ to ⅕ or sometimes of length as long as the lamina or ⅔ of the lamina length, terete, dark brown, striate or scabrous. Scales sub-basifixed or basifixed, variable shape, color (atrocastaneous or castaneous center and whitish margins), margins irregularly laciniate, apices cuneate caudate. Squamules ca. 5 cells in the central area, light castaneous, arachnoid-like, filamentous. LAMINAE: 9-26 cm (x 15.26 cm) long, linear or narrowly lanceolate, bi-or-tripinnate, scaly. Scales 2.3-5.5 mm (x 3.68 mm) × 0.6-2.0 mm (x 1.22 mm), imbricate, basifixed, or sub-basifixed along axes. Rachises striate, scabrous, indument of all lamina axes as their petioles. Pinnae petiolulate. Segment petiolulate, orbiculate or flabellate, margins revolute. Abaxial surface scales with reduced cell lumen, castaneous or greyish, lanceate, triangular, ovate or elliptic, apices caudate, acute, margins laciniate accompanied with long filaments extending to the adaxial surface, arachnoid-like, ca. 5 cells in the central area (called squamules). Pseudoindusium not differentiated or sometimes hyaline. Sori discrete, sporangia with 32 spores (Fig. 15).

Observations. Cheilanthes scariosa can be recognized from other scaly species by its mostly orbiculate ultimate segment, laminar scales with margins laciniate accompanied with long filaments extending to the adaxial surface, arachnoid-like, ca. 5 cells in the central area (see in taxonomic key), and with reduced cell lumen. Together with other densely scaly Peruvian species, that include C. arequipensis, C. incarum, C. lonchophylla and C. peruviana, is recognized as members of the South American-Australasian clade (Ponce & Scataglini 2022).
Distribution and habitat. Cheilanthes scariosa is known from Peru and Bolivia. In Peru, it occurs in Ancash, Apurimac, Cusco, Huancavelica, Junin, La Libertad, Lima, Pasco and Puno, between 2450-4400 m a.s.l. It occupies calcareous rocky sites with Andean grasslands and high Andean shrublands and relict Polylepis forests ecosystems.
Examined specimens. Ancash: Cano A. 13222 (USM); Cerrate E. 2200, 12145 (USM); López A. 2388 (USM, HUT); Zósimo s.n. (HUT). Apurimac: Rivera S. 51, 52, 53, 54 (USM); Vargas 8792 (CUZ, UC). Cusco: Vargas C. 23239 (GH). Huancavelica: Tovar O. 113, 798, 2087 (USM). Junin: Cerrate E. 2909 (USM, HUT); Gentry A. 39760 (USM); Iltis H. 60, 117 (USM); Hodge W. s.n. (USM); Petersson U. s.n. (USM); Tryon R. 5457 (USM); Tvlles 804 (USM); Weberbauer A. 2531 (USM). La Libertad: Gonzáles P. 6012 (USM); López A. 3395 (HUT); Sagástegui A. 17344 (HUT). Lima: Goodspeed T. 11347 (GH, UC); Macbride J. 424 (F, US); Rauh W. 153, 1833 (USM); Rivera S. 23 (USM). Pasco: Baldeón S. 1444, 3853, 7517, 7532 (USM); Gentry A. 37495 (USM). Puno: Weberbauer A. 425 (B).
Conservation status of Peruvian endemic Cheilanthes
Levels of endemism among Peruvian ferns is lower compared to other vascular plants, however, there are important patterns, for example richness, as in the case of Cheilanthes, with 25% of its species (i.e., C. cantangensis, C. fractifera, C. lonchophylla, and C. peruviana) being restricted to Peru.
None of these species are listed by the IUCN, even though they were evaluated earlier by León (2006). Here we reassessed their conservation status following the IUCN criteria (version 3.1, 2012). The justification for the reevaluation is found in technical sheets developed per species (Appendix 1). Two species are considered Endangered (EN), and two of Least Concern (LC).
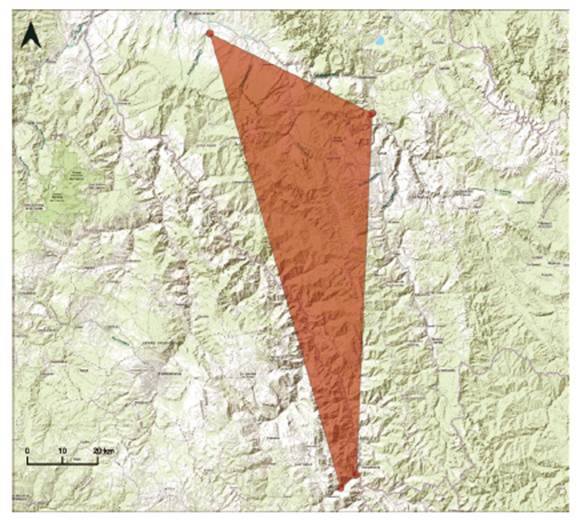
-Cheilanthes cantangensis is here proposed as Endangered (EN) B1ab(iii). It was previously categorized by León (2006) as Critically Endangered (CR), B1ab (iii) because at that point in time, it was known only from a single collection made in the 1960’s. This terrestrial species is currently known from two localities in the Marañon and Utcubamba basins (Fig. 16). It occupies seasonally dry forests and scrub vegetation, in the departments of Cajamarca and Amazonas, and it might occur in other sites within its range.
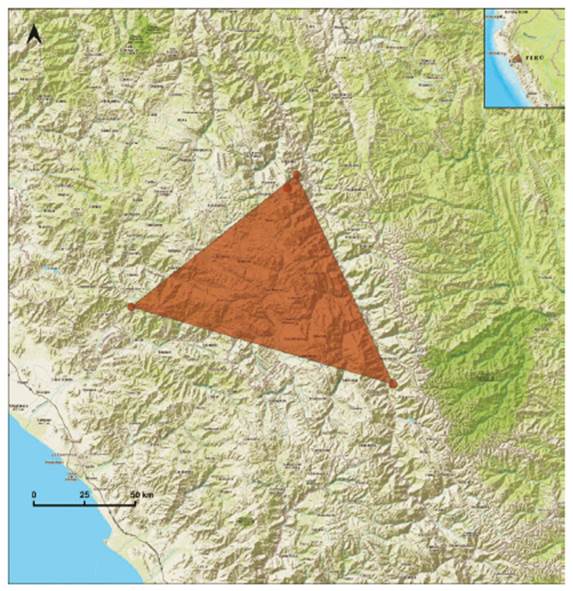
-Cheilanthes fractifera is proposed as of Least Concern (LC). It was previously categorized by León (2006) as Vulnerable (VU) B2ab(iii). This species occurs in dispersed and highly fragmented populations on the Pacific basin, from La Libertad to the north, Lima and Ayacucho in the center, to Arequipa in the south (Fig. 17). Possibly, the species occupies other areas within its range on the Pacific slopes. The type specimen was collected at 2200 m asl, 45 km from Nasca, on the road to Puquio, Ayacucho, in 1958, but unfortunately no additional records exist for this site.
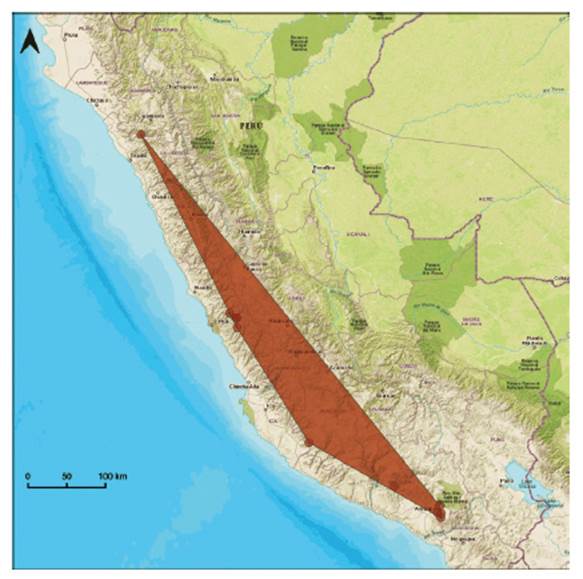
-Cheilanthes lonchophylla maintains the category of Endangered (EN) B1ab(iii) proposed by León (2006). Most records were made between the years 1960-1980, while a single collection was made in 2020. It is known from both sides of the Andes, has disjunct localities on the Pacific slopes and seasonally dry valleys of the Marañon basin (Fig. 18). Specific threats are unknown.
-Cheilanthes peruviana was categorized as of Least Concern (LC), a category also proposed by León (2006). It is known from several populations distributed in the western slopes of the Andes, and some interandean valleys in northern Peru, having a wide altitudinal range. It grows in “lomas” ecosystems, such as Atiquipa, Capac, Cachendo and Tacahuay (Fig. 19), also in western Andean forests, and dry scrubs. Despite occupying different habitats, the status of each population has not been evaluated.
All four endemic species require further work to clarify conditions and population trends (i.e., size, biology, location, and threats), as it is also of their habitats. Only one species is represented within a protected area. The uses for these four species remain unknown.
For species not restricted to Peru, three of them (C. incarun, C. scariosa and C. pruinata) require attention on their conservation status, given that they are being extracted from natural populations and traded as medicinal in most cases. Some of these species are considered a substitute for Erythroxylum coca "coca" in Peru (Tryon 1959; Navarrete et al. 2006). Cheilanthes incarum and C. scariosa are morphologically similar and could be confused by their users. Both are known from Peru and Bolivia and possible impacts on the state of their populations are unknown. Cheilanthes pruinata “cuti cuti” is used in folk-medicine (Navarrete et al. 2006) and is extracted from wild populations and traded locally.
Recently, a photograph of an unconfirmed species of Cheilanthes, from Peru, is advertised as the “Amazon diabetic tea”, a product sold online (https://amazonrivertea.com/the-tea/), however, its attributes are those linked to Argyrochosma nivea. This case demonstrates the importance of accurate surveys, sound identifications, and evaluations on potential impacts on wild populations.
Conclusions
The Peruvian species of Cheilanthes are important components of Andean ecosystems (Tryon 1964, Tryon and Stolze 1989, León and Valencia 1998, León et al. 1996, 2022), some extending to the flanks of the Amazon basin.
The fifteen species of Cheilanthes recognized here represent 31% of all species in the subfamily Cheilanthoideae in Peru’s flora. They are morphologically distinguishable; however, species limits are still a work in progress, especially when using only morphological characters in the frond, as lamina-petiole ratio, frond division, and indument density, as it has been discussed in other Andean fern floras (Ponce 2016, Kessler et al. 2017).
The type of indument shared by Peruvian Cheilanthes allow the distinction of three main groups that are not completely congruent with phylogenetic clades proposed by Ponce and Scataglini (2022). The first group includes simple or branched hairs or glabrous species, i.e., C. mollis, C. fractifera, C. fraseri, C. rufopunctata, C. poeppigiana, C. pruinata, C. pilosa and C. andina, all these species representing the South American clade; the second group have acaroid-like hairs or a combination of simple hairs plus scales in the frond, i.e., C. obducta and C. cantangensis; in the third group, there is a dominance of scales in one or both surfaces of the frond and linear-acicular rhizome scales, i.e., C. scariosa, C. incarum, C. lonchophylla, C. arequipensis and C. peruviana. These last two groups are members of the Australasian-South American clade in the new concept of Cheilanthes.
The genus Cheilanthes still requires additional observations for several morphological characters, such reproductive expression (examined for two species of the Peruvian flora by Martínez et al. 2017), spore surface macro-ornamentation (not examined here) and venation patterns. The genus is known to present variable positions of the meristematic tissue in the gametophyte, that may inform phylogenetic affinities (as shown by George et al. 2019), a life stage rarely explored for Peruvian taxa.
This study documents that sampling of Cheilanthes species in Peru is still in deficit, as is poorly represented in Peruvian herbaria. For example, C. pruinata, a common species along the Andes has still not been recorded from Piura and Pasco, despite its presence in neighboring departments. For rare species, this shortfall of collections is accompanied by highly spaced collections in years, and highly dispersed known populations in climatic seasonal environments. A detailed survey of the pteridoflora, and particularly Cheilanthes, will be important for future assessment of threats and reevaluation of their conservation status.
For traded species, exploited from wild populations (Navarrete et al. 2006), detailed field evaluation of population size and locations are needed, as it was demonstrated for Dicksonia in Santa Catarina, Brazil (Gasper et al. 2011).

